多酚氧化酶的性質及其在茶葉加工中的研究進展
2021-09-10 07:22:44鄒純尹軍峰
中國茶葉 2021年6期
關鍵詞:應用
鄒純 尹軍峰
摘要:多酚氧化酶是茶葉加工中一類重要的氧化還原酶。文章歸納了多酚氧化酶的結構性質和酶學性質,重點闡述了其在紅茶、綠茶、烏龍茶和黑茶加工中的研究現狀,并展望了其未來的研究方向,以期為深入研究和應用多酚氧化酶提供參考。
關鍵詞:多酚氧化酶;茶葉加工;結構性質;酶學性質;應用
The Properties of Polyphenol Oxidase and Its
Research Progress in Tea Processing
ZOU Chun, YIN Junfeng*
Tea Research Institute, Chinese Academy of Agricultural Sciences, Hangzhou 310008, China
Abstract: Polyphenol oxidase is an important oxidoreductase in tea processing. This paper summarized the structural?and enzymatic properties of polyphenol oxidase, emphasized its research status in the processing of black tea, greentea, oolong tea and dark tea, and prospected its future research directions, in order to provide a reference for in-depthresearch and application of polyphenol oxidase.
Keywords: polyphenol oxidase, tea processing, structural properties, enzymatic properties, application
多酚氧化酶(Polyphenol oxidase,PPO)廣泛存在于植物、動物、真菌體內,屬于氧化還原酶,在廣義上根據催化底物酚羥基數量的不同,可分為三大類[1]:單酚氧化酶(酪氨酸酶,EC 1.14.18.1)、雙酚氧化酶(兒茶酚氧化酶,EC 1.10.3.1)和漆酶(EC 1.10.3.2)。植物中的PPO主要是兒茶酚氧化酶,能夠在有氧條件下催化多酚類物質生成對應的醌類。
PPO在茶葉加工中具有重要作用,通過不同加工工序或工藝抑制或提升PPO活性,可制備出風味迥異的各類茶。本文對多酚氧化酶的性質及其在茶葉加工中的研究現狀進行了綜述,展望其未來的研究方向,以期為深入研究和應用多酚氧化酶提供參考。
一、PPO的結構與性質
1. PPO的結構
(1)蛋白結構
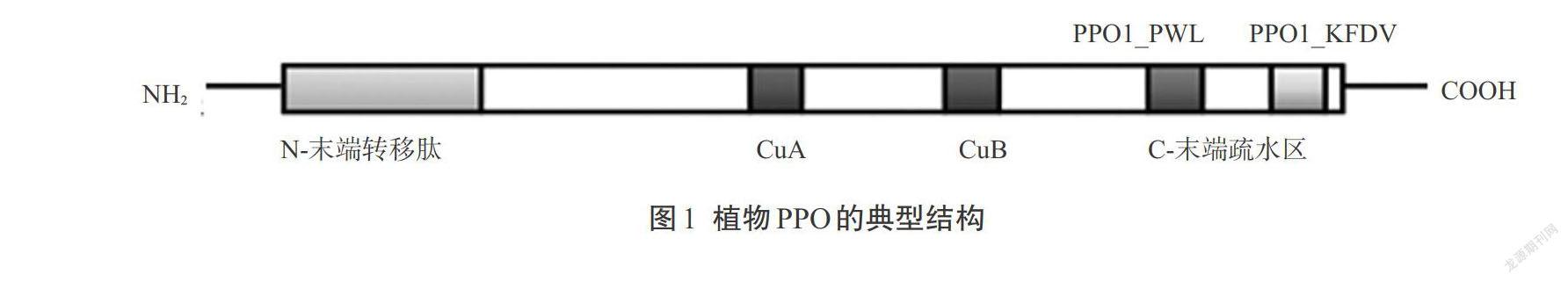
植物PPO通常先以無活性的前體形式存在,其典型結構主要分為3個部分:N-末端轉移肽、中間銅離子結合區、C-末端疏水區(圖1)[2]。N-末端轉移肽的大小通常為8~12 kDa,大部分為葉綠素轉移肽,其作用是將PPO前體蛋白轉運至葉綠體類囊體腔,在成熟的PPO中轉移肽已被去除[3]。中間銅離子結合區是維持PPO催化活性所必須的,通常由CuA和CuB兩個銅離子活性中心組成,每個銅離子活性中心分別由1個銅離子和3個組氨酸殘基以配位鍵相連,形成具有特定三維結構的活性部位。CuA和CuB結構域在不同植物種類中表現出高度保守性,但是CuB還具有一定的變異性,這種變異性可能造成不同PPO對底物的偏向性。C-末端疏水區有兩個保守的結構域PPO1_PWL和PPO1_KFDV,其功能目前尚不明確,但是有研究者認為它可以通過遮擋酶活性中心的“催化口袋”來控制PPO的催化活力,切除該結構域可能對PPO起激活作用[4]。
