果蠅心臟發育標記基因Lbe抗體的制備與應用
2021-09-09 01:07:14趙夢婧吳秀山江志鋼萬永奇
激光生物學報 2021年4期
關鍵詞:血清
趙夢婧,覃 彬,陳 宇,吳秀山,江志鋼,萬永奇
(湖南師范大學生命科學學院心臟發育研究中心,長沙 410081)
黑腹果蠅(Drosophila melanogaster)是一種應用廣泛的模式生物,擁有許多獨特的優點,如體型小、便于飼養管理、繁殖力強、性狀易于觀察等[1]。果蠅的大多數發育相關基因與人類基因高度保守,近年來其在人類發育與疾病研究領域也揮著重要作用[2]。數據表明,預計到2030年,全球死于心血管疾病的人數將達到2 300萬,占據非傳染性疾病致死人數的44%,是死于癌癥人數的2倍[3-4]。而果蠅心臟發育過程與人類的心臟發育過程高度相似,研究果蠅心臟發育的調控機制將能為人類心臟疾病的治療提供重要的線索[5-7]。
Lbe基因是果蠅同源框基因的成員之一,也是果蠅心臟發育的重要標記基因。它參與了果蠅的心源性途徑,并在果蠅胚胎早期的成心細胞和心臟前體細胞中均有表達,與中胚層Tinman基因、Wg基因和神經基因等共同調控果蠅的心臟發育[8-10]。有研究表明,特異性干擾Lbe將會擾亂心臟中胚層內特定細胞類型的基因表達空間模式,從而導致心臟祖細胞分化與定位紊亂,如Lbe過表達將導致心臟前體細胞的異常增多,而Lbe基因失活將導致心臟循環異常等[10-13]。這表明Lbe的調控對于心臟的正常發育過程是必不可少的。因此,制備針對Lbe的抗體可以更清楚地了解該基因的表達狀況,為研究其在心臟發育過程中的具體功能奠定基礎。
DNA免疫是利用基因重組技術將目的基因與合適的表達載體相連作為抗原,直接免疫機體以表達相應抗體的新技術[14]。與傳統的抗體制備方法相比,DNA免疫具有許多優勢[15-16],如DNA免疫不需要在體外合成蛋白質作為抗原,體內表達的蛋白質可以最大程度地維持天然構象和翻譯后修飾形成的獨特結構,從而有助于針對天然構象抗原的高親和力抗體的產生。基于以上優點,本文選擇通過DNA免疫來制備Lbe抗體。
1 材料與方法
1.1 試驗試劑及材料
試驗所用的大腸桿菌菌株DH5α由本實驗室保種;哺乳動物表達載體pCAGGS-P7由本實驗室提供;限制性內切酶KpnI、XhoI以及2×MIX Taq酶購買自sigma公司;蛋白胨、酵母提取物、氯化鈉、丙烯酰胺、通用胚胎固定液、含有0.5%牛血清蛋白的磷酸鹽緩沖液(phosphate buffered saline with 0.5% bovine serum albumin,PBSBT)購買自生工公司;組織快速裂解液(radio-immunoprecipitation assay lysis buffer,RIPA)和蛋白上樣緩沖液購買自康為試劑公司;RNA試劑盒購買自上海飛捷生物技術有限公司;反轉錄試劑盒購買自Takara公司;無內毒質粒提取試劑盒(離心柱型)購自康為試劑公司;昆明白小鼠由本動物實驗室提供。
1.2 試驗方法
1.2.1 引物設計與合成
在NCBI(https://www.ncbi.nlm.nih.gov/)數據庫中找到果蠅的Lbe基因序列,并找出抗原性最佳的片段,隨后利用Primer5.0 軟件設計產物長度約為440 bp的聚合酶鏈式反應(polymerase chain reaction, PCR)引物。正義引物:5'CTATAGGGCGAATTGGG TACCATGCTCTGCCCTCCAACCAT 3'(劃線部分是KpnI同源臂);反義引物:5'ATCGATACCGTCGACC TCGAGCGCTGCTTCTCCGAGTGACC 3'(劃線部分是XhoI同源臂)。引物由北京擎科生物公司合成。Lbe蛋白的分子量大小約為51 kD。
1.2.2 基因克隆及載體構建
選取w1118野生型果蠅發育后期(10 h)胚胎50枚,使用RNA提取試劑盒提取其RNA,再進行反轉錄,構建cDNA文庫;隨后以cDNA為模板進行PCR擴增,設計梯度為53、56、59、62及65℃的退火溫度,得到擴增長度為440 bp的片段。經純化后將片段與哺乳動物載體pCAGGS-P7連接并轉化至DH5α大腸桿菌中,挑取單克隆進行菌液擴大培養和測序分析。
