BCG誘導RAW264.7細胞脂肪酸氧化對細胞自噬和促炎因子表達的調控作用
2021-09-27 09:03:38徐金瑞王玉炯
畜牧獸醫學報 2021年9期
關鍵詞:檢測
駱 佳,徐金瑞,李 武,王玉炯*
(1.西部特色資源保護與利用教育部重點實驗室,銀川 750021;2.寧夏大學生命科學學院,銀川 750021)
結核病(tuberculosis, TB)是由結核分枝桿菌(Mycobacteriumtuberculosis,Mtb)感染引起的慢性呼吸道傳染病,所造成的死亡是全球十大致死原因之一。巨噬細胞作為Mtb的主要宿主細胞和免疫細胞,通過調節自身死亡方式(凋亡、壞死、焦亡和自噬)決定Mtb感染的最終轉歸[1]。細胞凋亡促進胞內菌的清除,細胞壞死則加速胞內滯留菌的擴散,而自噬決定著胞內菌的命運(清除或滯留)[2]。
Mtb感染導致巨噬細胞代謝進程變化,致使脂滴和細胞器脂質聚集在胞質中而形成泡沫巨噬細胞(foamy macrophage)[3]。三酰甘油、膽固醇和脂肪酸是泡沫巨噬細胞中的主要脂質成分,Mtb通過利用這些脂質為其自身提供能源物質從而維持在巨噬細胞中的復制和存活[4-5]。Mtb感染促進肺泡巨噬細胞脂肪酸氧化(fatty acid oxidation, FAO),干預巨噬細胞脂肪酸氧化,抑制Mtb在巨噬細胞中繁殖[6]。此外,炎性介質的產生也與脂肪酸代謝有著密切的聯系[7]。然而,Mtb感染過程中,FAO對巨噬細胞自噬以及促炎因子表達的調控作用還尚不明確。本研究采用牛分枝桿菌疫苗株卡介苗(bacillus Calmette-guérin, BCG)感染小鼠巨噬細胞RAW264.7,用乙莫克舍[8](etomoxir, Eto)抑制FAO,通過檢測自噬相關蛋白表達、自噬小體聚集、自噬流以及促炎因子IL-1β、IL-6和TNF-α的表達情況,揭示BCG感染巨噬細胞過程中,FAO途徑對巨噬細胞自噬和炎性反應的調控作用,為基于代謝途徑的結核病免疫機制研究提供新的思路。
1 材料與方法
1.1 材料
小鼠巨噬細胞RAW264.7購自中國科學院細胞庫;胎牛血清、DMEM-High Glucose購自Gibco公司;細胞總RNA提取試劑盒購自OMEGA公司;TransSciptAll-in One SuperMix for qPCR、PerfectStartGreen q-PCR SuperMix試劑盒購自全式金公司;兔抗LC3、Beclin1、CPT-1A和GAPDH抗體購自Cell Signal Technology(CST);兔抗Rab7抗體和游離脂肪酸定量試劑盒(free fatty acid quantification assay kit)購自Abcam公司;熒光素偶聯山羊抗兔IgG和BODIPY-488購自Invitrogen 公司;乙莫克舍(Etomoxir, Eto)購自MedChemExpress公司;mRFP-GFP-LC3熒光雙標腺病毒購自漢恒生物科技有限公司;小鼠IL-1β、IL-6 和TNF-α ELISA試劑盒購自博士德生物工程有限公司;相關引物由生工生物工程有限公司合成。
1.2 細胞培養