研究發現,PPO在體外實驗中可以被多種方式激活[5]:蛋白酶有限水解、酸或堿處理、加熱處理、老化(失水)作用、強洗滌劑(SDS)或者醇處理。Nozue等[6]研究發現,在甘薯中存在兩種不同亞型的PPO:分子量為40 ku的高活性PPO和分子量為60 ku的低活性PPO;在絲氨酸蛋白酶的作用下,低活性PPO可被水解為高活性形式。Gandia-Herrero等[7]通過SDS對甜菜根PPO前體酶進行體外激活,發現PPO前體酶在激活過程中活性中心沒有受到SDS的影響,而是蛋白構象逐漸改變成利于疏水性底物進入。Kampatsikas等[8]研究發現,蘋果PPO前體酶的C-末端結構域遮擋了其活性中心,影響其催化活性;而在C-末端結構域與活性中心之間的Lys355-Val370連接序列可發生自我斷裂,將活性中心暴露,使底物更容易進入其活性中心,從而實現PPO的自我激活。由此可知,PPO酶蛋白結構和構象對其催化活性起決定作用。
(2)基因結構
植物PPO由核基因編碼表達,在植物的光合組織以及非光合組織質粒的膜上或膜腔中都有分布[9]。隨著分子生物學技術的成熟和廣泛應用,許多植物如蘋果、梨、桃、荔枝、甘薯、西紅柿、蠶豆、馬鈴薯、葡萄、菠菜、甘蔗、煙草、杏、枇杷等的ppo基因都已被克隆[10]。目前研究發現,ppo基因在單子葉植物中普遍存在內含子,而在大部分雙子葉植物中沒有內含子,僅在番荔枝[11]、白楊[12]等少數雙子葉植物中發現內含子的存在。2001年,趙東等[13]根據已報道的ppo基因中的保守序列設計引物,首次克隆了茶樹ppo基因部分序列;通過序列分析發現茶樹ppo基因與其他植物的ppo基因同源性較高,尤其是銅離子結合部位。通過檢索最新NCBI數據庫發現,目前已有宜紅早、碧螺春、安徽1號等茶樹品種的17條ppo基因完整或部分序列公布,大小為537~1 800 bp。在茶樹ppo基因中均未發現內含子。
植物ppo基因具有多基因家族特性,在不同種類植物和同種植物不同器官中具有不同的時空表達模式。Chi等[14]通過分析馬鈴薯基因組數據得到9個可能的ppo基因,通過實時定量PCR發現,這些基因不僅在不同組織中的分布和表達量存在較大差異,而且在不同品種中的分布也不同。曾澤媛等[15]通過茶樹基因組數據庫中獲得了5條ppo基因,它們的CDS區全長為597 ~ 1 839 bp,編碼氨基酸數目為198 ~ 612個;通過實時定量PCR發現,這些基因在不同品種間呈現出不同表達模式。
2. 酶學性質
(1)底物特異性
PPO能夠催化一元酚、二元酚和多元酚等酚類物質氧化形成相應的醌,且對不同類型的底物催化效率差異較大。常用于PPO催化的底物包括鄰苯二酚、焦性沒食子酸、兒茶素、沒食子酸等,其中鄰苯二酚常用于茶樹PPO酶活測定。Samynathan等[16]分離純化了UPASI品種中的PPO,研究其底物特異性發現,最適底物為兒茶素,然后依次是鄰苯二酚、咖啡酸、L-多巴和連苯三酚,而對沒食子酸不起催化作用。伍夢瑤等[17]研究了勐庫大葉種PPO的催化特性,發現它對焦性沒食子酸、鄰苯二酚有催化活性,且對焦性沒食子酸的催化效果優于鄰苯二酚,而對間苯二酚、對苯二酚基本無催化活性。虞昕磊[18]研究發現,鳳慶PPO的最適底物為鄰苯二酚和焦沒食子酸,可部分氧化沒食子酸和對苯二酚,但對間苯二酚催化作用最弱。
(2)pH
