放牧對高寒草地微尺度斑塊及其構件特征的影響
2021-09-25 03:09:50林麗曹廣民樊博柯潯李茜蘭玉婷朋措吉戴黎聰李以康周春麗
草原與草坪 2021年4期
林麗,曹廣民,樊博,柯潯,李茜,蘭玉婷,朋措吉,戴黎聰,李以康,周春麗
(1.中國科學院西北高原生物研究所,青海 西寧 810008;2.中國科學院大學,北京 100049)
高寒草地是青藏高原的主體,占青藏高原可利用草地面積的50.9%[1],對青藏高原生態系統服務價值貢獻率達89.6%[2-3],具有生態、生產和生活的“三生”服務功能[4]。高寒草甸和高寒草原是高寒草地生態系統的主體,廣布于青藏高原東部,是高原地帶性與山地垂直帶譜的主要植被類型[5],對支撐高原畜牧業發展、維系農牧民生活、維護邊疆穩定、保護生物多樣性、涵養水源和維持碳素平衡具有不可替代的作用[4]。高寒草地所處區域氣候惡劣、生存環境嚴酷、生態系統結構簡單、自我更新能力弱,加之固有的“惰性”特征成為退化生態系統恢復的世界性難題[4-6]。近年來,受到氣候、經濟、政策、文化等多重因素的影響[7],高寒草地植被發生不同程度的演化,其演化的主流方向為退化,尤其在圍欄建設和家庭牧場自主經營模式下,牧區草地景觀破碎化日益加劇,出現了草地多演替穩態并存的景觀特征[8]。植被斑塊化分布是放牧生態系統的固有特性,是系統結構與功能維持的表現形式[9-10],斑塊鑲嵌不僅是景觀尺度草地的基本結構,也是微尺度草地資源的賦存形式和草地演化的歷史承載體。草地生態系統微尺度斑塊(簡稱微斑塊)的研究主要圍繞斑塊屬性特征同系統之間的關系、斑塊構件特征(斑塊構件在生態過程中的功能及作用)以及斑塊動態特征(即斑塊屬性特征及構件特征在時間尺度上的變化過程)開展[11]。從生態學的角度出發對生態系統斑塊的研究逐漸從宏觀向微觀尺度發展,在此過程中研究者更關注微斑塊的鑲嵌結構、微斑塊構件特征及其動態變化所引發的連鎖反應對環境總體特征的影響。從微斑塊尺度上進行試驗研究的優點主要包括:1)測量方法精細;2)樣本分級尺度自由,可以根據研究需要在給定強度上進行取樣分級設定,反映觀察對象局域和整體特征;3)試驗處理相對易于操作;4)易于在樣地內進行重復試驗或觀測[12]。對微斑塊及其構件特征的研究有助于較為全面的認識高寒草地生態系統構件屬性,剖析生態系統組織形式、資源配置過程及其應對干擾的調節機制,對探討高寒草地生態系統演化過程、系統穩定性維持機制具有重要的理論指導意義,對受損高寒草地恢復研究提供技術支撐。
1 微斑塊是放牧生態系統構件特征的賦存和組織形式
對放牧型生態系統微斑塊屬性特征的研究首先應該確定微斑塊類型。目前對微斑塊類型的劃分還沒有十分明確的標準,一般研究人員可以按照研究目的,結合植物特征和地表特征定義和劃分[12-14],常見的劃分標準包括4種:1)以生活型劃分,如直根型植物斑塊、根莖疏叢型植物斑塊、根莖密叢型植物斑塊、密叢型植物斑塊;2)以植物群落特征劃分,如高草型(如早熟禾斑塊、針茅斑塊、羊茅型斑塊)、低草型(如矮嵩草斑塊、小嵩草斑塊)、伏地型(如地衣斑塊、苔蘚斑塊、藻類斑塊);3)以經濟類群劃分,如禾本科斑塊、莎草科斑塊、雜類草斑塊、豆科斑塊;4)以牧用價值劃分,如可食性牧草斑塊、不可食性牧草斑塊等[15-29]。