用含10%胎牛血清的DMEM高糖培養基,于37 ℃、5% CO2培養箱中培養RAW264.7細胞。培養細胞至對數生長期,用于后繼試驗。
1.3 BCG培養
牛結核分枝桿菌疫苗株BCG培養于添加Tween-80(0.5%)和增菌劑(OADC)的Middlebrook 7H9培養基中,37 ℃ 5% CO2培養至菌液OD600 nm=1.5后進行傳代培養。
1.4 試驗分組設計
試驗共分3組,分別為對照組(Ctrl)、BCG感染組和Eto與BCG共處理組(Eto+BCG)。Ctrl組為不經任何處理正常狀態下的RAW264.7細胞;BCG組用感染復數為(multiplicity of infection, MOI)10的BCG培養物感染細胞6 h;Eto+BCG組用Eto預處理RAW264.7細胞2 h后,用MOI=10的BCG培養物感染細胞6 h。
1.5 Western blot檢測
Eto預處理RAW264.7細胞2 h后,用MOI=10的BCG培養物感染細胞6 h,提取細胞總蛋白,BCA定量后進行SDS-PAGE凝膠電泳。PVDF膜轉印后,5%脫脂奶粉室溫封閉1 h,過夜孵育 CPT-1A(1∶1 000)、Beclin1(1∶1 000)、LC3-II(1∶1 000)以及Rab7(1∶1 000)抗體,次日TBST(Tween-20含量0.3%)洗膜6次(5 min·次-1),二抗室溫孵育1 h,TBST 洗膜6次后,化學發光檢測蛋白表達水平。
1.6 游離脂肪酸定量試驗
用MOI=10的BCG懸液感染RAW264.7細胞6 h后收集細胞,用含1% Triton X-100的氯仿充分裂解細胞,13 000×g離心10 min,去除不可溶物質并收集有機相。將收集的有機相置于50 ℃烘箱中至氯仿去除后,真空干燥30 min,從而提取細胞中游離脂肪酸(free fatty acid, FFA)。所提取的FFA根據說明書進行定量并與細胞蛋白含量進行歸一化處理。
1.7 BODIPY檢測脂滴
以3×105cell·mL-1細胞量接種于放有蓋玻片的12孔板中,MOI=10的BCG懸液感染RAW264.7細胞6 h后,PBS漂洗細胞3次(5 min·次-1),4%多聚甲醛固定20 min,BODIPY 染色15 min, PBS漂洗3次(5 min·次-1),用含DAPI封片劑封片。
1.8 免疫熒光檢測自噬小體
以3×105cell·mL-1的細胞量接種于放有蓋玻片的12孔板中,Eto預處理RAW264.7細胞2 h后,用MOI=10的BCG培養物感染細胞6 h,PBS漂洗感染后的細胞3次(5 min·次-1),4%多聚甲醛固定20 min,Triton X-100室溫通透20 min,5% 山羊血清室溫封閉30 min,LC3抗體(1∶200)37 ℃孵育2 h,熒光二抗(1∶1 000)37 ℃孵育30 min。用含DAPI的封片劑封片,激光共聚焦顯微鏡觀察細胞中自噬小體聚集情況。
1.9 自噬流檢測
