苜蓿抗旱性分子研究進(jìn)展
2021-09-14 04:37:12李倩江文波王玉祥張博龐永珍
生物技術(shù)通報(bào) 2021年8期
李倩 江文波 王玉祥 張博 龐永珍
(1. 中國農(nóng)業(yè)科學(xué)院北京畜牧獸醫(yī)研究所,北京 100193;2. 新疆農(nóng)業(yè)大學(xué)草業(yè)與環(huán)境科學(xué)學(xué)院,烏魯木齊 833400)
苜蓿屬(Medicago)植物屬豆科蝶形花亞科,包括83個(gè)種,目前研究較為廣泛的飼草主要包括模式植物蒺藜苜蓿(Medicago truncatula)、黃花苜蓿(Medicago falcata L.)和紫花苜蓿(Medicago sativa L.)[1]。其中紫花苜蓿是世界上種植面積最為廣泛且最具經(jīng)濟(jì)價(jià)值的豆科栽培牧草[2],全世界的紫花苜蓿種植面積約 3.22×107hm2,而我國苜蓿種植面積約為3.77×106hm2[3],主要分布在西北、華北、東北各省區(qū),南方各地也開始栽種[4]。黃花苜蓿在我國分布廣泛,大部分為野生資源,具有較好的抗性,適口性好,為牛、羊提供豐富的營養(yǎng),可與紫花苜蓿雜交形成抗性強(qiáng)、利用價(jià)值更高的雜花苜蓿,是苜蓿抗性育種的重要基因庫[5]。雜花苜蓿抗逆性強(qiáng),雜種優(yōu)勢(shì)顯著,表型多樣性豐富,具有良好的選育潛質(zhì),可為進(jìn)一步選育不同優(yōu)良特性的新品種提供適宜的育種材料[6];蒺藜苜蓿是一年生植物,其基因組小且自花授粉,是研究豆科牧草的模式植物[7]。盡管苜蓿品種能夠適應(yīng)水分有限的地區(qū),但水分虧缺仍然影響紫花苜蓿的產(chǎn)量。因此,通過提高苜蓿抗旱能力進(jìn)而提高草產(chǎn)量已成為紫花苜蓿研究的重要方向[8-9]。