當代的高寒草地生態系統屬于人類干擾條件下的特殊自然生態系統,具有開放性、連續性、自組織性等自然生態系統的固有屬性特征,而這些特性主要是以微斑塊為賦存載體和組織形式。
首先,高寒草地生態系統具有一定的等級特征,可以將其分解成若干有序的離散等級層次(discrete hierarchical level),且不同等級單元均具有一定的結構、功能和動態變化規律[12-14],微斑塊則可以作為基本研究載體,根據研究需要以不同時空尺度反映生態系統等級結構和組織形式。高寒草地生態系統斑塊研究單元大致可以劃分成5個時空尺度水平,包括巨大尺度(megascale)、大尺度(macroscale)、中尺度(mesoscale)、小尺度(microscale)[12,15]和微尺度(miniscale),而微斑塊的研究多集中在微尺度(空間范圍小于1 m2,時間范圍小于1年)和小尺度(空間范圍1~1×106m2,時間范圍1~500年)范圍內。
微(小)尺度斑塊研究可以根據研究需要選擇不同研究方法。1)數學模型法。此方法以微(小)尺度斑塊屬性特征為實體,通過尺度維繹轉換探討斑塊彼此之間屬性特征及其變化過程。其缺點是在尺度的轉換過程中基礎數據的代表性決定了模型的適用性[16-18]。2)數量統計法。應用經典數量生態學統計方法對微斑塊屬性進行統計分析(如概率分布統計、排序等)以探討生態系統斑塊特征的分異和變化[19],較適用于不同尺度斑塊屬性特征的空間配置分析。3)遙感技術。應用遙感照片對斑塊鑲嵌體進行分割,通過野外實地抽樣或圖件信息獲得斑塊的屬性信息,利用適宜的模型完成斑塊屬性的空間格局圖像,此方法像元的精確度決定了其研究結果的精度,其缺點為屬性數據的獲取與遙感影像資料需要高度匹配,地物矯正數據的獲取及高分辨率影像資料的獲取需要消耗大量的人力物力[20]。4)地理信息系統(GIS)。地理信息系統中的柵格數據可以實現對微(小)尺度時域數據和屬性數據的格局特征分析,其缺點同遙感技術相似,高分辨率屬性數據比較適用于微(小)尺度斑塊特征研究,但如果推廣到較大尺度,則可能受到諸多因素的限制[21-22]。
第二,高寒草地生態系統不同微斑塊屬性特征在空間上相互隔離,功能上彼此聯系,具有一定的自組織能力,即在一定程度上可以實現生態系統局部易變而整體穩定的特征。高寒草地生態系統植物群落具有群聚效應,加之所處區域地形地貌復雜、氣候多變,容易形成不同分布格局的植物鑲嵌體斑塊。根據島嶼生物地理學理論(island biogeography theory),可以將植物鑲嵌體微斑塊視為島嶼,其周圍基質視為大陸,植物鑲嵌體斑塊物種豐富度的穩定性取決于物種周轉率、斑塊面積和植物鑲嵌體斑塊彼此之間的距離。對于同一植物鑲嵌體斑塊,當物種周轉率為零時,物種的數目相對穩定,達到動態平衡[23]。放牧是高寒草地生態系統的固有干擾因素。