細菌脂多糖對甜味受體T1R2可變剪接與功能的調控作用
2021-08-14 07:03:04朱健慧鄭欣彭顯徐欣RobertMargolskee周學東
華西口腔醫學雜志 2021年4期
關鍵詞:小鼠
朱健慧 鄭欣 彭顯 徐欣 Robert Margolskee 周學東
1.口腔疾病研究國家重點實驗室 國家口腔疾病臨床研究中心四川大學華西口腔醫院牙體牙髓病科,成都610041;2.Monell Chemical Senses Center,美國費城PA 19104
哺乳動物可感知甜、酸、咸、苦、鮮五種基本味覺,味覺感知對于個體攝取營養物質、鑒別有毒有害物質有重要意義[1]。味覺異常是以味覺改變為特征的疾病,處于味覺異常狀態的患者容易出現食欲減退、營養不良、情緒消極等,嚴重影響生活質量[2]。近日研究[3]報道新冠肺炎患者出現味覺感知障礙或味覺喪失,提示微生物對味覺感知的重要影響。味覺異常的病因尚不明確,但口腔衛生狀況不佳、牙周病等口腔局部微生物因素與味覺感知的變化具有相關性[4]。
味覺感受器中的味覺受體細胞被某些物質激活后,通過感覺神經將味覺信號傳遞至大腦相應區域,形成味覺感知。甜味受體屬于G蛋白偶聯受體(G protein-coupled receptors,GPCRs)C家族,為Ⅰ型味覺受體家族(taste receptor type 1,T1R)中的兩種亞型T1R2和T1R3組成的異源二聚體,可被糖(葡萄糖、果糖等),人工甜味劑(糖精、安賽蜜等)及甜味蛋白(莫奈林、神秘果素等)等甜味配體激活產生甜味信號[5]。研究[6]表明,小鼠攝取脂多糖(lipopolysaccharide,LPS)后,對蔗糖的甜味敏感度顯著降低,但其機制尚不明確。
可變剪接指來源于同一基因的前體mRNA經過不同方式的剪接,形成不同的成熟mRNA,進而翻譯為結構、功能不盡相同的蛋白質。可變剪接是真核生物中廣泛存在的轉錄后調控方式,具有重要的生物學意義[7]。LPS可通過調控Toll樣受體下游分子MyD88編碼基因的可變剪接影響其在炎癥反應中的功能[8]。T1R2前體mRNA包含6個外顯子和5個內含子,可能受到可變剪接調控,影響甜味感知功能。本研究旨在探究LPS對甜味受體可變剪接的影響,從轉錄后調控角度揭示口腔微生物感染引起味覺感知異常的可能機制。