PPO的最適pH受到酶來源和選用底物影響。根據資料報道[19],茶樹PPO適宜的反應pH為4.0 ~ 9.0,不同茶樹品種PPO或同一品種的不同同工酶的最適pH值存在一定差異。?ztürk等[20]以鄰苯二酚和4-甲基鄰苯二酚兩種底物測定了茶鮮葉PPO的最適pH,分別為9.0和8.5。Teng等[21]從政和大白茶中純化出PPO1和PPO2兩種同工酶,其最適pH分別為5.5和6.0。許雷[22]純化出龍井43中的兩種PPO同工酶,以鄰苯二酚為底物確定兩者的最適pH分別為5.6和6.8。
(3)溫度
不同來源PPO的活性隨溫度變化的趨勢具有相似性,一般隨溫度升高先上升后下降,但其最適溫度存在差異。茶樹PPO最適反應溫度在20~40 ℃。Samynathan等[16]分析了UPASI品種中PPO的酶學性質,發現其最適溫度為25 ℃,在32 ℃下十分穩定,而在62 ℃下僅15 min就完全失活。Altunkaya[23]純化了茶鮮葉中的PPO,研究確定其最適溫度為30 ℃,并發現其在30 ~ 80 ℃范圍內,熱穩定性隨溫度升高逐步下降。許雷[22]分析了龍井43中的兩種PPO同工酶催化活性和溫度的關系,發現它們的最適溫度分別為30 ℃和35 ℃;在25~40 ℃范圍內,二者活性均較高,而溫度超過60 ℃后,二者基本失活。
(4)金屬離子
植物PPO屬于金屬酶,其催化活性受金屬離子的種類和濃度影響。Samynathan等[16]研究了2 mmol/L金屬離子對茶樹PPO活性的影響,發現K+、Na+、Mg2+和Cu2+具有激活作用,而Hg2+和Ca2+具有抑制作用,其中Hg2+表現出強抑制作用,可抑制90%的酶活力。張元等[24]研究發現,Cu2+在低濃度下對茶鮮葉中PPO具有激活作用,而在高濃度下起抑制作用;Cu2+濃度為0.25×10-7? mol/L時PPO酶活性最高,當Cu2+濃度達2.5×10-7 mol/L時PPO酶活性消失。
二、PPO在茶葉加工中的研究概況
1. 紅茶
紅茶屬于全發酵茶,在其加工過程中通過酶促氧化兒茶素類物質產生茶黃素類色素,并形成其特有的色澤和香氣。PPO是酶促氧化中的關鍵酶,在紅茶加工中其酶活性處于動態變化。紅茶加工工序主要包括:萎凋、揉捻、發酵和干燥。萎凋是紅茶加工的首道工序,該階段葉片細胞萎縮、細胞汁濃度提高,部分水解酶、氧化酶活性上升。豐金玉等[25]研究發現,PPO在萎凋期間活性隨萎凋時間的延長而增加,萎凋結束時PPO活性達到鮮葉PPO活性的2.9倍。揉捻通過外力作用使葉片迅速緊卷形成茶條,破損葉細胞組織,使葉片中的多酚類化合物、蛋白質、氨基酸等物質溢聚葉表皮,與PPO、氧氣等充分接觸,促進生化變化。Mahanta等[26]研究發現,PPO活性在揉捻過程中達到最高。發酵是形成紅茶品質特征的關鍵工序,實質是以多酚類化合物為主體發生酶促或非酶促氧化反應的化學變化過程。紅茶發酵條件對PPO活性有較大影響:高濕有利于PPO酶活性的保持;缺氧會使PPO的含Cu2+活性中心呈脫氧態形式,不利于酶活性的保持和多酚類物質的酶促氧化。Samanta等[27]研究了不同溫度對紅茶發酵的影響,在不同溫度下PPO活性均隨發酵進行明顯下降。干燥是紅茶加工的最后一道工序,不僅能迅速鈍化各種酶的活性、固定成茶外形,還能通過高溫熱作用來激化高沸點的芳香物質,使內含成分發生微妙變化,形成紅茶特有的品質特征。
根據資料報道,在紅茶加工中,PPO不僅活性發生變化,而且酶組成和性質也有較大改變。Takeo[28]研究發現,PPO活性從萎凋階段到揉捻階段持續上升,且在揉捻階段活性達到了鮮葉活性的2~3倍,而在發酵過程中由于形成不溶性多酚-酶復合體,PPO活性下降。進一步的研究還發現萎凋中有新的PPO酶蛋白出現。屠幼英[29]的研究也得到了類似的結論,通過酶學性質分析提出鮮葉與萎凋葉 PPO 有著質的區別。Ozdemir等[30]研究發現,在揉捻中有不同分子量的PPO形成,對氧化反應和紅茶品質起關鍵作用。