放牧家畜對草地植物鑲嵌體斑塊的作用強度具有非勻質性,極易導致生境破碎化,當生境破碎后,空間上容易形成具有一定距離的生境斑塊(habitat patch)或小的局部種群鑲嵌結構體斑塊(local population)[24],這種種群鑲嵌結體斑塊在一定程度上保有了不同退化演替階段的植物群落特征,成為生態系統草地資源的保存形式之一;同時草地生態系統種群鑲嵌結構體斑塊之間的關聯性、滲透性客觀上促進了高寒草地生態系統微斑塊特征的易變性,區域整體特征的穩定性,是高寒草地系統穩定性維持的原因之一。
第三,高寒草地生態系統微斑塊具有一定的連續性,構件屬性在一定閾值范圍內可以彼此滲透和轉化。高寒草地微斑塊系統是一個開放系統,對外界干擾具有一定程度的自我調節能力,且自我調節能力一般表現在不同子系統對外界干擾反饋的非線性疊加效應上,這種非線性疊加效應容易造成高寒草地生態系統對干擾反應的滯后性和生態系統在平衡點周圍的震蕩(波動)性。此外,在放牧干擾條件下,當微斑塊之間的基質媒介物密度達到臨界值時,可以實現微斑塊屬性特征的滲透和轉化,從而實現降低生境破碎化對種群動態影響的目的[25]。目前高寒草地生態系統微斑塊屬性特征研究多集中在生態系統構件特征與干擾因子之間的耦合關系及生態系統自組織能力維持的閾值區間上,而對系統自組織能力維持的時效性研究較為薄弱。
2 放牧對生態系統微斑塊構件特征的影響
2.1 土壤構件特征
天然放牧草地的微斑塊特性與其成因和過程表現在多重時間尺度上[26-27]。地形、地貌以及由此造成的水熱資源的差異決定了微斑塊土壤養分的基本特征,環境變化或放牧干擾導致草地生態系統景觀破碎化,形成不同形態及大小的植物-土體微斑塊復合體[28],引起草地土壤養分的異質化[9,29]。因此,土壤構建特征是草地微斑塊的重要構件特征。與生命活動密切相關的土壤養分(土壤全碳、全氮、速效氮、速效磷)空間分布同植物群落的微斑塊特征密切相關[30]。微斑塊土壤資源的有效性及轉移過程強烈地受制于微斑塊植物群落特征,微斑塊土壤養分(如可溶性有機碳、無機氮等)的積累同植物群落對養分的攔截具有明顯關聯性[31-32],因此,微斑塊土壤養分含量(如土壤交換性離子含量)、養分供給能力(如土壤水分滲透速率)、土壤微生物活性(提高土壤呼吸速率)均受制于植物群落特征。放牧干擾強度不同亦會改變土壤微斑塊對養分的富集能力、效率及在系統中的分配比例[33-35]。土壤理化特性的變化可以反作用于微斑塊植物群落的生長、組成結構的改變、多樣性的維持、群落的演替及地表特征的塑造[36-38]。
2.2 植物構件特征
2.2.1 植物群落數量特征 放牧對高寒草地不同微斑塊植物群落的干擾強度可以影響整個草地生態系統的植物群落特征[39]。隨著放牧強度的增加,植被微斑塊的組成、蓋度、高度和地上生物量等性狀總體呈現下降趨勢,微斑塊群落內植物種類組成趨于簡單化,放牧敏感物種消失,不可食性植物、先鋒種及R對策植物所在斑塊比例和面積顯著增加。但不同類型的斑塊在變化時段、幅度和格局上存在較大差異。微斑塊植物群落的多樣性指數、均勻度指數與退化程度呈正相關關系,而優勢度表現為負相關關系。在外界干擾下破碎化程度較低的草地易于形成較大的植被斑塊,邊緣效應的影響會促使物種多樣性增加;反之,在破碎化程度較高的草地,物種多樣性會急劇下降[40]。天然草地一定程度的植物群落微斑塊破碎度伴隨著植被微斑塊邊緣數量的增加而增加[41-44],因此,隨放牧強度增高,植物群落多樣性特征出現降低—增加—降低的趨勢,而微斑塊格局表現為勻質性—非勻質性—勻質性的趨勢[29-30]。
