胡椒PnPAL基因家族全基因組鑒定及表達模式分析
2021-08-18 06:50:44蘇岳峰丁元昊郝朝運胡麗松鄭其向
福建農業學報 2021年6期
關鍵詞:分析
蘇岳峰,丁元昊,郝朝運,胡麗松,鄭其向,3,范 睿
(1.海南大學,海南 海口 570208;2.中國熱帶農業科學院香料飲料研究所,海南 萬寧 571533;3.云南農業大學熱帶作物學院,云南 普洱 665000)
0 引言
【研究意義】胡椒(Piper nigrumL.)被稱為“香料之王”,是胡椒科胡椒屬植物,主產我國海南、云南和廣東等地區。胡椒瘟病是由辣椒疫霉菌侵染胡椒的根系、葉片、花序、果穗、果實等部位而發病,具有氣候依賴性且傳染力極強的特點[1];病情嚴重時可導致全園毀滅,嚴重阻礙胡椒產業發展,缺乏抗瘟病種質是影響我國胡椒產業發展的主要問題[2]。因此,篩選胡椒關鍵基因進行功能驗證對植物材料的遺傳改良尤為重要。【前人研究進展】次生代謝是植物生長發育以及適應環境的重要環節。苯丙烷途徑是響應多種逆境的關鍵途徑,如:紫外線損傷、病原侵染、機械損傷、低溫脅迫等。苯丙氨酸解氨酶是苯丙烷途徑的關鍵酶和入口酶[3]。玉米的苯丙氨酸解氨酶(Phenylalanine ammonialyase,PAL)抵抗甘蔗花葉病毒,與木質素和水楊酸合成有關,陸地棉和馬鈴薯等中也有PAL 抵抗病菌侵染的報道[3-4]。此外苯丙氨酸解氨酶還是合成花青素和類黃酮等重要的脅迫響應次生代謝物質的必要酶[5]。目前,PAL的生物信息學分析已在水稻、棉花、桑樹等作物中開展,分析結果顯示PAL基因響應生物和非生物脅迫反應[6-8]。PAL在單雙子葉植物分化之前已完成分化,通常是一個多基因家族,含有5 個左右家族成員[9]。PAL家族在不同作物中保守域不同,主要包括:苯丙氨酸保守結構域、PLN02457 和PLN02457 super family、PAL-HAL、phe-am-lyase 和Lyase aromatic 保守結構域、裂解酶I 類超家族保守結構域等[10-11]。然而,各物種所含保守域不同可能行使不同的生理功能,當歸中PAL 催化類似的β 消除反應,與具有裂解酶I 類超家族保守結構域有關[12]。在柱花草中,PAL-HAL 是保守的脫氨位點[11]。目前,胡椒瘟病防控多以預防為主,防控成本極高。在分子生物學方面對胡椒瘟病的研究也僅限于栽培種與野生近緣種的比較轉錄組學研究和抗瘟病基因挖掘[13]。項目組前期的轉錄學結果顯示胡椒抗/感種質(熱引1 號、黃花胡椒)差異基因多富集于苯丙烷代謝途徑,且組織化學顯示苯丙氨酸解氨酶活性在抗性種質中較高[13]。本研究首次開展胡椒PnPAL基因研究。【本研究切入點】苯丙氨酸解氨酶是苯丙烷途徑的入口酶及限速酶,在植物與病原菌互作過程中具有重要意義。因此,研究胡椒PnPAL的家族特征、進化關系及表達模式,能夠為進一步確定胡椒PnPAL基因在抗瘟病中的功能奠定基礎。【擬解決的關鍵問題】采用在線分析工具及TB tools 工具,胡椒PnPAL的順式作用元件、motif、外顯子和內含子位置、保守結構域,親緣關系及接種病原菌下PnPAL基因的表達模式,有利于了解胡椒PnPAL的基因功能,便于胡椒新品種選育。
1 材料與方法
1.1 試驗材料
我國主栽胡椒品種熱引1 號(Piper nigrumL.)全基因組數據來自美國生物信息中心https://www.ncbi.nlm.nih.gov/bioproject/PRJNA529758。
1.2 試驗方法