干旱脅迫嚴(yán)重影響苜蓿的生長(zhǎng)、發(fā)育及產(chǎn)量。據(jù)估計(jì),干旱和半干旱地區(qū)約占全球總面積的30%[10]。如今,人們已充分認(rèn)識(shí)到干旱脅迫嚴(yán)重制約了苜蓿產(chǎn)業(yè)的發(fā)展,并通過分子生物學(xué)手段和技術(shù)對(duì)紫花苜蓿加以改良。因此,以常規(guī)育種為基礎(chǔ),借助于生物技術(shù)改良抗旱性狀,培育高產(chǎn)、高抗、優(yōu)質(zhì)的新品種,是一種更直接、有效的方式。本文將從分子水平上對(duì)4種苜蓿的抗旱性進(jìn)行分析,詳細(xì)總結(jié)目前苜蓿抗旱基因及抗旱分子育種方面相關(guān)的研究進(jìn)展,以期為苜蓿的抗旱品種改良提供參考。
1 分子學(xué)水平的研究進(jìn)展
1.1 組學(xué)研究進(jìn)展
隨著對(duì)苜蓿的研究不斷深入,苜蓿的抗性育種和品種改良已成為當(dāng)下研究的重點(diǎn)。但由于黃花苜蓿的基因組信息尚未公布,只能參考豆科模式植物蒺藜苜蓿和紫花苜蓿的基因組信息輔助研究黃花苜蓿的抗旱性。在紫花苜蓿(新疆大葉)的基因組已經(jīng)公布的情況下[11],結(jié)合轉(zhuǎn)錄組測(cè)序技術(shù),對(duì)苜蓿的抗逆基因挖掘和生物信息學(xué)分析具有重要的參考價(jià)值。
在紫花苜蓿基因組公布之前,有多個(gè)關(guān)于紫花苜蓿轉(zhuǎn)錄組測(cè)序的研究報(bào)道,為研究紫花苜蓿抗旱分子機(jī)理提供參考。通過轉(zhuǎn)錄組測(cè)序技術(shù)揭示了苜蓿內(nèi)源NO脅迫耐受的相關(guān)基因和途徑[12],Luo等[13]利用三代測(cè)序技術(shù)PacBio構(gòu)建了第一個(gè)紫花苜蓿根尖在連續(xù)甘露醇處理下的全長(zhǎng)轉(zhuǎn)錄組數(shù)據(jù)庫,共鑒定了8 016個(gè)受甘露醇調(diào)節(jié)的差異表達(dá)基因。還通過代謝分析發(fā)現(xiàn)滲透相關(guān)基因,篩選出來的基因可作為未來研究抗旱的關(guān)鍵候選基因。研究人員于2020年利用三代PacBio、Bionano及HIC技術(shù),結(jié)合最先進(jìn)組裝策略破譯了紫花苜蓿栽培種“中苜1號(hào)”基因組,并對(duì)世界范圍內(nèi)的紫花苜蓿核心種質(zhì)進(jìn)行重測(cè)序并獲得許多重要農(nóng)藝性狀關(guān)聯(lián)候選區(qū)域[14]。