2.2.2 植物根系分泌物特征 根系分泌物是保持根際微生態系統活力的關鍵,也是根際微生態系統中物質遷移和調節的內驅力[45]。研究植物根系分泌物,對人類認識植物生長發育規律,揭示植物與環境互作關系,調控和評價植物種群及群落的生長狀態、明晰植被群落演化機理,提高草地健康管理能力具有重要意義[46]。廣義的根系分泌物主要包括滲出物、分泌物、粘膠質和裂解物質[47];狹義的根系分泌物僅包括植物通過溢泌作用進入土壤的可溶性有機物。
根系分泌物對放牧行為的響應可以在一定程度上反映植物體或微斑塊植物群落應對干擾的生理生化過程,主要表現在:1)當草地處于適度放牧條件下,微斑塊植物群落健康指數高,根系新陳代謝旺盛,根細胞的膜透性增強,一方面可以促進根系向環境中釋放更多代謝產物(如氨基酸、糖等),吸引有益土壤微生物聚集,改善根際營養環境[49];另一方面可以增加土壤礦物質養分的溶解度,促進根系對營養元素的吸收;2)過度放牧可以引起根系釋放根系分泌物用以干擾土壤微生物群體感應信號分子(quorum sensing signal)信息的傳導,控制土壤微生物的種群密度[50]、誘發土壤微生物生物膜(biofilm)的形成和有毒物質基因的表達、吸引有害生物天敵,控制有害病原體[51],優化植物生長環境[50],進而影響微斑塊土壤微生物的組分、活性、代謝產物及生態系統整體物質能量代謝速率與健康[52-53];3)不合理的放牧管理,可以引發根系分泌物代謝產物組分,增加化感物質和自毒素分泌,進而控制植物群落種群結構和密度,實現系統資源最大限度的可持續利用[54]。
2.3 土壤微生物構件特征
土壤微生物是土壤中最活躍的組分之一,它廣泛地參與成土過程,是土壤中物質和能量積累、循環、轉化等過程中的重要媒介和載體[55-57],是生物多樣性構建及系統穩定性維持的核心[58],是土壤肥力形成的基礎[59],是生態系統應對干擾的啟動因子[60],對評估土壤健康狀況和土體生物因子活力具有重要意義[61]。
青藏高原地域廣闊,生境復雜,氣候獨特,土壤微生物種類豐富[62],且土壤微生物組成、結構和分布特征同植物群落和土壤關系更為密切[63-64]。放牧對生態系統微斑塊土壤微生物組成、結構和分布特征的影響主要表現為改變土壤可利用養分、土壤微環境以及寄主的健康狀況。首先,放牧可以直接改變土壤養分濃度和土壤酶活性,進而影響土壤微生物組成和結構。如過度放牧可以顯著增加高寒小嵩草微斑塊真菌多樣性,降低土壤微生物總生物量[65-66]、土壤微生物活性[67]、生態系統物質能量的循環速率和系統的恢復性[65-66]。其次,放牧可以改變土壤養分配比,進而改變微生物的組成、結構和活性。