1 材料和方法
1.1 主要試劑和材料
RNA提取試劑盒MiniBEST Universal RNA Extraction Kit、逆轉錄試劑盒PrimeScript?RT reagent Kit with gDNA Eraser、高保真DNA聚合酶Prime-STAR?Max DNA Polymerase、實時熒光定量聚合酶鏈反應(quantitative real-time polymerase chain reaction,qPCR)試劑TB Green?Premix Ex Taq?Ⅱ、感受態細胞E.coliDH5αCompetent Cells(TaKaRa公司,日本),限制性核酸內切酶BamHⅠ-HF/NotⅠ-HF、T4連接酶(New England Biolabs公司,美國),無內毒素質粒小提中量試劑盒(北京Tiangen公司),lipofectamine 2000、標簽抗體6x-His Tag Monoclonal Antibody(Thermo Scientific公司,美國),胎牛血清(fetal bovine serum,FBS)、DMEM培養基、添加鈣鎂的DPBS溶液(Gibco公司,美國),penicillin/streptomycin(P/S,Hyclone公司,美國),標簽抗體Monoclonal ANTI-FLAG?M2 antibody(Sigma公司,美國),抗鼠熒光二抗Goat Anti-Mouse IgG H&L(Alexa Fluor?488/594,Abcam公司,英國),牛血清蛋白(bovine serum albumin,BSA)(BioFroxx公司,德國)。引物合成及DNA測序由北京擎科生物科技公司完成。
1.2 逆轉錄聚合酶鏈反應(reverse transcriptionpolymerase chain reaction,RT-PCR)