另一項(xiàng)研究通過GWAS與基因分型的測(cè)序鑒定了紫花苜蓿與抗旱系數(shù)和葉片相對(duì)含水量等抗旱性狀相關(guān)的位點(diǎn)[15]。Ray等[16]利用重測(cè)序技術(shù)檢測(cè)了在干旱脅迫下紫花苜蓿生物量相關(guān)的數(shù)量性狀位點(diǎn),Wang[17]等也證明GWAS結(jié)合GBS是分析紫花苜蓿農(nóng)藝性狀的一種有效方法,其結(jié)果對(duì)進(jìn)一步鑒定表型相關(guān)候選基因具有重要的參考價(jià)值。
為了篩選黃花苜蓿抗旱相關(guān)基因,研究人員通過構(gòu)建抑制性差減雜交文庫和轉(zhuǎn)錄組測(cè)序技術(shù),挖掘了植物激素代謝相關(guān)信號(hào)途徑中的關(guān)鍵基因,篩選了黃花苜蓿中干旱響應(yīng)的特異基因[18]和一個(gè)受脫水干旱脅迫誘導(dǎo)顯著上調(diào)表達(dá)的NAC轉(zhuǎn)錄因子MfNACsa[19],這些基因都可用于牧草和作物育種的候選基因。對(duì)干旱脅迫下黃花苜蓿轉(zhuǎn)錄組的sRNA測(cè)序,結(jié)果表明黃花苜蓿一方面通過調(diào)控膜蛋白來增加水分和離子的流動(dòng)以調(diào)節(jié)滲透勢(shì),另一方面通過促進(jìn)磷脂代謝,提高膜系統(tǒng)的穩(wěn)定性,從而調(diào)控干旱[20]。
隨著苜蓿基因組的公布以及測(cè)序技術(shù)的不斷更新,多組學(xué)相結(jié)合分析苜蓿抗旱機(jī)理成為目前的研究趨勢(shì)。通過轉(zhuǎn)錄組挖掘干旱逆境脅迫相關(guān)基因[21],發(fā)現(xiàn)了蒺藜苜蓿 R2R3-MYB 轉(zhuǎn)錄因子主要參與調(diào)控氧化還原反應(yīng)、細(xì)胞信號(hào)轉(zhuǎn)導(dǎo)以及轉(zhuǎn)錄調(diào)控等過程,從而抵御干旱等非生物脅迫[22]。