1.2.1PnPAL基因家族成員鑒定 根據胡椒基因組注釋文件信息,鑒定到胡椒PnPAL基因家族成員和胡椒PnPAL的蛋白序列及染色體定位信息,再利用在線預測蛋白信息工具Expasy(https://www.expasy.org/)分析各個PnPAL的蛋白質分子質量、等電點和氨基酸個數,運用BUSCA 在線分析工具預測各PnPAL亞細胞定位(http://busca.biocomp.unibo.it/59a16590-35 06-4818-8017-7032b057b649/showresult/)。
1.2.2PnPAL基因結構及進化分析 根據基因組注釋信息中的外顯子和內含子長度分布信息,結合基因結構顯示在線分析工具(GSDS)繪制胡椒PnPAL外顯子、內含子位置示意圖。利用Mega 5.0(http://gsds.cbi.pku.edu.cn/)的鄰接法構建胡椒各PnPAL的親緣關系。
1.2.3 胡椒PnPAL保守基序分析 運用motif 基礎上的序列分析工具(MEME)預測蛋白序列保守基序(http://meme-suite.org/)和美國生物信息技術中心的CDD 工具分析所得domain和TB tools 繪制成圖(https://www.ncbi.nlm.nih.gov/cdd)。
1.2.4 胡椒PnPAL啟動子分析 利用在線數據庫Plant CARE 和TB tools 分析胡椒PnPAL起始密碼子上游2 000 bp 的順式作用元件(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)。
1.2.5 蛋白系統進化分析 運用美國生物信息中心的Blast 工具,獲得注釋信息為苯丙氨酸解氨酶的蛋白序列,并結合擬南芥(Arabidopsis thaliana)、煙草(Nicotiana tabacumL.)、小麥(Triticum aestivumL.)、玉米(Zea mays)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大麥(Hordeum vulgareL.)、菠蘿(Ananas comosus)、香蕉(Musa nana Lour.)、橡膠(Hevea brasiliensis)、棉花(Gossypium arboreum)、油菜(Brassica napusL.)、鵝掌楸(Liriodendron tulipifera)、牛樟(Cinnamomum micranthumf.kanehirae)、鱷梨(Persea americana)、睡蓮(Nymphaea colorata)、無油樟(Amborella trichopoda)等作物的蛋白序列,采用Mega5.0 的鄰接法分析物種間苯丙氨酸解氨酶的親緣關系。數據來源于美國生物信息中心NCBI(https://www.ncbi.nlm.nih.gov/),擬南芥信息網站(https://www.arabidopsis.org/index.jsp),國家水稻數據中心(http://www.ricedata.cn/)。
1.2.6 針刺法接種病原菌 在扦插苗靠近土層的地上部用無菌注射器刺3 個小洞,呈三角形,用打孔器取一塊直徑為1 cm 的菌絲快,將有菌絲的一面覆在刺破的洞上,用浸濕的棉花纏繞固定菌塊,起到保濕和固定的作用。