取8周齡C57BL/6J小鼠6只(雄性3只,雌性3只),使用頸椎脫臼法處死小鼠,體視顯微鏡下分離舌背包含味蕾組織的輪廓乳頭、菌狀乳頭及葉狀乳頭,分別提取RNA,逆轉錄獲得cDNA,RT-PCR擴增完整T1R2轉錄本(正向序列:5’-AAGGCTGTTACTTGGCTGGC-3’,反向序列:5’-GGTGGGTGGGACTAGCTCTT-3’),行瓊脂糖凝膠電泳,對長度小于T1R2本構體的DNA片段行TA克隆,隨機挑取單菌落測序。為進一步驗證異構體表達,在異構體缺失序列上、下游設計引物(正向序列5’-CAGTGGAACTGGATCGTGGT-3’,反向序列5’-GCGTCACAGTCCTGGTTACA-3’),以前述cDNA為模板,PCR擴增異構體對應DNA片段,并通過瓊脂糖凝膠電泳進行驗證。
1.3 甜味受體異源性表達實驗
1.3.1 質粒載體構建 RT-PCR擴增T1R2/T1R3本構體轉錄本、T1R2剪接異構體(T1R2_Δe3p)轉錄本,克隆至真核表達載體pcDNA3.1(+),測序驗證序列無誤。甜味受體信號通路下游G蛋白表達質粒Gα16-gust44-pcDNA3.1-hygro由Monell Chemical Sensens Center(費城,美國)提供。
1.3.2 細胞培養和轉染 HEK293細胞在添加10%FBS和1%P/S的高糖DMEM培養基中培養。轉染前將細胞接種至黑色透明底96孔板,按照lipofectamine 2000說明書操作,將等量T1R2本構體表達質粒和(或)異構體(T1R2_Δe3p)表達質粒、T1R3本構體表達質粒及Gα16-gust44表達質粒轉染至HEK293細胞。
1.3.3 FlexStation 3檢測 甜味受體功能轉染48 h后,吸出培養基,加載終濃度為2.5μmol·L-1的鈣離子熒光染料Fluo-4,避光孵育1 h后,再次DPBS漂洗,每孔加入50μL DPBS后避光孵育30 min。以DPBS為溶劑配制甜味配體溶液,其終濃度為蔗 糖100 mmol·L-1、葡 萄 糖100 mmol·L-1、果 糖100 mmol·L-1、糖精1 mmol·L-1、安賽蜜10 mmol·L-1、三氯蔗糖10 mmol·L-1[9],FlexStation 3自動移液酶標儀(Molecular Devices公司,美國)動態監測熒光值變化。