1.2.3 重組質粒的提取
將測序正確的單克隆擴大培養后,用無內毒素質粒試劑盒進行質粒提取。質粒提取后于-20℃保存。
1.2.4 多克隆抗體的制備
將重組質粒pCAGGS-P7-Lbe免疫4周齡小鼠。分別對體型相似、性別一致的3只小鼠其后肢股四頭肌注射質量濃度為500 ng/μL的重組質粒DNA 40 μL,注射完畢后立即在注射部位兩側5 mm處用細胞融合轉基因儀兩端電極電擊3次,讓質粒DNA盡快擴散均勻。分別在注射7、14、21 d后,按上述方法給這3只小鼠注射同種相同劑量質粒DNA用于加強免疫。于35 d后取血收集血清,將血清分裝后加入等體積濃度為0.05%的疊氮化鈉,于-80℃保存。
1.2.5 多克隆抗體的效價評價
收集的血清用于后續的蛋白質印跡(Western blot)試驗檢測效價。取發育后期(10 h)的w1118野生型果蠅胚胎100枚研磨,加入RIPA組織裂解液后靜置10 min,待溶液澄清后加入蛋白上樣緩沖液約100 μL,上下混勻后煮沸10 min,即得到蛋白樣本。使用前先12 000 r/min離心1 min,取用上清。將Lbe血清抗體稀釋為1∶200、1∶500后,采用Western blot試驗檢測多克隆抗體的特異性。相同條件下,用免疫前的小鼠血清作為對照。在果蠅發育不同時期的試驗中,不同發育時間的胚胎均取用100枚,幼蟲與成蠅均取用1只。
1.2.6 果蠅胚胎固定
收集胚胎后,用毛筆輕輕掃下胚胎于30%的次氯酸鈉溶液中脫殼,約5 min后可在顯微鏡下觀察到胚胎觸角消失;此時用逆滲透(reverses osmosis,RO)水沖洗3~4遍,再將胚胎轉入到含有胚胎固定液(4 mL正庚烷、3 mL通用胚胎固定液、1 mL甲醛)的帶蓋玻璃瓶中;震蕩30 min后,將下層吸棄,并用RO水清洗2遍,再將胚胎吸入到裝有1 mL甲醛的EP管中。
1.2.7 果蠅胚胎抗體染色
將固定好的胚胎加入到1 mL PBSBT中,搖洗1 min后,除去廢液。加入1 mL PBSBT,搖洗10 min,再吸出廢液。加入1 mL PBSBT,在室溫搖洗1 h,進行封閉。封閉完成后,吸棄液體,加入200 μL PBSBT與1 μL一抗,室溫孵育2 h。回收一抗后,加入PBSBT搖洗30 min,隨后按200∶1的體積比混合PBSBT與熒光二抗,避光孵育2 h。最后PBSBT搖洗30 min后即可在顯微鏡下進行觀察。
1.2.8 胚胎染色觀察與圖像處理
果蠅胚胎染色制片后使用蔡司(ZEISS)正置智能顯微鏡 Axio Imager M2 進行觀察拍攝,拍攝所得圖片使用Photoshop CS6軟件進行標注編輯。
2 結果與分析
2.1 目的片段的克隆及重組載體鑒定
利用添加同源臂后設計的引物進行PCR。為了找到最適PCR退火溫度,本研究設計了53、56、59、62及65℃ 5個溫度梯度。PCR結果如圖1所示。PCR擴增條帶均一且位置符合預期,表明目的片段成功擴增。將目的片段純化后與pCAGGS-P7質粒同源重組,構建如圖2所示的pCAGGS-P7-Lbe重組載體。使用KpnI和XhoI對重組質粒進行雙酶切,電泳結果顯示,所得條帶大小分別與載體和目的片段一致(圖3)。對單克隆菌液進行擴大培養后測序,序列比對結果表明pCAGGS-P7-Lbe載體構建成功(圖4)。序列比對發現重組載體中有3處點突變,分別位于171、371和431 bp處,但3處突變均為同義突變(CCG-CCA-脯氨酸;GTT-GTC-纈氨酸),不影響氨基酸序列。
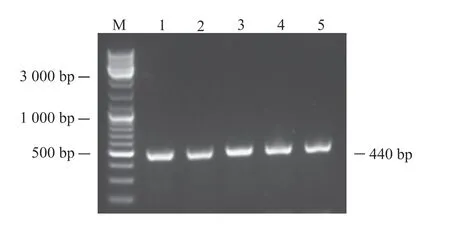