根據最小限制因子定律,環境中的限制因子可以同時影響微斑塊土壤微生物活性和數量,進而對土壤微生物的組成、結構和分布產生影響[68-69]。第三,放牧可以改變微斑塊土壤微生物作用底物的品質,進而改變土壤微生物組成、結構和分布特征[70]。第四,放牧可以改變植物群落特征,直接改變微斑塊植物根系分泌物,間接改變土壤微生物特征[71-74]。第五,放牧強度和利用方式的改變可以直接影響生態系統微斑塊土壤微生物組成結構特征。由于土壤微生物群落的世代交替速度快于植物,因此土壤微生物類群對外界干擾更為敏感,同時土壤微生物對逆境環境的調節能力有限,在不同生態系統、不同演替過程會出現不同反應結果。如對高寒小嵩草進行短期禁牧對相同微斑塊細菌多樣性無顯著影響,而長期禁牧(如高于7年)則將顯著增加細菌生物量[75-76]。第六,土壤微生物自身對放牧干擾的敏感性決定了其在微斑塊中的組成和結構。如不同真菌對放牧干擾強度的調節能力不同,叢枝菌根真菌(AMF)與植物具有互利共生的作用,其多樣性與植物(根系生物量、地上部分的磷含量)數量特征、土壤養分儲量和結構(碳、氮、磷含量,化學計量等)密切相關[77],地上生物量的移除(采食、刈割等)對微斑塊植物群落生長造成威脅時,AMF的多樣性也將隨著微斑塊多樣性的降低而降低[74,78-79]。
此外,土壤微生物同微斑塊之間存在著相互“營養供應”關系,土壤微生物量的增加可以直接增加其所在微斑塊土壤有機質、有效磷等養分的含量和儲量,間接增加速效氮等養分含量,改善土壤根土比、土壤容重等理化性質,改善土壤環境,維持微斑塊土壤微生物系統多樣性,防治土源性病蟲害威脅,促進植物健康生長,維持一定程度的植物群落生產能力[80-82],且上述調控結果可以對土壤微生物量的形成產生正反饋效應[83](圖1)。
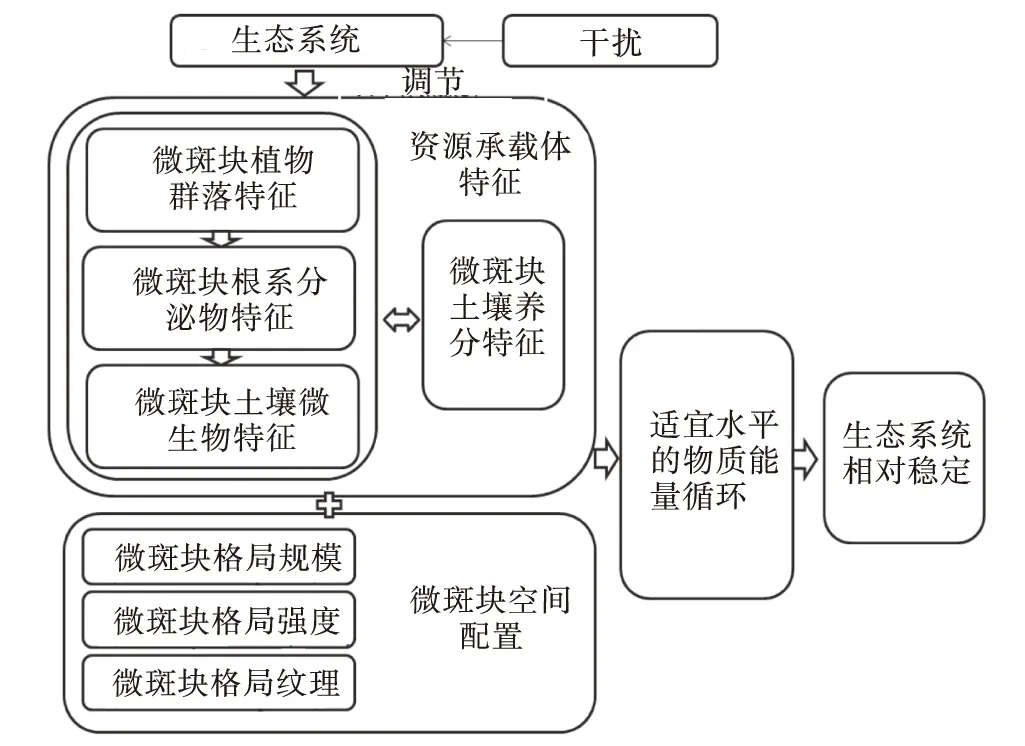
圖1 生態系統微斑塊同系統穩定性關系Fig.1 The relationship between the mini-patch and the ecosystem stabilization
3 放牧對生態系統微斑塊動態特征的影響
影響微斑塊配置格局、空間異質性及演化行為的主要驅動因子包括:1)非生物因子(氣候、土壤、地形地貌等)。微斑塊的資源異質性是決定微斑塊空間分布格局動態的首要因子[84-93],天然草地土壤養分和水分等分布的異質性是導致植被斑塊狀分布的先決條件[94]。如在干旱半干旱條件下,水分是草地生長的限制性因子,當天然草地處于長期低降水量時,微斑塊外圍生長的植被可以起到攔截地表徑流的作用,從而使其內層植物由于缺水而死亡,進而造成草地環狀植被微斑塊的形成[95]。2)生物因子。植物的生物學特性是微斑塊格局動態變化的決定性因素[96-97]。草地植被斑塊的形成受植物繁殖方式的影響[97]。3)管理因子。人類活動對草地植被微斑塊分布格局的控制主要體現在草地的放牧管理上(禁牧、放牧強度等)[98]。不同植物對放牧強度的響應具有差異,高強度放牧可限制天然草地不耐牧植物斑塊的生長發育甚至導致其死亡而變為裸地,從而形成由草地植被和土壤裸地鑲嵌分布的微斑塊,最終形成耐牧程度不同微斑塊系統復合體[99]。