繪制熒光強度(F)-時間曲線,如果配體可激活甜味受體,該曲線會在30~60 s之間出現峰值,計作Fpeak;每孔選取最后20 s內10個讀數的平均值作為基線,計作F0;繪制標準化熒光強度(F/F0)-時間曲線,其縱坐標可間接反映胞內鈣離子濃度變化。計算(Fpeak-F0)/F0代表受體-配體效應強度,繪制柱狀圖比較甜味受體功能。
1.4 細胞免疫熒光檢測
在異構體T1R2_Δe3p 3’端添加6His標簽,在T1R2本構體3’端添加FLAG標簽,克隆至真核表達載體pcDNA3.1(+)。使用lipofectamine 2000將等量T1R2-FLAG、T1R2_Δe3p-6His、T1R3及Gα 16-gust44表達質粒轉染至HEK293細胞。轉染48 h后,4%多聚甲醛固定15 min,PBS漂洗,0.2%Triton X-100通透10 min,3%BSA封閉30 min,6His或FLAG一抗(1%BSA 1∶1 000稀釋)孵育2 h,漂洗,用帶有熒光基團的二抗孵育1 h,再次漂洗,在10μg·mL-1DAPI溶液中孵育5 min,漂洗后使用倒置熒光顯微鏡觀察并拍照。
1.5 LPS局部注射及qPCR檢測
將8周齡、雄性C57BL/6J小鼠隨機分為LPS處理組(n=5)及PBS對照組(n=5),行注射麻醉,使用LPS(5 mg·kg-1)或等體積PBS行舌背局部注射,6 h后使用頸椎脫臼法處死小鼠。分離小鼠舌背輪廓乳頭與菌狀乳頭,分別提取RNA,逆轉錄為cDNA,利用T1R2_Δe3p(正向序列5’-GATCGTGGTGCTGGAGGTTC-3’,反向序列5’-GCTCAGCTCTGGCGAGAATA-3’)、T1R2(正向序列5’-CCAAGTGCAATGAGTACAACATGA-3’,反向序列5’-AGCAGACATCCACCATCTCG-3’)、T1R3(正向序列5’-GCAACCAGGTGCCAGTCT-3’,反向序列5’-AGCAGGCTGTGCTTTTCTCT-3’)特異性引物進行qPCR擴增。相對表達量以βactin(正向序列5’-GGTCAGAAGGACTCCTATGTGG-3’,反向序列5’-TGTCGTCCCAGTTGGTAACA-3’)為內參,采用2-ΔΔCt法計算,并計算T1R2_Δe3p相對T1R2的表達比例。