Kang等[23]利用蒺藜苜蓿全基因組關(guān)聯(lián)分析發(fā)現(xiàn),醛脫氫酶家族蛋白與氣孔密度具有顯著相關(guān)性,說明該基因在干旱脅迫下嚴(yán)重影響氣孔密度。為探究 miRNAs 在植物響應(yīng)干旱脅迫中的作用,Wang等[24]通過Illumina高通量測(cè)序在蒺藜苜蓿中鑒定了miRNA家族的新成員發(fā)現(xiàn),這些miRNA均可響應(yīng)干旱脅迫。
1.2 功能基因研究
對(duì)苜蓿抗旱功能基因的研究主要集中在轉(zhuǎn)錄因子和基因表達(dá)調(diào)控方面,通過克隆抗旱相關(guān)基因,可闡明紫花苜蓿抗旱調(diào)控中的分子機(jī)制,并有望應(yīng)用于分子改良中[25]。目前,一部分轉(zhuǎn)錄因子也已經(jīng)被發(fā)現(xiàn)參與了苜蓿應(yīng)答干旱脅迫(表1)。
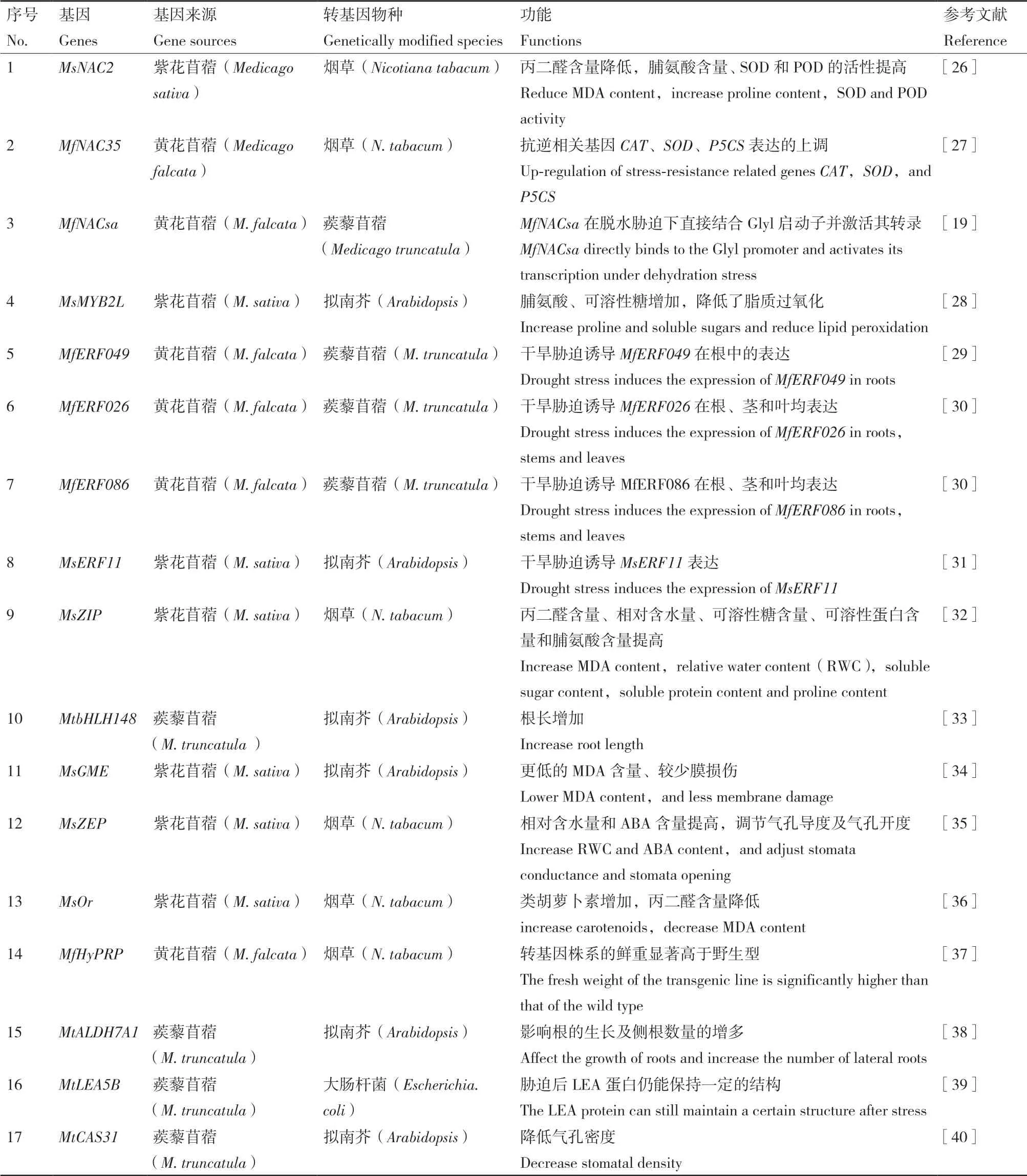
表1 苜蓿來源的抗旱相關(guān)基因及其功能研究Table 1 Research on drought resistance related genes and their functions from Medicago
NAC轉(zhuǎn)錄因子是植物特有的一類轉(zhuǎn)錄因子,在植物生長(zhǎng)發(fā)育、響應(yīng)逆境脅迫途徑中起重要作用。目前,紫花苜蓿中MsNAC2[26]、黃花苜蓿中MfNAC35[27]和MfNACsa[19]等基因都被成功克隆,基因表達(dá)模式均已明確,實(shí)驗(yàn)證明這些基因可能參與調(diào)控抗旱逆境脅迫的響應(yīng)。MYB型轉(zhuǎn)錄因子參與植物對(duì)多種脅迫因子的反應(yīng)。然而,在苜蓿中,只有少數(shù)幾個(gè)MYB基因與脅迫反應(yīng)相關(guān)。將紫花苜蓿的MsMYB2L基因在擬南芥中過表達(dá)時(shí),干旱脅迫處理后發(fā)現(xiàn),該基因的過量表達(dá)促進(jìn)了脯氨酸、可溶性糖等滲透調(diào)節(jié)物質(zhì)的合成,降低了脂質(zhì)過氧化,從而提高了轉(zhuǎn)基因擬南芥的抗旱性[28]。