將對數生長期的RAW264.7細胞以2×105cell·mL-1細胞量接種于放置有蓋玻片的12孔板中,過夜培養。mRFP-GFP-LC3熒光雙標腺病毒以感染復數(multiplicity of infection,MOI)50感染6 h,更換新鮮完全培養基繼續培養24 h。Eto預處理2 h,BCG(MOI=10)感染6 h,PBS漂洗3次(5 min·次-1),4%多聚甲醛固定20 min,抗熒光淬滅封片劑封片,激光共聚焦顯微鏡觀察細胞中mRFP-LC3和mRFP-GFP-LC3點狀聚集。
1.10 qRT-PCR檢測IL-1β、IL-6、TNF-α mRNA表達
Eto預處理RAW264.7細胞2 h后,用MOI=10的BCG培養物感染細胞6 h,用細胞總RNA提取試劑盒提取細胞總RNA,并根據反轉錄試劑盒說明書將提取到的RNA反轉錄合成cDNA。以cDNA作為模板進行qRT-PCR操作。qRT-PCR試驗數據用2-ΔΔCt法分析各試驗組中IL-1β、IL-6、TNF-αmRNA的相對表達水平。鼠IL-1β、IL-6、TNF-α及GAPDH上下游引物見表1。
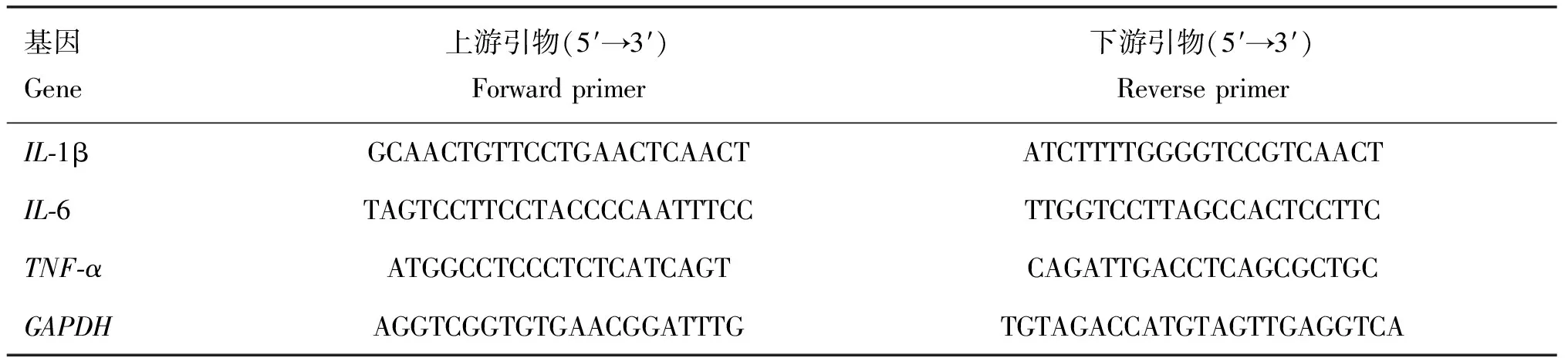
表1 qRT-PCR引物序列
1.11 ELISA檢測IL-1β、IL-6和TNF-α含量
Eto預處理RAW264.7細胞2 h后,用MOI=10的BCG培養物感染細胞6 h,收集細胞培養上清,1 000 r·min-1離心20 min,用小鼠IL-1β、IL-6和TNF-α ELISA試劑盒檢測上清中IL-1β、IL-6和TNF-α含量。
1.12 胞內菌存留試驗
Eto預處理RAW264.7細胞2 h后,用MOI=10的BCG培養物感染細胞2 h,PBS漂洗細胞3次清除胞外菌,繼續培養細胞6 h,0.5% Triton X-100 裂解細胞;待巨噬細胞全部裂解后,將裂解產物10倍稀釋并均勻涂布在含有OADC的Middlebrook7H10 瓊脂培養基上,培養3周后,進行菌落計數。
1.13 統計學分析
所有試驗數據均經過3次獨立試驗的驗證,試驗數據采用GraphPad Prism 9.0中T-test或One Way ANOVA進行統計學分析,數據用平均數±標準誤(x±sx)表示。
2 結 果
2.1 BCG感染促進RAW264.7細胞脂肪酸氧化
BODIPY標記脂質檢測BCG感染前后細胞中脂滴含量,結果顯示:BCG感染引起RAW264.7細胞中脂滴積累(圖1A、B),而游離脂肪酸含量顯著下調(P<0.05)(圖1C);且BCG感染促進了肉毒堿棕櫚酰基轉移酶1A(carnitine palmitoyltransferase 1A, CPT-1A)表達(圖1D),并在BCG感染4 h 時其表達量達到最高。

A、B.BODIPY標記脂質,激光共聚焦觀察BCG感染前后RAW264.7細胞脂滴聚集情況,BODIPY為綠色,DAPI為藍色;C.吸光光度法檢測BCG感染前后,RAW264.7細胞細胞中游離脂肪酸含量; D.Western blot檢測BCG感染不同時間點CPT-1A表達水平。*.P<0.05,**.P<0.01
2.2 抑制BCG誘導RAW264.7細胞FAO抑制RAW264.7細胞中BCG存留量