1.6 統計學分析
采用SPSS 20.0軟件進行分析,采用獨立樣本t檢驗,以雙向P<0.05為差異有統計學意義。
2 結果
2.1 甜味受體T1R2存在可變剪接異構體T1R2_Δe3p
味蕾組織RNA行RT-PCR擴增T1R2轉錄本,產物行瓊脂糖凝膠電泳后,發現T1R2存在若干較短的剪接異構體(圖1A),測序結果顯示其中一個剪接異構體的3號外顯子部分丟失。特異性引物qPCR反應后通過瓊脂糖凝膠電泳驗證該異構體表達,將其命名為T1R2_Δe3p(圖1B)。對丟失序列進行分析,發現其5’端(GU)及3’端(AG)為內含子特征序列,在mRNA剪接過程中可能被識別為內含子而丟失(圖1C)。開放閱讀框分析顯示T1R2_Δe3p翻譯起始與終止位點未發生改變,其翻譯產物缺少位于胞外段配體結合區域的29個氨基酸殘基(圖1D)。
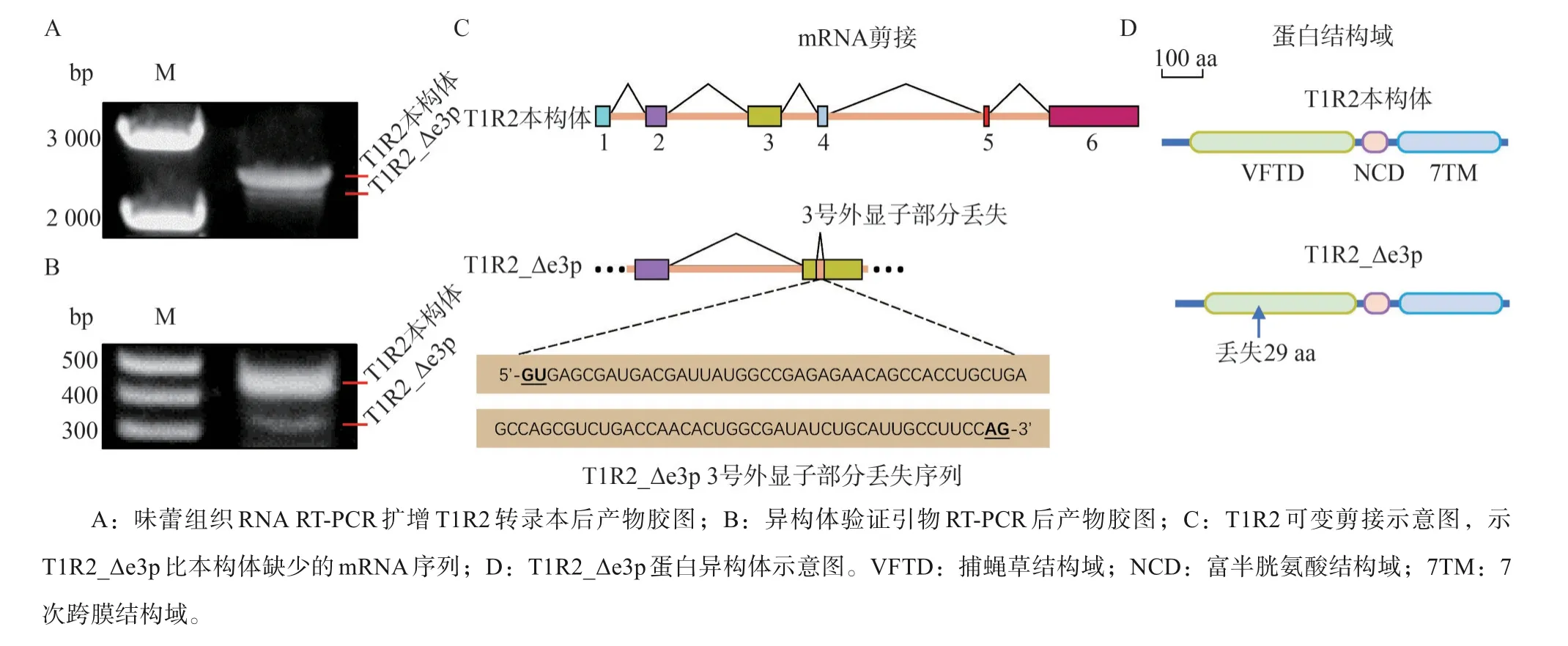
圖1 剪接異構體T1R2_Δe3p的發現Fig 1 Identification of splicing isoform T1R2_Δe3p
2.2 可變剪接異構體T1R2_Δe3p導致甜味受體功能下調
與T1R2/T1R3本構體相比,T1R2_Δe3p與T1-R3組成的甜味受體受到適當濃度的糖精、安賽蜜、三氯蔗糖、葡萄糖、果糖、蔗糖等甜味配體刺激后,細胞內鈣離子濃度變化顯著下調,且與陰性對照相比差異無統計學意義(圖2)。T1R2_Δe3p與T1R3組成的甜味受體無法與甜味刺激物結合產生味覺信號,表現為甜味感知功能的缺失。將T1R2_Δe3p與T1R2/T1R3共同轉染至HEK-293細胞,甜味配體刺激后胞內鈣離子濃度變化顯著下調,但高于陰性對照組(圖2)。qPCR與細胞免疫熒光圖像顯示過表達T1R2_Δe3p后,甜味受體本構體T1R2/T1R3的mRNA和蛋白表達水平無明顯變化(圖3)。
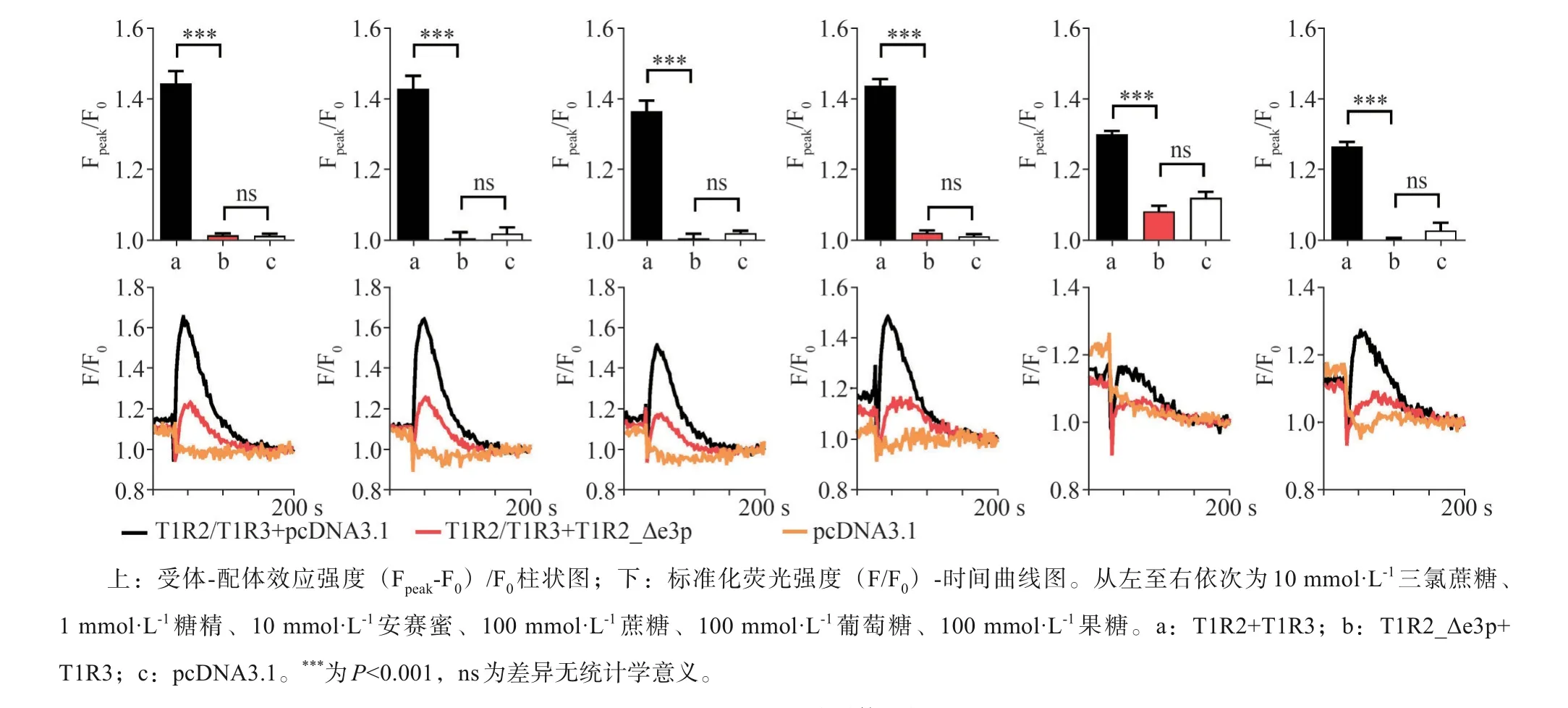