1.2.7PnPAL表達模式分析 使用Prime primer 5.0設計引物,引物見表1。針刺法在胡椒莖基部接種辣椒疫霉菌(參照1.2.6),以接種病原菌后0、8、12、24、48 h 的胡椒根系為材料,運用Real Time PCR 的方法分析7 條含有完整保守序列的PnPAL表達模式。

表1 實時熒光定量PCR 分析所用引物Table 1 Primers used in RT-PCR analysis
2 結果與分析
2.1 胡椒PnPAL 全基因組鑒定
如表2 所示,根據基因組注釋信息,共鑒定到14 條胡椒苯丙氨酸解氨酶PnPAL基因,不均勻分布在9 條染色體上,其中有4 條位于2 號染色體上,3 條位于6 號染色體上,3 號、5 號、8 號、9 號、18 號、28 號和38 號染色體上各含有1 條。理化性質分析表明,PnPAL所含氨基酸殘基數目在72~736,其中PnPAL14 含有的氨基酸殘基數目最多,PnPAL8含有的氨基酸殘基數目最少,所編碼氨基酸殘基數量均值為500 個。各個PAL 蛋白質分子量在7.374 41~83.431 07 kDa,蛋白質平均分子量為50.933 kDa。胡椒PnPAL編碼的蛋白質等電點在5.76~9.77,平均等電點為7.18,其中PnPAL1、2、3、4、10、13、14 為酸性蛋白(理論等電點pI<7),PnPAL5、6、7、8、9、11、12 為堿性蛋白(理論等電點pI>7)。亞細胞分析表明,PnPAL1、2、3、4、10 位于細胞質中,PnPAL5、7、8、9 位于細胞核中,PnPAL6、11 位于葉綠體中,PnPAL12 位于內膜系統中,PnPAL13、14 位于質膜中。

表2 胡椒PnPAL 基因家族成員的理化性質Table 2 Physiochemical properties of PnPAL family in black peppers
2.2 胡椒PnPAL 基因結構及進化分析
如圖1 所示,運用mega 對鑒定到的14 條PnPAL聚類分析表明,PnPAL4、10、3、13、14、1、2、5、7 聚為一類,PnPAL11、12 聚為一類,PnPAL6、8 聚為一類,表明胡椒PnPAL家族可分為3 個進化單元。運用GSDS 分析基因結構結果表明,家族成員外顯子個數在1~5 個,內含子個數在1~4 個,其中PnPAL2、7、8、12、13 沒有內含子,PnPAL3、4、5、10、14 含有兩個外顯子,PnPAL1 含有3 個外顯子,PnPAL6 含有4 個外顯子,PnPAL11 含有5 個外顯子,平均外顯子個數約為2 個。

圖1 胡椒PnPAL 基因家族基因結構分析Fig.1 Structure of PnPAL family in black peppers
2.3 胡椒PnPAL 保守基序分析
如圖2 所示,在線分析工具MEME 鑒定到8 個保守基序,命名為Motif1~Motif8。在14 條胡椒PnPAL中,每條家族成員所包含motif 0~8 個不等,其中7 條PnPAL所含motif 個數最多,含有完整的8 個motif,3 條不含有motif,1 條含有5 個motif,1 條含有3 個motif。

圖2 胡椒PnPAL motif 分析Fig.2 PnPAL motifs of black peppers
如圖3 所示,NCBI CDD 結果表明,7 條含有PLN02457 結合位點,4 條PnPAL屬于PLN02457 超家族,1 條含有保守結構域NEMP,1 條屬于FNRlike 超家族,1 條屬于bHLH-MYC-N 超家族。