圖1 Lbe基因 PCR擴增的電泳結果Fig. 1 PCR amplification and electrophoresis result of Lbe geneM:DNA marker;1:53℃退火溫度時PCR擴增條帶;2:56℃退火溫度時PCR擴增條帶;3:59℃退火溫度時PCR擴增條帶;4:62℃退火溫度時PCR擴增條帶;5:65℃退火溫度時PCR擴增條帶。M: DNA Marker; 1: PCR amplification band at 53℃ annealing temperature; 2: PCR amplification band at 56℃ annealing temperature; 3: PCR amplification band at 59℃ annealing temperature; 4: PCR amplification band at 62℃ annealing temperature; 5: PCR amplification band at 65℃ annealing temperature.
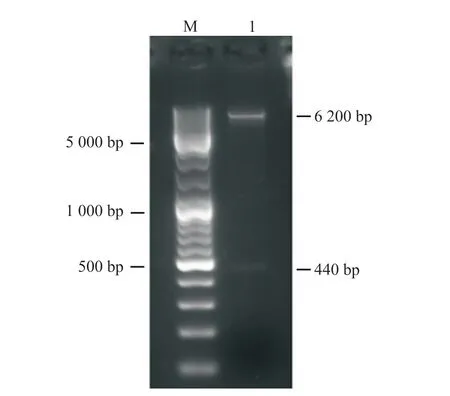
圖3 pCAGGS-P7-Lbe重組質粒Kpn I和Xho I雙酶切的電泳結果Fig. 3 Double digested with Kpn I and Xho I and electrophoresis result of pCAGGS-P7-Lbe recombinant plasmidM:DNA Marker;1:pCAGGS-P7-Lbe重組質粒Kpn I和Xho I雙酶切的產物。M: DNA marker; 1: The recombinant plasmid pCAGGS-P7-Lbe was double digested with Kpn I and Xho I.

圖4 pCAGGS-P7-Lbe重組質粒測序的比對結果Fig. 4 Sequencing comparison result of recombinant plasmid pCAGGS-P7-Lbe紅色方框內為3處同義突變(CCG-CCA-脯氨酸;GTT-GTC-纈氨酸)。Three synonymous mutations were identified in the red box (CCG-CCA-proline; GTT-GTC-valine).
2.2 Lbe多克隆抗體的Western blot鑒定
將免疫后小鼠血清作為一抗針對野生型果蠅胚胎全蛋白進行Western blot試驗,結果顯示,該血清的全蛋白檢測中只出現了一條蛋白帶(圖5a),且大小與預計的Lbe蛋白大小接近,表明該血清的特異性較好,無交叉反應。隨后,對該血清使用的最佳稀釋比例進行摸索,分別以1∶200、1∶500兩種稀釋比例進行Western blot試驗(圖5b、5c),結果顯示當稀釋比例為1:200時,條帶清晰且背景干凈(圖5b),表明1:200為該抗血清的適宜稀釋比例。所有Western blot試驗均以β-actin作為內參(圖5d)。
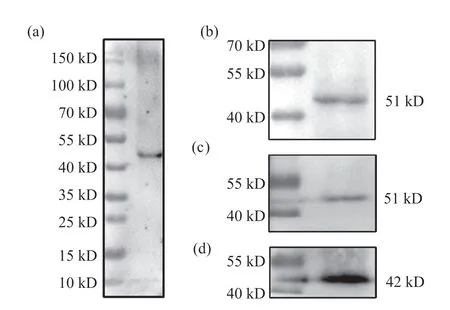
圖5 抗Lbe血清特異性與適宜稀釋比例檢測結果Fig. 5 Specificity and appropriate dilution ratio detection results of anti-Lbe serum(a)抗Lbe血清的全蛋白檢測結果;(b)血清稀釋比例為1∶200時的Western blot結果;(c)血清稀釋比例為1∶500時的Western blot結果;(d)β-actin抗體稀釋比例為1∶1 000時的Western blot結果。(a) Whole protein detection result of anti-Lbe serum; (b) Western blot result when the dilution ratio of serum is 1∶200; (c) Western blot result when the dilution ratio of serum is 1∶500; (d) Western blot result when the dilution ratio of anti-β-actin is 1∶1 000.