圖2 T1R2_Δe3p對甜味受體功能的影響Fig 2 The effect of T1R2_Δe3p on thefunction of sweet tastereceptor
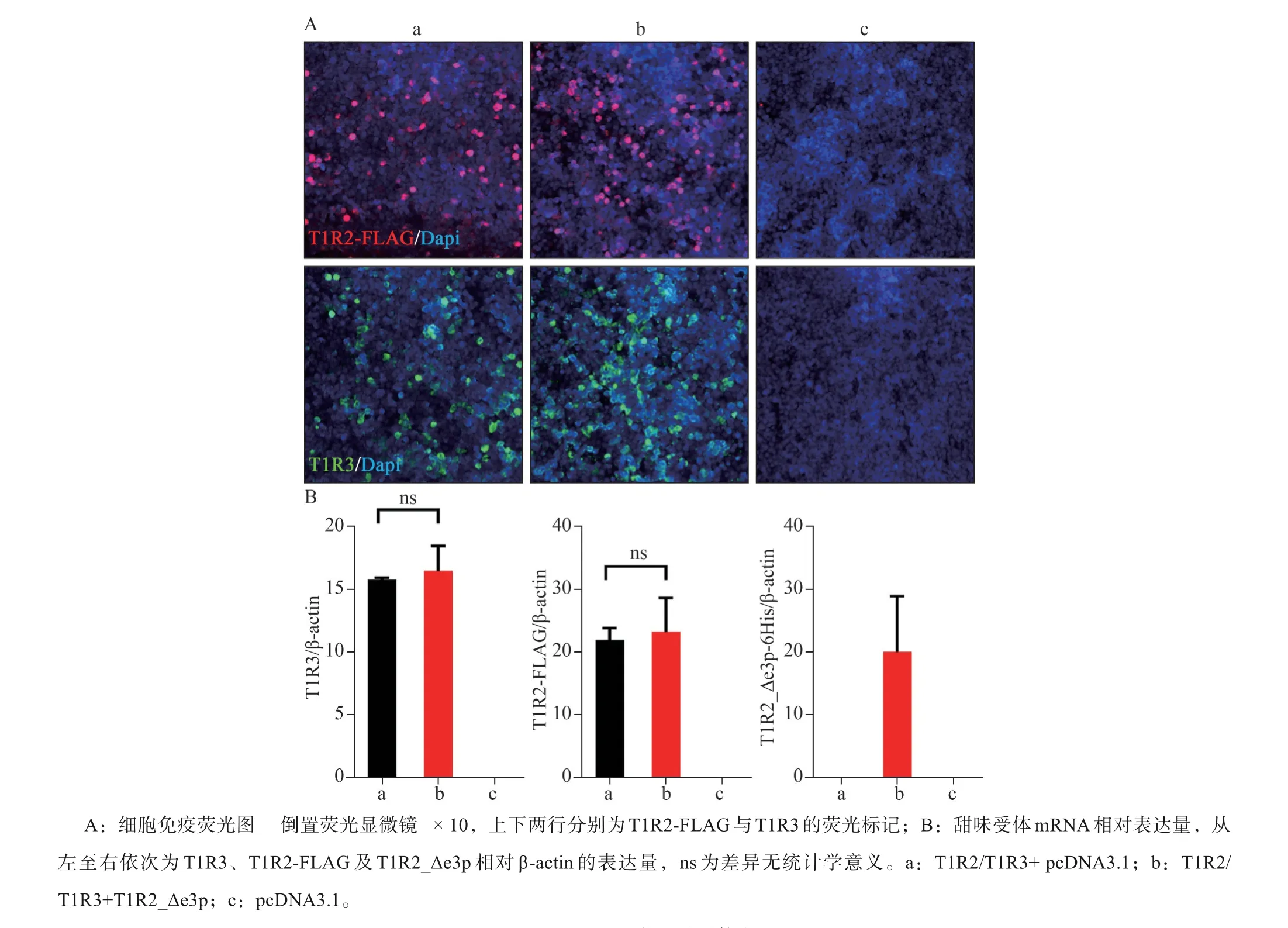
圖3 T1R2_Δe3p對本構甜味受體表達量的影響Fig 3 The effect of T1R2_Δe3p on the expression of canonical sweet taste receptor
2.3 局部注射LPS后T1R2_Δe3p表達比例上調
小鼠舌背局部注射LPS后,菌狀乳頭、輪廓乳頭組織qPCR結果顯示,甜味受體T1R2/T1R3表達量與對照組比較差異無統計學意義(圖4)。菌狀乳頭中T1R2_Δe3p表達量(P=0.045 4)及相對T1R2本構體表比例(P=0.017 8)均有顯著上升(圖4),輪廓乳頭T1R2_Δe3p相對T1R2本構體表比例顯著上升(P=0.037 1,圖4)。
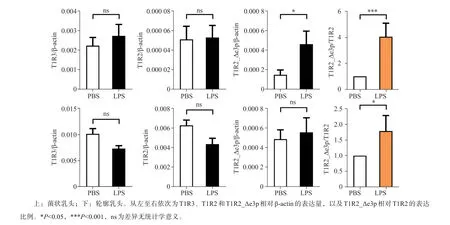
圖4 LPS對T1R2/T1R3本構體與T1R2_Δe3p表達量的影響Fig 4 Theexpression of canonical T1R2/T1R3 and T1R2_Δe3p affected by LPS
3 討論
人類的口腔作為消化道的起始,是人體與環境進行物質交換的重要場所,有超過600種微生物定植[10]。口腔微生物不僅是齲病、牙髓病、根尖周病、牙周病等口腔常見病、多發病的最主要致病因素,還與惡性腫瘤、糖尿病、心血管疾病等全身性疾病密切相關[11]。味覺感受器位于味覺乳頭內的味蕾中,主要分布于口腔內的舌背黏膜,微生物作為局部刺激因素對味覺傳導有一定影響。
口腔衛生狀況及感染性疾病的患病情況與味覺異常的發生有關[4]。LPS為革蘭陰性菌細胞壁的組成成分,可通過活細菌外膜囊泡或死細菌崩解的方式釋放,是重要的活性致病物質[12]。有研究[13]報道,腹腔注射LPS可升高小鼠味覺乳頭炎性因子的表達量,可能通過影響味覺前體細胞增殖參與味覺異常的發生。本研究發現了對甜味受體功能有顯著影響的T1R2_Δe3p,LPS局部刺激可使其表達比例升高,從轉錄后調控的角度解釋了微生物抗原成分引起味覺感知能力下降的原因。