ERF轉(zhuǎn)錄因子是植物特有的一類轉(zhuǎn)錄因子,廣泛參與植物多種逆境脅迫。苜蓿中MfERF049[29]、MfERF026、MfERF086[30]和MsERF11[31]被成功克隆,不同的非生物脅迫都可誘導(dǎo)這些基因的表達(dá),表明這些基因可能參與了多種逆境脅迫的調(diào)控過程。Li等[32]克隆了MsZIP基因,過量表達(dá)該基因的煙草對(duì)滲透壓脅迫的耐受性增強(qiáng)。王菊萍等[33]從蒺藜苜蓿幼苗中克隆了MtbHLH148 基因,發(fā)現(xiàn)該基因受自然脫水脅迫處理后表達(dá)量上調(diào)。目前,對(duì)于苜蓿中抗旱相關(guān)轉(zhuǎn)錄因子的研究,也只是將候選基因進(jìn)行克隆奠定,并且證明這些基因受不同非生物脅迫的誘導(dǎo),部分基因被轉(zhuǎn)化進(jìn)模式植物煙草或者擬南芥中進(jìn)行功能驗(yàn)證,但均未轉(zhuǎn)入本體植物苜蓿中進(jìn)行功能驗(yàn)證。
除了轉(zhuǎn)錄因子,苜蓿來源的其它類型的基因也參與干旱等多種逆境脅迫反應(yīng)(表2)。研究表明,紫花苜蓿來源的GDP-甘露糖3,5-表異構(gòu)酶MsGME基因在擬南芥中異源表達(dá)時(shí),可通過降低MDA含量、較少膜損傷,以及增加抗壞血酸的積累來增強(qiáng)其抗旱能力[34]。紫花苜蓿來源的玉米黃質(zhì)環(huán)氧化 酶(MsZEP)[35]和Orange(MsOr)基 因[36]在煙草中表達(dá)時(shí),可通過分別提高相對(duì)含水量、ABA 含量、類胡蘿卜素含量,調(diào)節(jié)氣孔導(dǎo)度及氣孔開度,從而增強(qiáng)抗旱性。在煙草中過量表達(dá)黃花苜蓿的MfHyPRP[37]基因時(shí),增加了肌醇、甜菜堿和棉籽糖的合成量,在干旱脅迫處理下,轉(zhuǎn)基因煙草相對(duì)于未轉(zhuǎn)基因?qū)φ站哂懈叩墓夂闲屎透偷碾x子滲漏,對(duì)干旱的耐受性增強(qiáng)。