在紅茶加工中,提升PPO活性是改善紅茶品質的有效途徑。Jiang等[31]將不同品種、季節和地區的茶鮮葉加工成工夫紅茶,發現具有高PPO活性的茶鮮葉加工的紅茶中茶黃素含量較高。Das等[32]以提高PPO和過氧化物酶(POD)活性為目標,優化了萎凋溫度、濕度以及持續時間,發現優化后對CTC紅茶和傳統紅茶的品質均有明顯提升。Hua等[33]將新型動態發酵方法應用于工夫紅茶加工中,發現相比傳統發酵方式,該方法通過增加發酵過程中的氧含量有效提高PPO活性,進而促進茶黃素和茶紅素形成并改善紅茶的品質。此外,在紅茶加工中添加水果、蔬菜等來源的PPO促進發酵,也能改善其湯色和香氣,并提高茶黃素含量。
2. 綠茶
綠茶是不發酵茶,其關鍵工序是殺青。殺青過程主要通過高溫使鮮葉中包括PPO在內的各種內源酶迅速變性失活,使成品茶保留較多的葉綠素、多酚類、維生素C等品質成分,從而形成綠茶“清湯綠葉”的品質。
吳本剛等[34]將催化式紅外殺青技術應用于綠茶殺青,發現該技術不僅能鈍化茶鮮葉中90%的PPO活力,而且能去除50%左右的水分,此外,還能使殺青葉保持較高的維生素C和茶多酚含量。鄭文佳等[35]通過滾筒加微波疊殺技術鈍化茶鮮葉中大部分PPO活性,顯著延長了殺青葉貯存時間,以此降低綠茶加工的勞動強度和成本。譚俊峰[36]研究了超高壓處理對茶鮮葉的影響,發現經500 MPa以上超高壓處理的茶鮮葉中PPO活性顯著下降,在殺青前對茶鮮葉進行超高壓處理,制得的成品綠茶湯色黃綠明亮,滋味醇和,其香氣物質芳樟醇含量較對照提高15倍。
3. 烏龍茶
烏龍茶做青是搖青和靜置多次反復交替的過程,搖青使葉片受到一定機械損傷,促進細胞中的內源酶與底物接觸,該過程茶青邊緣受損程度更高,在PPO等的酶促氧化下引起葉緣紅變,達到“綠葉紅鑲邊”的效果。靜置主要促進葉片內物質緩慢氧化形成烏龍茶特有品質成分。烏龍茶加工須嚴格控制好做青程度,若做青不足,不僅酶促反應不夠充分,而且葉片和莖梗中內含物不能充分交換,導致茶葉香氣前體不能充分釋放;做青過度會導致酶促反應過度,致使部分香氣物質含量降低,而且還會破壞茶多糖等活性物質。
烏龍茶加工中的溫濕度等環境因子與做青工藝對PPO活性和茶葉品質有重要影響。王麗霞等[37]研究了萎凋條件對烏龍茶鮮葉PPO活性的影響,發現相比自然萎凋(溫度25 ℃,相對濕度50%~60%),低溫(20 ℃)和高濕(相對濕度90 %)條件下萎凋PPO活性更高。王承莉等[38]以PPO比活力和粗多糖質量分數為指標對烏龍茶機械化做青工藝進行了研究,發現投葉量為20 g、溫度30 ℃、振搖時間20 min、靜置時間10 min、重復2次時,葉片適度發酵、香氣良好,有利于形成烏龍茶品質特征。陳倩蓮等[39]研究比較了采用炭火和暖氣機組兩種升溫方式對大紅袍做青中PPO活性的影響,發現兩種升溫方式做青后葉片PPO活性均顯著下降,但暖氣機組升溫能改善做青環境,延長做青過程中青葉的活力,促進香氣物質的轉化生成。
4. 黑茶
黑茶渥堆過程中,微生物大量繁殖,分泌PPO、蛋白酶、纖維素酶等胞外酶,與茶葉的內源酶一起酶促作用,形成黑茶特有的風味品質。
目前,研究者對黑茶渥堆中PPO活性和種類變化展開了系統研究,并分析了PPO對其品質的影響。柴潔等[40]以云南臨滄鳳慶大葉種曬青毛茶為原料進行普洱茶固態發酵,發現渥堆過程中PPO活性與比活力均呈波浪式上升趨勢,發酵末期PPO活性較初期增強,而酶蛋白含量總體呈下降趨勢,但變化比較平穩。胡沛然[41]對六堡茶渥堆前后的PPO進行分析發現,渥堆前毛茶PPO分子量為59 kDa左右,而渥堆結束的樣品出現了分別約為59 kDa和70 kDa的兩種PPO,其原因可能是真菌在渥堆過程中產生的新的PPO。付贏萱等[42]在普洱茶渥堆中添加外源PPO作用發現,PPO能夠加速普洱茶發酵,縮短發酵周期,并改善其品質。