2.3 Lbe蛋白在果蠅不同發育階段的表達狀況
為了深入了解Lbe蛋白在果蠅不同發育階段的表達情況,我們利用制備的抗體檢測了w1118果蠅不同時期的胚胎中Lbe蛋白的表達水平。結果顯示,在胚胎中,Lbe在4 h時開始表達,在7 h和10 h時表達不斷增強,但在13 h和16 h時表達有所下降(圖6a)。我們認為這可能是由于Lbe基因在胚胎發育早期發揮了作用,在果蠅胚胎生殖帶伸長時期便有表達,隨后參與中胚層心臟前體細胞與神經系統的發育,故在此期間表達量不斷增加,而在果蠅胚胎中線閉合后表達下降。為了驗證上述猜想,我們選擇了果蠅發育的3個不同時期,即胚胎、幼蟲和成蟲,檢測這3個時期Lbe蛋白的表達水平。結果表明,在胚胎階段,Lbe蛋白的表達量較高,而在幼蟲階段表達明顯減少,在成蠅中基本檢測不到Lbe的表達(圖6b)。這與我們的猜想相一致。上述結果提示,Lbe基因在果蠅胚胎早期發育過程中發揮了重要作用,我們制備的抗體能夠準確反映Lbe蛋白的表達水平。
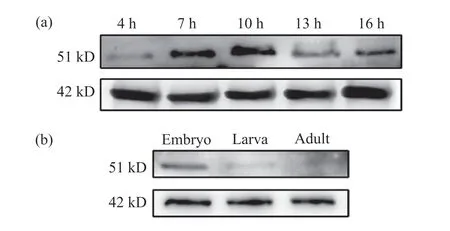
圖6 果蠅不同發育時期的Lbe蛋白表達情況Fig. 6 Expression of Lbe protein in Drosophila melanogaster at different developmental stages(a)胚胎發育到4、7、10、13和16 h時 Lbe的蛋白表達;(b)果蠅胚胎、幼蟲、成蠅階段的Lbe蛋白表達。(a) The expression level of Lbe protein in Drosophila melanogaster embryo at 4, 7, 10, 13 and 16 h; (b) The expression of Lbe protein in Drosophila melanogaster embryo, larva and adult stages.