微斑塊的形成與草地演化過程有著相同的驅動力,這種驅動力對草地不同微斑塊的作用強度不同,導致微尺度斑塊分布格局的改變,進而影響生態系統整體特征的改變[29]。放牧對草地植物群落微斑塊格局分布驅動力主要包括3個方面:1)家畜的選擇性采食是影響放牧生態系統植物群落微斑塊格局動態變化的重要因子[100-101]。天然草地植物群落的營養價值和能值是影響有蹄類家畜是否優先采食的重要衡量標準,能為家畜提供豐富能量和營養的植物群落斑塊(如禾本科植物和莎草科植物斑塊)被采食的頻度和強度顯著高于其他斑塊[100-101];喜食性牧草的分布特性是影響家畜采食的重要因子[102-103]。具有相同營養價值和能值的植物群落斑塊,集群分布的植物群落斑塊被家畜采食的頻度和強度高于隨機分布和均勻分布的植物群落斑塊,宏觀尺度表現為根莖密叢型、密叢型植物較根莖型植物群落斑塊被采食的幾率高[104-105]。此外,植物斑塊可食性牧草的多樣性決定該微斑塊是否能被家畜優先采食,因為多種植物比一種植物更能滿足家畜對營養需求和對毒素的排解[106],以上多種原因形成高寒草地生態系統微斑塊動態變化的原動力。2)放牧家畜排泄物通過覆蓋草地對植物起到窒息和灼燒的作用,導致天然草地中出現小裸斑;排泄物污染及異味,造成家畜對糞尿斑及其周圍區域的植被棄食,從而放大了家畜對排泄物斑塊棄食的空間尺度,客觀上造成不同微斑塊的動態轉移和滲透。3)糞尿斑的養分輸入造成斑塊土壤養分的沃島效應,成為導致放牧生態系統植被斑塊形成和分布的另一重要原因。非生物因子和人為因子可以在多種尺度上對微斑塊格局形成和動態變化產生作用,而生物因子(如捕食、競爭、植物-土壤相互作用等)主要在微(小)尺度斑塊格局的形成和變化中起主要作用[107-108]。
4 展望
土壤和植物是生態系統微斑塊物質能量運轉的載體和基礎,土壤養分、植物群落、植物代謝產物及其組合方式可以從不同角度描述微斑塊物質、能量代謝行為及變化[109-110],因此被廣泛的應用于生態系統穩定性的評價體系中。
理論認為具有高抵抗力的生態系統,受到干擾后不易偏離穩態結構,而一旦原有穩態結構被破壞,其恢復亦困難[111],其原因是,生態系統對其內部構件產生不同程度自適應調節作用,以達到干擾狀態下的最優組合[112],但這種調節能力有限,超過閾值,系統就很難再恢復為原狀態。生態系統自適應調控就是系統為達到最優狀態而進行的物質能量的調節和優化,是系統穩定性維持的基礎[113]。高寒草地生態系統是青藏高原可利用草地的主體,受到自然生態系統空間異質性、干擾源復雜性、生態系統構件耦合性、生態系統對干擾因子反應的異步性等因素的影響,傳統的隨機采樣方法獲得的生態系統構件特征對高空間異質性系統的表征能力有限,研究結果彼此之間差異大,對生態系統演化本質的描述顯得力不從心。因此,如何在高空間異質性條件下研究不同干擾源對高寒草地生態系統構件特征的影響,如何利用微斑塊格局及其構件特征評估生態系統整體應對干擾的響應過程和調節機制的研究目前比較薄弱,限制了研究人員對生態系統穩定性維持過程的全面認識。