三、展望
盡管目前關于茶樹PPO的研究與應用已取得了諸多顯著成果,但在以下幾個方面仍值得深入研究。
1. 茶樹PPO蛋白質結構的解析
目前研究者已成功純化了多個品種茶樹中的PPO,若能進一步解析其蛋白結構,將有助于深入探明其催化機制。
2. 茶葉加工工藝對PPO活性影響的機制
當前的研究主要分析不同加工工藝對PPO酶活力的影響,深入探明其影響機制將有助于更科學地控制各工藝參數。
3. PPO的高效重組表達
當前制約PPO廣泛應用的主要因素是難以獲得低成本的高活性酶,通過基因工程手段實現PPO的高效重組表達將大幅度拓展其應用范圍。
參考文獻
[1] MAYER A M. Polyphenol oxidases in plants and fungi: Going places?? A review[J]. Phytochemistry, 2006, 67 (21): 2318-2331.
[2] 王麗, 王萬興, 索海翠, 等. 植物中多酚氧化酶基因研究進展[J]. 分子植物育種, 2020, 18 (14): 4629-4636.
[3] TRAN L T, TAYLOR J S, CONSTABEL C P. The polyphenol oxidase
gene family in land plants: Lineage-specific duplication and expansion[J]. Bmc Genomics, 2012, 13 (1): 1-13.
[4] KAINTZ C, MAURACHER S G, ROMPEL A. Type-3 copper proteins: recent advances on polyphenol oxidases[J]. Metal-Containing Enzymes, 2014, 97: 1-35.
[5] IQBAL A, MURTAZA A, HU W F, et al. Activation and inactivation mechanisms of polyphenol oxidase during thermal and non-thermal methods of food processing[J]. Food and Bioproducts Processing,? 2019, 117: 170-182.
[6] NOZUE M, ARAKAWA D, IWATA Y, et al. Activation by proteolysis in vivo of 60-ku latent polyphenol oxidases in sweet potato cells in
suspension culture[J]. Journal of Plant Physiology, 1999, 155 (3):? 297-301.
[7] GANDIA HERRERO F, JIMENEZ ATIENZAR M, CABANES J,? et al. Differential activation of a latent polyphenol oxidase mediated
by sodium dodecyl sulfate[J]. Journal of Agricultural and Food? Chemistry, 2005, 53 (17): 6825-6830.