2.4 胚胎抗體染色
為了檢測抗體能否用于免疫染色,我們用血清對不同發育時期的果蠅胚胎進行免疫染色,發現在stage 8時期,Lbe基因的表達集中在胚胎的肛板處(圖7b)。隨著發育的進行,到stage 10時期,Lbe基因的表達在胚胎內開始增加,主要集中在胚胎觸角段、上下頜節段和唇部等部位,并且在胚胎邊緣的神經母細胞中也能檢測到Lbe的表達,肛板處的表達仍然強烈(圖7c)。stage 11時期,Lbe基因的表達相較上一時期有了明顯回縮的趨勢,集中在上下頜節段中。值得注意的是,在位于表皮內側的成心前體細胞中能檢測到較強的Lbe表達信號(圖7d)。這表明Lbe基因確實參與了成心前體細胞的發育。而在胚胎發育末期,在節段邊界肌形成細胞中與表皮內側能檢測到廣泛的Lbe蛋白陽性信號,并且表達Lbe蛋白的表皮內側細胞的寬度也有所增加(圖7e)。在stage 15時期,表達Lbe蛋白的成心前體細胞有向背中線遷移的趨勢(圖7f)。這也證明了Lbe的正確表達與果蠅心管的形成密不可分。上述免疫染色結果中Lbe陽性信號與其發育規律高度符合,表明我們制備的抗體能夠精確檢測Lbe蛋白的表達定位,可用于胚胎抗體染色,且效果良好。
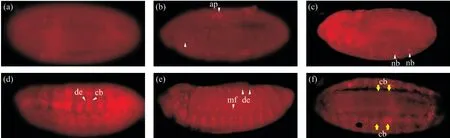
圖7 Lbe多克隆抗體胚胎染色鑒定(×200)Fig. 7 Immunofluorescence identification of Lbe polyclonal antibody (×200)(a)空白血清對照組;(b)stage 8時期的野生型果蠅胚胎(發育時間為3 h 30 min);(c)stage 10時期的野生型果蠅胚胎(發育時間為4 h 20 min);(d)stage 11時期的野生型果蠅胚胎(發育時間為7 h);(e)stage 15時期的野生型果蠅胚胎(發育時間為13 h);(f)stage 13時期的野生型果蠅胚胎(發育時間為10 h)的背觀圖。圖中的胚胎均按照頭部向左,腹部向下的姿勢擺放。ap:肛板;nb:神經母細胞;de:表皮內側;cb:成心前體細胞;mf:節段性邊界肌形成細胞。黃色箭頭所示均為細胞類型和特定細胞區域;白色箭頭所示為細胞運動趨勢。(a) Blank serum control group; (b) Wild type Drosophila embryos at stage 8 (development time 3 h 30 min); (c) Wild type Drosophila embryos at stage10 (development time 4 h 20 min); (d) Wild type Drosophila embryos at stage 11 (development time 7 h); (e) Wild type Drosophila embryos at stage 15 (development time 13 h); (f) Wild type Drosophila embryos at stage 13 (development time 10 h). All whole-mounts are oriented with the anterior to the left; (f) is the back view. ap: Analplate; nb: Mandibular segment; de: Dorsal epidermis; cb: Cardioblast precursors; mf: Segmental border muscle founder cells. yellow arrow shows cell type and specific cell area; white arrow shows cell movement trendency.
3 討論
心臟作為身體內最重要的器官之一,從早期發育階段就擔負著全身循環的功能。在脊椎動物和無脊椎動物中,心臟起源于雙側中胚層的原基細胞,并且越來越多的證據表明,心臟譜系的分子控制機制在生物進化過程中是保守的。果蠅雖然在進化上離脊椎動物較遠,但它的心管在形態上類似于脊椎動物心臟發育早期的線性管,其心管發育基因的同源物也在脊椎動物心臟發育中發揮重要調控功能。因此,本文選擇果蠅為模式生物來研究心臟發育。Lbe基因會在果蠅特定的成心細胞亞群中表達,并在心臟前體的多樣化中發揮作用。已有研究表明,當Lbe基因被敲除表達時,果蠅心臟前體細胞的數量會減少,而過度表達Lbe將會導致果蠅表現出心臟結構缺陷,以及心律失常、心臟停搏等癥狀[17-18]。Lbe基因在進化上是保守的,在小鼠和人類中已經發現同源基因[13]。
果蠅與人體的心臟發育基因具有高度保守性,故研究果蠅心臟發育基因是探索人類心臟發育基因的一條有效途徑。但由于市面上的果蠅心臟基因抗體難以購得,導致果蠅心臟發育基因研究缺少合適的工具。本文通過DNA免疫技術成功制備了可用于Western blot和胚胎染色的果蠅Lbe基因的抗體。雖然與原核表達載體相比,血清抗體的效價略低,但DNA免疫的獨特優點(如可以直接免疫小鼠、方法更加簡便、耗時更少等)使其成為快速制備分子量較小的蛋白抗體的有效途徑之一[15-17]。Western blot和免疫染色結果表明,所制備抗體的特異性和敏感性都較好,這為將來運用免疫共沉淀等方法研究Lbe基因調控果蠅心臟發育的分子機制奠定了基礎。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