在蒺藜苜蓿中對(duì)MtALDH7A1[38]和MtLEA5B[39]基因的克隆和深入研究證明這些基因?qū)Ω珊瞪车目鼓骓憫?yīng)具有重要作用。另外的一項(xiàng)研究表明,在擬南芥中過表達(dá)苜蓿來源的低溫馴化特異蛋白基因MtCAS31時(shí),顯著降低了轉(zhuǎn)基因擬南芥的氣孔密度,大大提高了轉(zhuǎn)基因擬南芥的耐旱性[40]。
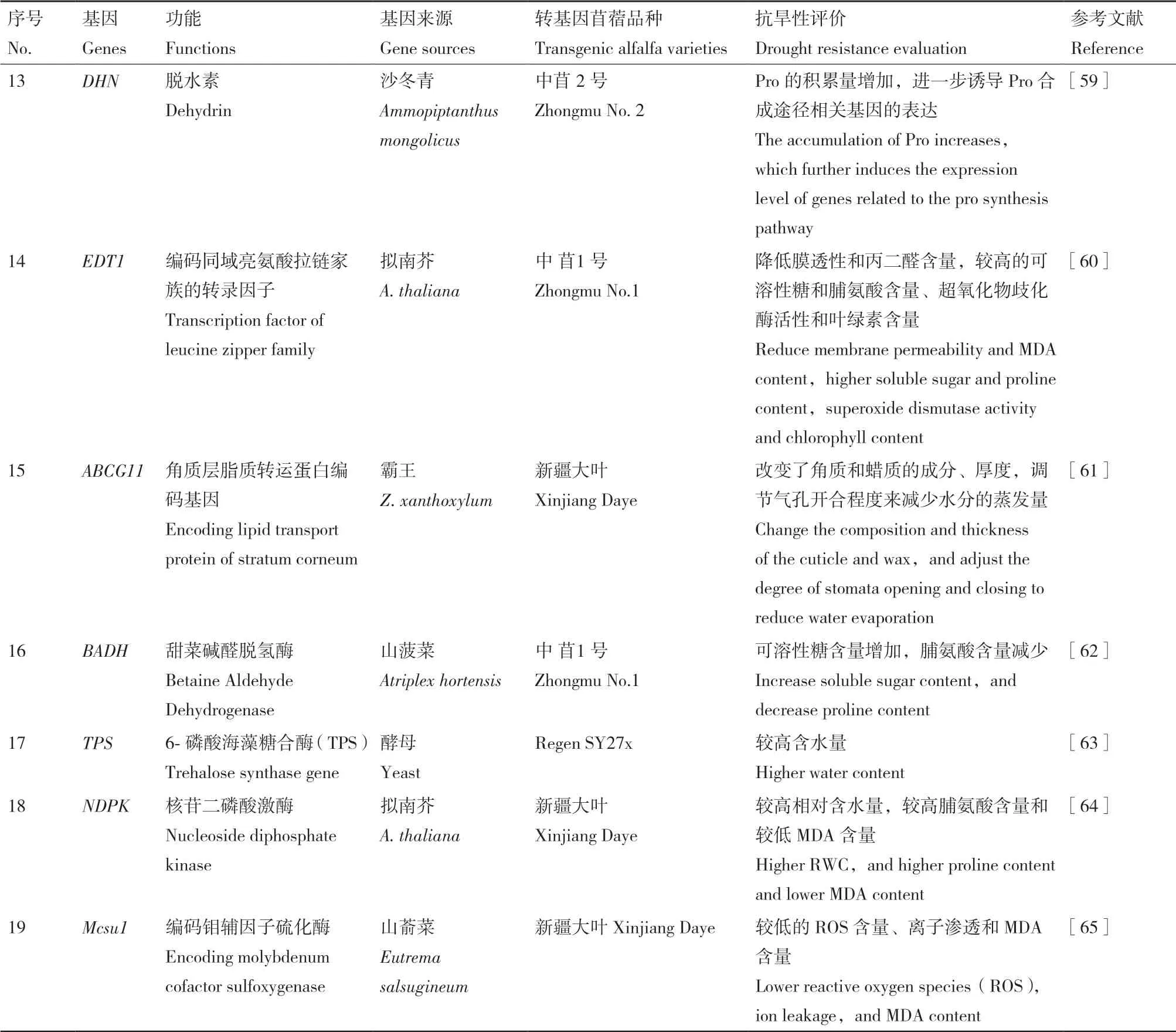
表2 續(xù)表 Continued
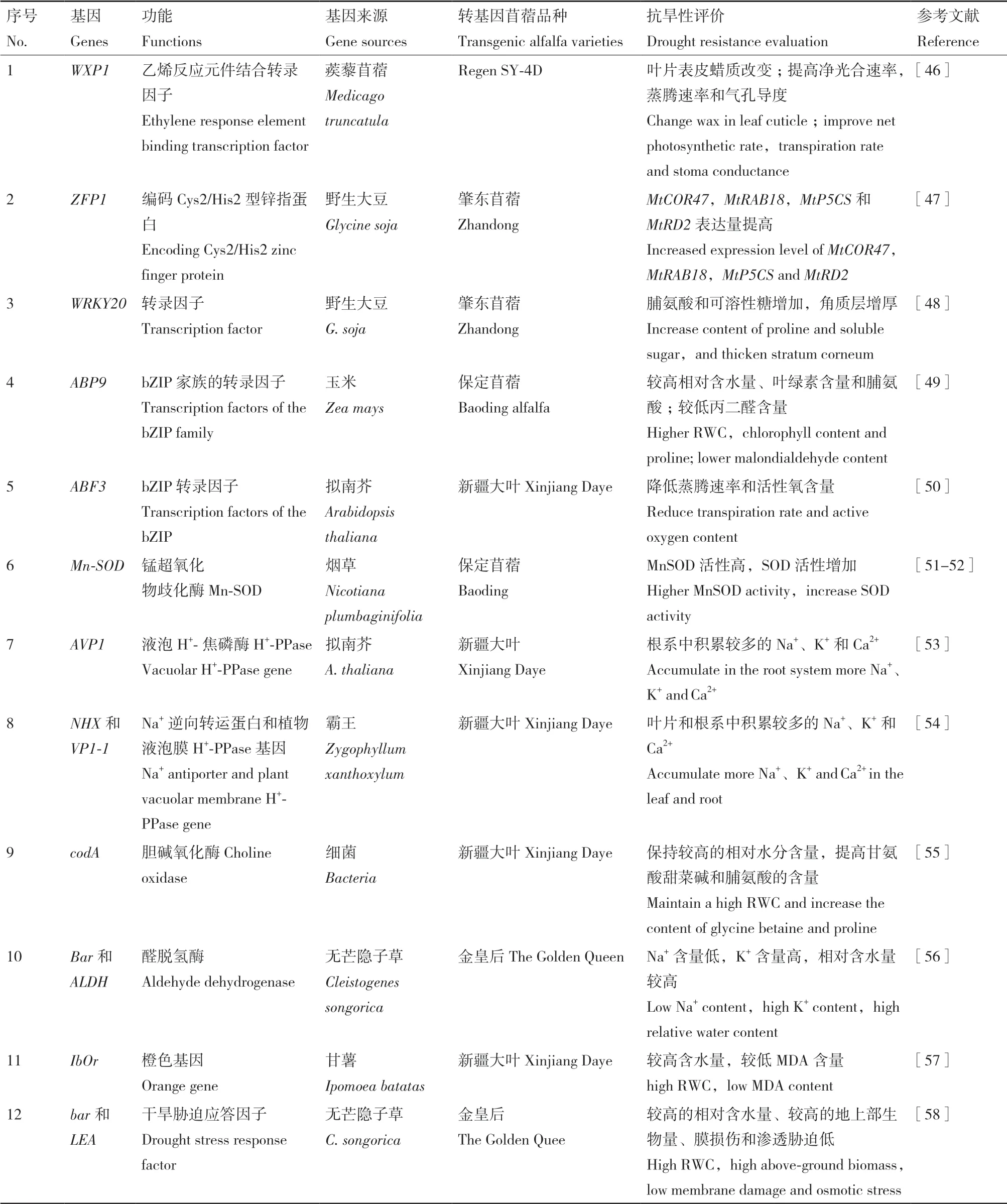
表2 用于提高紫花苜蓿抗旱性的基因及其作用Table 2 Genes used to improve drought resistance of alfalfa and their functions
以上研究大多通過同源克隆的方式研究苜蓿的關(guān)鍵轉(zhuǎn)錄因子或酶基因,并進(jìn)行序列分析,闡明基因在不同組織中的表達(dá)模式;相對(duì)于對(duì)模式植物抗旱機(jī)制的研究而言,目前對(duì)苜蓿抗旱相關(guān)基因的功能驗(yàn)證和應(yīng)用較為粗淺,大多數(shù)研究均是將相關(guān)基因通過生物技術(shù)轉(zhuǎn)入其它模式植物或者紫花苜蓿/蒺藜苜蓿中,進(jìn)行初步的功能分析,更具體的抗旱調(diào)控分子機(jī)理尚不清楚,還需要進(jìn)一步的研究和探索。
2 利用基因工程進(jìn)行分子育種
目前我國苜蓿育種工作中可供利用的優(yōu)異種質(zhì)資源嚴(yán)重缺乏,迫切需要發(fā)掘和創(chuàng)造新種質(zhì)、新材料、新基因[41]。目前只有抗農(nóng)達(dá)除草劑和低木質(zhì)素含量的苜蓿新品種已經(jīng)投放市場(chǎng)[42-43],但在我國審定登記的苜蓿品種中,仍無單純運(yùn)用生物技術(shù)培育成的紫花苜蓿品種。