[8] KAMPATSIKAS I, BIJELIC A, PRETZLER M, et al. A peptide-induced self-cleavage reaction initiates the activation of tyrosinase[J].? ?Angewandte Chemie-International Edition, 2019, 58 (22): 7475-7479.
[9] 劉芳, 趙金紅, 朱明慧, 等. 多酚氧化酶結構及褐變機理研究進展[J]. 食品研究與開發, 2015, 36 (6): 113-119.
[10] 周仲華, 張達, 倪賀, 等. 生物信息學篩選催化茶黃素合成的多酚氧化酶[J]. 華南師范大學學報(自然科學版), 2017, 49 (3): 59-67.
[11] PRIETO H, UTZ D, CASTRO A, et al. Browning in annona cherimola fruit: role of polyphenol oxidase and characterization of a coding sequence of the enzyme[J]. Journal of Agricultural and Food Chemistry, 2007, 55 (22): 9208-9218.
[12] TRAN L T, CONSTABEL C P. The polyphenol oxidase gene family ? in poplar: phylogeny, differential expression and identification of ? a novel, vacuolar isoform[J]. Planta, 2011, 234 (4): 799-813.
[13] 趙東, 劉祖生, 奚彪. 茶樹多酚氧化酶基因的克隆及其序列比較[J]. 茶葉科學, 2001, 21 (2): 94-98.
[14] CHI M, BHAGWAT B, LANE W D, et al. Reduced polyphenol ? oxidase gene expression and enzymatic browning in potato (Solanum tuberosum L.) with artificial microRNAs[J]. Bmc Plant Biology,? ? 2014, 14 (62): 1-18.
[15] 曾澤媛, 羅勇, 鄭楚楚, 等. 茶樹PPO基因家族的鑒定與分析[J]. 茶葉科學, 2018, 38 (4): 385-395.
[16] SAMYNATHAN R, PALANISAMY C P, GANDHI S, et al. Isolation and characterization of polyphenol oxidase from UPASI selected clone of Camellia sinensis (L.) O. Kuntze[J]. Indo American Journal of Pharmaceutical Research, 2015, 5: 241-252.
[17] 伍夢瑤, 黃瑩捷, 姚燕妮, 等. 勐庫大葉種茶樹多酚氧化酶粗酶 ?的酶學性質[J]. 浙江大學學報(農業與生命科學版), 2017, 43 ? (5): 579-588.
[18] 虞昕磊. 云南大葉種茶樹多酚氧化酶的分離純化與性質研究[D]. 武漢: 華中農業大學, 2015.
[19] 王麗霞, 陸蒸, 林啟訓, 等. 福云6號茶葉茶多酚氧化酶特性的研究[J]. 江蘇農業科學, 2008 (1): 213-215.
[20] ?ZTüRK C, AKSOY M, KUFREVIOGLU O I. Purification of tea ? leaf (Camellia sinensis) polyphenol oxidase by using affinity chro- ? matography and investigation of its kinetic properties[J]. Journal of? ? Food Measurement and Characterization, 2020, 14 (1): 31-38.
[21] TENG J, GONG Z H, DENG Y L, et al. Purification, characterization and enzymatic synthesis of theaflavins of polyphenol oxidase isozymes from tea leaf (Camellia sinensis)[J]. Lwt-Food Science and Technology, 2017, 84: 263-270.
[22] 許雷. 茶樹多酚氧化酶的提取、分離純化及其部分酶性質研究[D]. 武漢: 華中農業大學, 2014.
[23] ALTUNKAYA A. Partial purification and characterization of polyphenoloxidase from Turkish tea leaf (Camellia sinensis L.)[J]. International Journal of Food Properties, 2014, 17 (7): 1490-1497.
[24] 張元, 錢文俊, 韓永濤, 等. HPLC分析Cu2+對茶酶源酶促氧化合成茶黃素的影響[J]. 西北農業學報, 2015, 24 (10): 118-123.
[25] 豐金玉, 劉昆言, 秦昱, 等. 紅茶加工中多酚氧化酶、過氧化物酶和β-葡萄糖苷酶活性變化[J]. 農學學報, 2014, 4 (11): 96-99, 113.