青藏高原是世界上海拔最高、環境最為惡劣的放牧型生態系統,過度放牧導致生態系統養分輸入和輸出的逆差,干擾了系統固有的養分積累和循環過程。同時,牧民由游牧轉變為定居的時間不足70年,這對于適應了長時間游牧利用方式的草地生態系統如何在短期內迅速調整系統構件,以適應干擾形式的改變是一個挑戰。而近30-40年草地的退化,草地一定程度的空間格局的破碎化,有助于保持生態系統資源斑塊多樣性、生物多樣性,本質上很可能是草地對放牧管理模式、放牧強度改變的適應性調節[114-116]。天然植物群落成分復雜,以放牧草地生態系統微斑塊為研究媒介,確定草地生態系統中可辨識粒度和研究尺度,有助于研究人員從不同層次認識高寒草地生態系統屬性變化的過程和特征,但如何規范草地微斑塊類型和等級,以保證研究實體兼顧表觀特征與內在特征的關聯性、試驗布置的可行性和研究尺度的精準性,是評判微斑塊構件是否能夠作為生態系統研究單元的重要標準。但該項工作的研究涉及學科復雜,領域廣泛,中間未知因素過多也成為限制其研究深入的重要因素。
草地植物群落斑塊源于生境的異質性及植物種間生物學與生態學特征的差異[117-118]。根據等級斑塊動態兼容機制理論(imcorportation)和復合穩定性理論(metastability)[119],斑塊鑲嵌體是連續植物群落不同路徑或起始點的演替結果,其在生態系統中配置格局的變化使天然草地無時不處于變化之中,其變化強度同干擾強度密切相關,即在一定干擾強度下,系統在大尺度斑塊中表現為相對穩定,在小尺度下斑塊表現為相對多變,即天然草地在一定時間范圍內表現出總體穩定而局部變化的特點[120];但隨著干擾強度的加劇,微斑塊格局發生變化,空間異質性增強,景觀破碎度加劇[120-123],系統恢復力變弱,植被群落整體特征改變,植被發生演替[70],導致青藏高原高寒草地出現多穩態共存現象,草地微斑塊的存在某種程度體現了其演化歷史,更反映了生態系統穩定性的維持策略。研究帶有不同歷史時期微斑塊的空間配置格局,分析生態系統構件物質能量潴留、循環和轉化過程,有助于研究人員深刻認識生態系統系統自組織過程。但相關研究仍缺乏理論、方法和技術的支撐[124-126]。
基于草地生態系統的復雜性,微斑塊構件特征的研究需要借鑒景觀生態學、動物學、植物學、微生物學、系統論、機械工程等多種學科的方法、理念、手段,才能全面的認識生態系統,才能更好的為退化生態系統恢復提供理論支撐。
猜你喜歡
數學小靈通·3-4年級(2024年2期)2024-05-15 02:02:28
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:36
瘋狂英語·新策略(2019年10期)2019-12-13 08:43:28
當代陜西(2019年10期)2019-06-03 10:12:04
數學小靈通·3-4年級(2017年9期)2017-10-13 08:10:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