圖3 胡椒PnPAL domain 分析Fig.3 PnPAL domain of black peppers
綜上,7 條含有完整motif 的家族成員均含有PLN02457 結合位點,推測此7 條家族成員為胡椒PnPAL家族成員。
2.4 胡椒PnPAL 啟動子分析
如圖4 所示,胡椒PnPAL基因家族成員除基本的順勢作用元件之外,還包括多個與激素和脅迫有關的順式作用元件,包括低溫響應元件、防御和應激相關元件、光反應元件以及水楊酸、茉莉酸甲酯響應元件等,其中含有脫落酸響應元件的PnPAL最多,達13 條,含有茉莉酸甲酯響應元件的PnPAL次之,有11 條,8 條含有生長素響應元件,5 條含有赤霉素響應元件,5 條含有防御和脅迫相關的元件,4 條含有水楊酸響應元件,4 條含有低溫響應元件,1 條含有機械損傷響應元件,表明PnPAL參與生物和非生物脅迫響應,在植物防御和激素信號轉導方面至關重要。PnPAL1所含順式作用元件最多,達36 個;PnPAL8所含順式作用元件最少,僅為17 個,平均順式作用元件個數約為23 個,表明PnPAL響應多項逆境,其中PnPAL1含有啟動子元件明顯多于其他家族成員,PnPAL1可能響應多項逆境。

圖4 胡椒PnPAL 啟動子分析Fig.4 PnPAL promoter of black peppers
2.5 蛋白系統進化分析
苯丙烷途徑是在裸子植物向被子植物進化過程中形成的,如圖5 所示,本研究將具有完整結構的7個胡椒PnPAL基因與擬南芥(Arabidopsis thaliana)、煙草(Nicotiana tabacumL.)、小麥(Triticum aestivumL.)、玉米(Zea mays)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大麥(Hordeum vulgareL.)、菠蘿(Ananas comosus)、香蕉(Musa nana Lour.)、橡膠(Hevea brasiliensis)、棉花(Gossypium arboreum)、油菜(Brassica napusL.)、鵝掌楸(Liriodendron tulipifera)、牛樟(Cinnamomum micranthumf.kanehirae)、鱷梨(Persea americana)、睡蓮(Nymphaea colorata)、無油樟(Amborella trichopoda)等植物的PAL基因進行進化分析,這些基因主要分為單子葉與雙子葉兩類。然而,胡椒的PnPAL基因均處于雙子葉植物的位置,且與木蘭類的植物親緣關系較為接近。此外,PnPAL1,PnPAL2,PnPAL13,PnPAL14 位于系統發生樹的基部,表明這4 個基因屬于較原始的類型;另一方面,PnPAL3,PnPAL4,PnPAL10 位于離系統發生樹基部較近的位置,且與木蘭類的植物聚在一起。現代植物學家普遍承認“真花學說”,認為木蘭類的植物屬于較原始的類群,因此,胡椒的PnPAL基因較為古老,與胡椒的系統地位相似[14-15]。
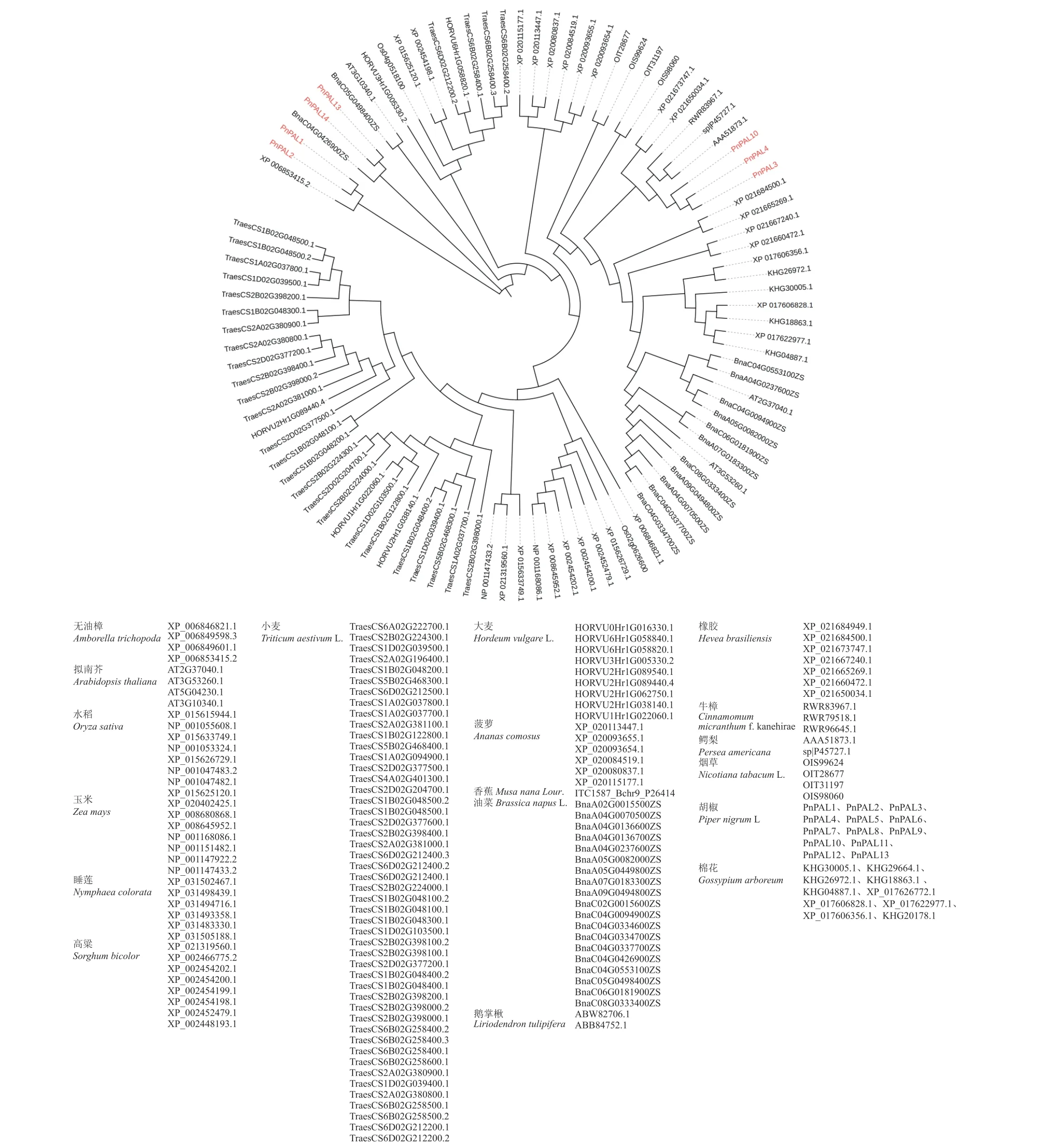
圖5 胡椒PnPAL 系統發育樹
2.6 胡椒PnPAL 表達模式分析