[26] MAHANTA P K, BORUAH S K, BORUAH H K, et al. Changes of? ? polyphenol oxidase and peroxidase activities and pigment compo- ?sition of some manufactured black teas (Camellia sinensis L.)[J].? ? Journal of Agricultural and Food Chemistry, 1993, 41 (2): 272-276.
[27] SAMANTA T, CHEENI V, DAS S, et al. Assessing biochemical ? changes during standardization of fermentation time and temperature ? for manufacturing quality black tea[J]. Journal of Food Science and? ? Technology, 2015, 52 (4): 2387-2393.
[28] TAKEO T. Tea leaf polyphenol oxidase: Part III. Studies on the? ? changes of polyphenol oxidase activity during black tea manufacture ? [J]. Agricultural and Biological Chemistry, 1966, 30 (6): 529-535.
[29] 屠幼英. 紅茶萎凋過程中多酚氧化酶生化特性的變化[J]. 茶葉科學簡報, 1990(4): 16-19.
[30] OZDEMIR F, TONTUL I, BALCI-TORUN F, et al. Effect of rolling ? methods and storage on volatile constituents of Turkish black tea[J]. ? Flavour and Fragrance Journal, 2017, 32 (5): 362-375.
[31] JIANG Y W, HUA J J, WANG B, et al. Effects of variety, season, and? ? region on theaflavins content of fermented chinese congou black? ? tea[J]. Journal of Food Quality, 2018: 1-9.
[32] DAS S, SAMANTA T, DATTA A K. Improving black tea quality? ? through optimization of withering conditions using artificial neural ? network and genetic algorithm[J]. Journal of Food Processing and ? Preservation, 2021, 45 (3): 1-9.
[33] HUA J J, XU Q, YUAN H B, et al. Effects of novel fermentation method on the biochemical components change and quality formation of Congou black tea[J]. Journal of Food Composition and Analysis, 2021, 96: 1-11.
[34] 吳本剛, 肖孟超, 劉美娟, 等. 催化式紅外殺青對綠茶熱風干燥的影響[J]. 食品科學, 2017, 38 (9): 126-132.
[35] 鄭文佳, 馮林, 沈強, 等. 綠茶疊殺后PPO和含水量變化及其對品質的影響[J]. 西南農業學報, 2011, 24 (5): 1714-1718.
[36] 譚俊峰. 超高壓對茶葉主要品質成分含量的影響[D]. 北京: 中國農業科學院, 2008.
[37] 王麗霞, 肖麗霞. 烏龍茶鮮葉萎凋過程中多酚氧化酶活性的變化[J]. 安徽農業科學, 2008 (23): 10191-10192.
[38] 王承莉, 張聰, 劉更生, 等. 高茶多糖烏龍茶的做青工藝研究[J]. 美食研究, 2015, 32 (4): 60-64.
[39] 陳倩蓮, 王芳, 莫楚紅, 等. 不同升溫方式下大紅袍做青中PPO和β-G活性變化對比[J]. 食品研究與開發, 2021, 42 (7): 28-32.
[40] 柴潔, 馬存強, 周斌星, 等. 普洱茶固態發酵過程中多酚氧化酶酶學活性研究[J]. 食品工業科技, 2013, 34 (21): 153-156, 161.
[41] 胡沛然. 六堡茶多酚氧化酶及其產生菌對茶葉品質的應用[D]. 南寧: 廣西大學, 2019.
[42] 付贏萱, 劉通訊. 多酚氧化酶對普洱茶渥堆發酵過程中品質變化的影響[J]. 現代食品科技, 2015, 31 (3): 197-201.
猜你喜歡
科技視界(2016年21期)2016-10-17 19:54:47
科技視界(2016年21期)2016-10-17 19:54:05
科技視界(2016年21期)2016-10-17 18:46:46
科技視界(2016年21期)2016-10-17 18:37:58
科技視界(2016年21期)2016-10-17 18:28:05
考試周刊(2016年76期)2016-10-09 08:45:44
大學教育(2016年9期)2016-10-09 08:28:55
科技視界(2016年20期)2016-09-29 14:22:00
科技視界(2016年20期)2016-09-29 12:03:12
科技視界(2016年20期)2016-09-29 11:47:01