CFU試驗檢測Eto預處理后,BCG感染的RAW264.7細胞中BCG存活量。結果顯示:Eto預處理后的細胞中BCG存留量較BCG組顯著減少(圖2)(P<0.01)。說明抑制BCG誘導RAW264.7細胞FAO抑制RAW264.7細胞中BCG存活。

**.P<0.01
2.3 BCG誘導RAW264.7細胞FAO參與調控細胞自噬
Western blot檢測自噬相關蛋白Beclin1和LC3-Ⅱ以及溶酶體蛋白Rab7表達變化。結果顯示,BCG感染促進了Beclin1、LC3-Ⅱ和 Rab7的表達;Eto預處理進一步上調Beclin1、LC3-Ⅱ和Rab7的表達,且差異具有統計學意義(P<0.05,圖3A~D)。激光共聚焦結果也顯示,與BCG組相比,Eto預處理后的細胞中聚集大量自噬小體(圖4A、B)。

Eto預處理2 h,BCG感染RAW264.7細胞6 h后,Western blot檢測Beclin1、LC3-Ⅱ和Rab7表達水平。*.P<0.05,**.P<0.01

A.激光共聚焦觀察細胞中自噬小體的聚集情況,LC3-Ⅱ為綠色,DAPI為藍色; B.每個細胞中自噬小體含量定量結果。**.P<0.01
2.4 抑制BCG誘導RAW264.7細胞FAO促進自噬流
激光共聚焦檢測Eto預處理后,BCG感染的RAW264.7細胞中自噬體(mRFP-GFP-LC3)和自噬溶酶體(mRFP-LC3)聚集情況。結果顯示:BCG感染的RAW264.7細胞中出現自噬體mRFP-GFP-LC3和自噬溶酶體mRFP-LC3點狀聚集,且經Eto處理后,RAW264.7細胞中自噬體和自噬溶酶體含量均出現顯著上調(P<0.05,圖5)。說明抑制BCG誘導RAW264.7細胞FAO途徑促進了自噬流發生。
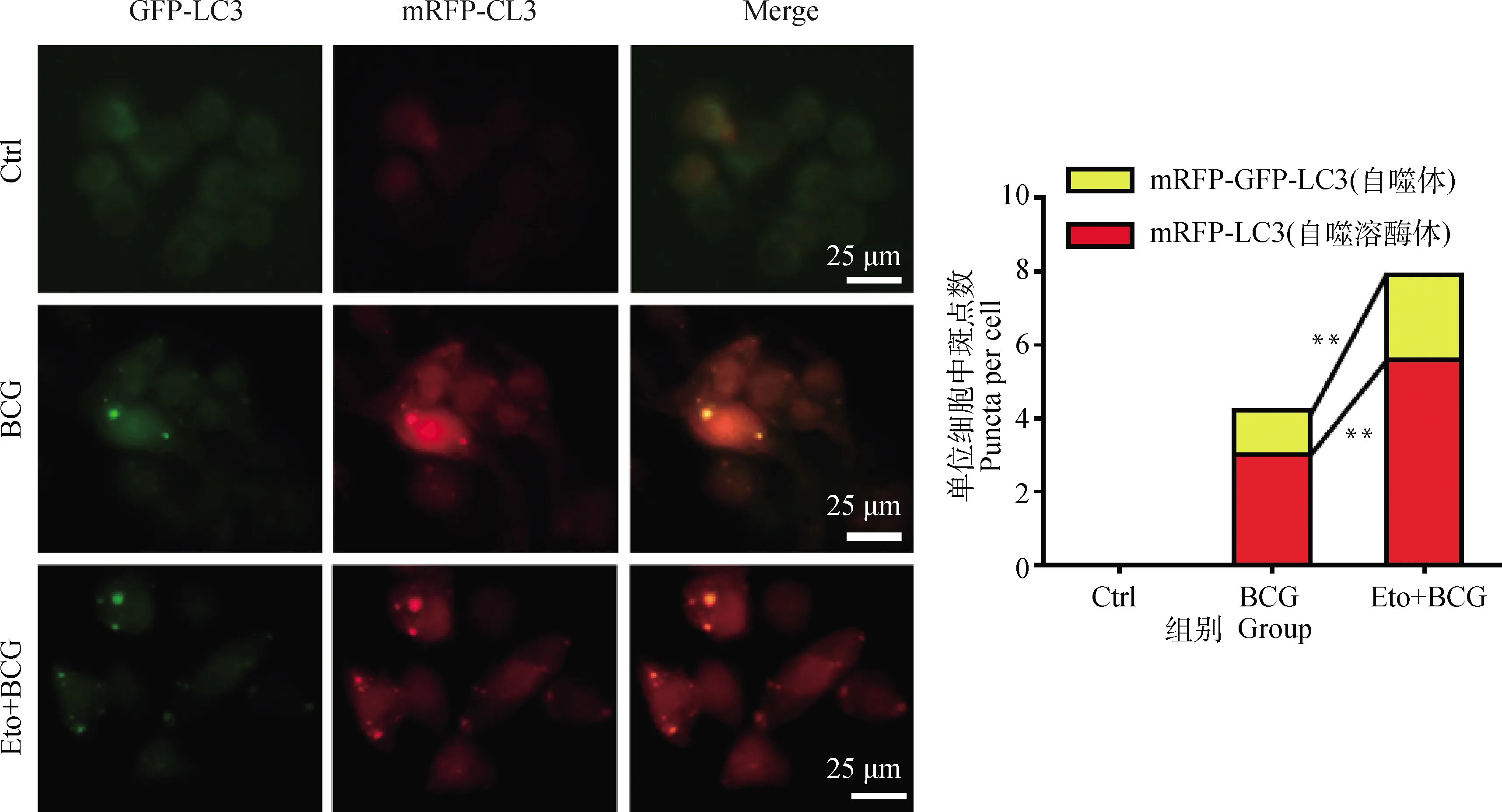
*.P<0.05,**.P<0.01
2.5 抑制BCG誘導RAW264.7細胞FAO下調IL-1β、IL-6和TNF-α表達
qRT-PCR檢測促炎因子IL-1β、IL-6和TNF-αmRNA表達情況。結果顯示,Eto預處理抑制BCG感染引起的IL-1β、IL-6和TNF-αmRNA表達(圖6A~C),且差異均具有統計學意義(P<0.05)。為了進一步探究BCG誘導RAW264.7細胞FAO對促炎因子IL-1β、IL-6和TNF-α表達的影響,收集細胞培養上清,用ELISA方法檢測上清中IL-1β、IL-6和TNF-α含量,試驗結果與qRT-PCR結果一致,即Eto預處理后,上清中IL-1β、IL-6和TNF-α含量較BCG感染組顯著下調(P<0.05,圖6D~F)。

A、B、C.Eto預處理2 h,BCG感染RAW264.7細胞6 h后,qRT-PCR檢測促炎因子IL-1β、IL-6和TNF-α mRNA表達水平;D、E、F.Eto預處理2 h,BCG感染RAW264.7細胞6 h后,收集細胞培養上清,ELISA 檢測細胞培養上清中促炎因子IL-1β、IL-6和TNF-α含量。*.P<0.05,**.P<0.01
3 討 論