對篩選到完整的7 條PnPAL進行表達模式分析,結果表明PnPAL3、PnPAL10 有表達,其余5 條未檢測到信號。其中PnPAL3 和PnPAL10 在接種后0、8、12、24、48 h 的根系中均有表達。如圖6 所示,PnPAL3 在接種病原菌初期表達相對較高,這可能與植物的應激反應有關。如圖7 所示,PnPAL10表達在處理組中基本呈上調趨勢,特別是在接種病原菌后的24、48 h 顯著高于對照組基因的相對表達,表明該基因可能在病原菌侵染胡椒根系過程中起主要作用。

圖6 胡椒PnPAL3 基因相對表達分析Fig.6 Relative expressions of PnPAL3 in black peppers

圖7 胡椒PnPAL10 基因相對表達分析Fig.7 Analysis on relative expressions of PnPAL10 in black pepper
3 討論
基因組測序技術的不斷深入以及功能基因組學的日漸成熟為基因鑒定提供了便利條件,生物信息學的地位也日益重要。苯丙氨酸解氨酶基因家族應對各項生物和非生物脅迫,目前,葉小真等在桉樹中克隆獲得了PAL基因序列并分析其表達模式,發現該基因在高抗抗焦枯病菌品種中表達最高[16];吳遠航等克隆了木薯的MePAL基因,并發現該基因可受低溫脅迫誘導增強表達[17];高紅勝等克隆了黃瓜的CsPAL基因,發現其在受白粉菌誘導后顯著高于對照[18]。本研究在胡椒中鑒定到14 條PnPAL,其中7 條為完整結構,略多于擬南芥、水稻等家族,可能與胡椒基因組復制有關[19-20]。植物在進化過程中一般都比較保守,保守序列可能調控基因表達且含有相同保守序列的基因行使相同功能,甚至一些屬于相同超家族的蛋白功能也相同[21]。筆者發現7 條胡椒PnPAL均含有保守的基序,推測它們可能行使相同功能。
由一個祖先通過基因復制或突變為適應不同環境而產生一個基因家族[22]。在植物進化過程中,某個祖先的倍增和變異可以產生多個家族成員,這些家族成員可能成簇的分布在一條染色體上,也可能分布在不同染色體上[23]。胡椒的PnPAL家族成員在染色體上不均勻分布,可能與基因組復制及染色體重復有關,基因組和染色體重復對物種進化有重要意義[24]。內含子的非法交換和變異可能是基因家族形成的原因[25],如植物12-氧-植物二烯酸還原酶(12-oxo-phytodienoic acid reductase,OPR)家族基因各內含子的位置和長度存在較大差異,故胡椒PnPAL成員之間基因結構的差異較大[26]。
苯丙氨酸解氨酶普遍存在于植物體中,鑒定到的胡椒PAL 蛋白理化性質、亞細胞定位、酸堿性、分子量、編碼長度等不同,表明胡椒PnPAL有較高的遺傳多樣性。伴隨著物種進化能力逐漸增強,環境適應能力愈強,均是遺傳變異豐富造成的,故遺傳變異是導致遺傳多樣性的根本原因[27]。近年來,越來越多研究表明基因表達調控對基因表達差異起重要作用[28]。在胡椒PnPAL中,各家族成員之間啟動子略有差異,每個PnPAL在不同的生物或非生物脅迫中起作用;家族成員之間啟動子的差異可能是進化過程中遺傳變異導致的;由于各個家族成員所含啟動子不同,啟動子對基因表達的調控又有一定差異,進而表現出PnPAL家族成員參與應對不同的生物和非生物脅迫環境。
胡椒苯丙氨酸解氨酶家族各基因功能還尚不清晰。本文首次運用胡椒基因組信息,鑒定到胡椒的PnPAL家族成員,結合生物信息學方法分析胡椒PnPAL基因家族的結構差異,進化關系及表達模式并篩選到抵抗病原菌的關鍵家族成員,為進一步驗證胡椒PnPAL基因功能及響應各項逆境研究提供基礎。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