為了加快育種進(jìn)程,克服傳統(tǒng)育種缺陷,全世界育種學(xué)家們熱衷于基因工程技術(shù)輔助的育種方法[44]。為了減輕苜蓿生產(chǎn)過程中的產(chǎn)量和品質(zhì)損失,大力發(fā)展適應(yīng)各種非生物脅迫的苜蓿新品種。自1986年苜蓿轉(zhuǎn)基因技術(shù)首次取得成功后[45],該技術(shù)為苜蓿改良提供了一種有效的技術(shù)手段,目前很多抗旱相關(guān)基因均用于苜蓿抗旱的分子改良中,這些基因主要分為兩類:轉(zhuǎn)錄因子和其它類型的基因。
將外源基因轉(zhuǎn)入紫花苜蓿中,轉(zhuǎn)基因植株通過減少膜損傷和滲透脅迫來提高耐旱性,這主要?dú)w因于增強(qiáng)葉片表皮蠟質(zhì)、增厚角質(zhì)層、減少水分流失和增強(qiáng)光合作用,進(jìn)而提高脯氨酸含量、可溶性糖的含量、葉綠素含量、相對(duì)含水量和酶活性、降低丙二醛含量、蒸騰速率和活性氧含量。盡管不同基因在抗旱中分子機(jī)制可能有所不同,但它們都能通過滲透調(diào)節(jié)、離子調(diào)節(jié)和改變酶活性,從而提高轉(zhuǎn)基因苜蓿的抗旱性。
3 展望
植物抗旱是一個(gè)非常復(fù)雜的機(jī)制,是受多基因控制的數(shù)量性狀,是耐旱相關(guān)代謝途徑以及其它多種代謝途徑協(xié)同作用的結(jié)果[66],苜蓿的耐旱分子機(jī)理仍不清楚。目前有關(guān)苜蓿抗旱性的分子研究進(jìn)展,主要集中在通過組學(xué)研究篩選抗旱相關(guān)基因,大多數(shù)研究均是將相關(guān)基因通過生物技術(shù)轉(zhuǎn)入其它模式植物或者紫花苜蓿/蒺藜苜蓿中,進(jìn)行初步的功能分析,更具體的抗旱調(diào)控分子機(jī)理尚不清楚,還需要進(jìn)一步的研究和探索。關(guān)于苜蓿分子育種研究較為遲緩,也無單純運(yùn)用生物技術(shù)培育成的紫花苜蓿品種。目前熱門的基因編輯技術(shù)可能對(duì)紫花苜蓿進(jìn)行定向遺傳改良,需要全面研究并解析紫花苜蓿耐旱相關(guān)代謝途徑、耐旱相關(guān)基因及其表達(dá)模式。