甜味受體T1R2/T1R3屬于GPCR的C家族,除擁有GPCR超家族7次跨膜結構和偶聯G蛋白引起下游分子信號轉導的共同特點以外,其與配體結合的結構域位于500~600個氨基酸殘基組成的氨基端末端,具有類似于捕蠅草的雙小葉空間結構,被命名為捕蠅草結構域(Venus flytrap domain,VFTD)[14]。本研究發現的T1R2剪接異構體蛋白缺少的29個氨基酸殘基位于本構體的VFTD內。異源性表達實驗結果顯示T1R2_Δe3p與T1R3共轉染后無法結合甜味配體,推斷其原因為異構體蛋白無法形成可與甜味刺激物結合的VFTD結構。過表達T1R2_Δe3p后,T1R2/T1R3本構體在mRNA及蛋白質水平表達量無明顯變化,但甜味受體功能明顯下調。推測其機制為T1R2_Δe3p競爭性與T1R3結合形成無功能的二聚體,抑制T1R2/T1R3二聚體的形成,整體下調甜味受體與配體的結合。
mRNA可變剪接是轉錄后調控的關鍵步驟,異常剪接與腫瘤、免疫性疾病、神經退行性疾病、代謝性疾病等多種疾病相關[15]。可變剪接存在復雜的調控網絡,其中剪接因子可識別前體mRNA中剪接調控元件,發揮促進或抑制剪接的作用[16]。本研究發現剪接異構體T1R2_Δe3p對甜味受體功能有顯著抑制作用,其表達受到LPS局部刺激的調控。LPS影響甜味受體可變剪接的分子機制有待于進一步研究,推測LPS可能通過調控某些關鍵剪接因子對T1R2_Δe3p表達產生影響。進一步發現調控甜味受體可變剪接的關鍵剪接因子將為人工干預甜味受體功能提供理論基礎。此外,LPS局部刺激影響小鼠味覺乳頭內T1R2_Δe3p豐度變化是否對其甜味敏感度產生影響仍有待進一步證實。
綜上,本研究發現了小鼠甜味受體T1R2的可變剪接異構體T1R2_Δe3p,其對甜味受體T1R2/T1R3的功能有顯著抑制作用,LPS局部刺激可顯著上調T1R2_Δe3p異構體表達,從轉錄后調控角度揭示微生物感染影響宿主味覺感知的分子機制。
利益沖突聲明:作者聲明本文無利益沖突。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