早期研究顯示,Mtb利用宿主細胞脂肪酸作為碳源維持其在胞內生長繁殖[9-12]。Podinovskaia等[13]應用熒光脂肪酸轉運試驗觀察到細胞中Mtb內存在宿主細胞來源的脂肪酸,說明Mtb攝取并利用了宿主脂肪酸[14]。在巨噬細胞、小鼠以及人肺組織Mtb感染模型中,參與脂肪酸利用相關基因表達上調[15-17]。本研究用BODIPY染色標記脂滴,發現BCG感染的RAW264.7細胞中出現大量脂滴聚集,而細胞中游離脂肪酸含量減少,且參與FAO的關鍵酶——CPT-1A表達上調。此結果說明:BCG感染促進了巨噬細胞FAO。
Mtb感染導致的巨噬細胞代謝變化將影響吞噬體與溶酶體的融合[18-19],從而為Mtb在宿主細胞中長期滯留提供了良好的條件。由此可見,改變Mtb與巨噬細胞代謝間的平衡機制可抑制胞內病原菌存活[20]。肉芽腫是結核病的主要病理特征,感染了Mtb的巨噬細胞在其胞漿區域積聚脂質,并分化形成泡沫巨噬細胞[21-22],構成了肉芽腫主要成分之一。泡沫巨噬細胞在Mtb感染擴散中發揮著重要作用。因此,調控巨噬細胞脂代謝進程可以控制結核病的發生與發展。目前,基于宿主細胞代謝的宿主導向治療(host-directed therapy, HDT)方法在人結核病的治療中,尤其在感染多重耐藥結核分枝桿菌患者的治療中取得了顯著效果[23-24]。自噬作為一種溶酶體清除機制,在調控巨噬細胞對Mtb的清除中發揮著重要的作用[25]。多種因素調控細胞自噬,代謝途徑改變所導致的能量變化則是其中因素之一[26-28]。FAO作為細胞供能方式之一,其變化也將影響巨噬細胞自噬清除作用。CPT-1A通過介導長鏈脂肪酸進入線粒體,進而進行FAO。Eto通過與CPT-1A結合,進而抑制FAO的進行[29]。為探究BCG誘導的RAW264.7細胞FAO對細胞自噬和促炎因子的表達調控作用,本研究用Eto抑制FAO并檢測BCG感染后自噬相關蛋白Beclin1和LC3-II以及炎性因子IL-1β、IL-6和TNF-α的表達情況,結果顯示,Beclin1和LC3-II的表達均顯著上調,且細胞中出現大量自噬小體;而IL-1β、IL-6和TNF-α的表達卻受到抑制。Rab7是自噬體與溶酶體融合進程中重要調節分子之一[30],參與調控自噬溶酶體的成熟過程,敲除Rab7將導致自噬小體聚集[31]。結果顯示,Eto預處理不僅上調了自噬相關蛋白的表達,同時促進了Rab7的表達,說明抑制FAO可以促進自噬小體和溶酶體融合,并且自噬流檢測結果也進一步顯示干預FAO后,細胞中自噬溶酶體數量增多,胞內存留BCG也顯著下調。由此可見,BCG誘導FAO參與調控巨噬細胞自噬,抑制FAO可以激活RAW264.7自噬途徑清除細胞中滯留的病原菌,并且可以防止因為過度炎性反應所致的炎性損傷。然而,FAO調控巨噬細胞自噬的具體機制將是本課題接下來要探究的問題。
4 結 論
BCG感染促進RAW264.7細胞FAO,且此代謝途徑參與調控RAW264.7細胞自噬:抑制BCG誘導的RAW264.7細胞FAO促進了細胞自噬從而抑制胞內菌的存活,且抑制了促炎因子IL-1β、IL-6和TNF-α的表達和分泌。說明FAO途徑在巨噬細胞抗結核分枝桿菌感染中發揮著雙重調控作用。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