而通過傳統(tǒng)育種也未培育出抗旱性強(qiáng)的紫花苜蓿材料和品種,所以通過分子生物學(xué)技術(shù)手段提高紫花苜蓿的抗旱性是進(jìn)行紫花苜蓿分子育種的有效手段,但仍然存在一些問題。首先,缺乏系統(tǒng)完整的抗旱性鑒定評(píng)價(jià)標(biāo)準(zhǔn);其次,轉(zhuǎn)基因株系一般是基于人工模擬的環(huán)境-實(shí)驗(yàn)室或溫室中進(jìn)行的干旱脅迫,大多缺少相應(yīng)的品種區(qū)域特性試驗(yàn),很少投入到實(shí)際生產(chǎn)過程中。而區(qū)域試驗(yàn)是評(píng)價(jià)新品種適應(yīng)性的最直接可靠的辦法,需要設(shè)立和增加轉(zhuǎn)基因紫花苜蓿區(qū)域試驗(yàn)站(點(diǎn))的數(shù)量,從而使轉(zhuǎn)基因的新品系得到大田驗(yàn)證并得以推廣示范用于生產(chǎn)應(yīng)用[67]。因此,進(jìn)行紫花苜蓿育種是一項(xiàng)長(zhǎng)期的工作。
苜蓿轉(zhuǎn)基因的安全性也是一個(gè)備受關(guān)注的熱點(diǎn)話題。首先,紫花苜蓿作為重要的豆科飼草主要應(yīng)用于牲畜中,而不是直接應(yīng)用于人類自身,相對(duì)而言也許較為安全。其次,相對(duì)于牧草品質(zhì)性狀的改良而言,將抗旱等抗逆性狀基因轉(zhuǎn)入紫花苜蓿中并不改變紫花苜蓿的品質(zhì)性狀,對(duì)動(dòng)物和人類的食品安全的影響可能更小。最后,黃花苜蓿是目前很多紫花苜蓿育成品種的親本材料,黃花苜蓿本身的抗旱能力很強(qiáng),如果將黃花苜蓿的抗旱基因轉(zhuǎn)入紫花苜蓿,在保證產(chǎn)量的前提下提高抗性,也許遠(yuǎn)比轉(zhuǎn)入其它植物來源的外源基因更加安全可靠,這可能是進(jìn)行紫花苜蓿抗旱性改良的最有效的策略,也是當(dāng)前苜蓿抗旱育種的研究目標(biāo)。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
