炒酸棗仁對焦慮大鼠基底外側杏仁核神經元信息編碼動態變化的影響*
2021-10-09 04:21:34華伊張正一王艷艷黃莉莉葉曉楠李廷利
醫藥導報 2021年10期
關鍵詞:實驗
華伊,張正一,王艷艷,黃莉莉,葉曉楠,李廷利
(黑龍江中醫藥大學藥學院,哈爾濱 150040)
酸棗仁為鼠李科植物酸棗[ZiziphusjujubaMill.Var.spinosae(Bunge)Hu ex H.F.Chou]的干燥成熟種子,主治虛煩不眠、驚悸多夢等癥[1],酸棗仁在臨床的應用普遍使用炒制品。現代研究發現炒酸棗仁能減少行為學測試中動物的焦慮樣行為,具有一定的抗焦慮作用[2-3]。腦科學領域普遍認為杏仁核(amygdala)是參與調節焦慮相關的情緒、行為和生理反應的一個重要腦區[4-5],杏仁核的基底外側核團(basolateral amygdala,BLA)是焦慮調節過程中接收刺激信息傳入的主要部分[6]。杏仁核功能的改變已被認為是焦慮癥的一個潛在條件[7-8],焦慮行為的增加與BLA區的過度活躍有關[9]。近年來,BLA區作為神經回路中治療焦慮相關狀態和行為的關鍵成分被廣泛研究[10-12]。筆者在本文將行為學與神經元在體同步電活動相結合,進行了焦慮大鼠的行為學及BLA區神經元信息編碼動態變化的研究,同時考察炒酸棗仁對其的干預作用。
1 材料與方法
1.1實驗動物 無特定病原體(SPF)級SD大鼠,雄性,體質量(220±20)g,購于黑龍江中醫藥大學GLP實驗動物中心,實驗動物生產許可證號:SCXK(黑)2013-004。動物飼養與實驗環境為通風、避光、隔音、電磁屏蔽狀態,并通過自動定時光控系統進行12 h/12 h明暗光照周期處理(每日7:00—19:00開啟照明,19:00至次日7:00關閉照明);室內溫度:20~24 ℃;相對濕度:50%~60%;噪聲≤40 dB。實驗前所有大鼠在本實驗室環境內適應7 d,整個實驗過程中所有大鼠不限制飲食和飲水。
1.2藥物與試劑 酸棗仁,原產地山西,由黑龍江中醫藥大學王振月教授鑒定為正品,依照文獻[13]方法,制備炒酸棗仁和炒酸棗仁水煎液;0.9%氯化鈉溶液(哈爾濱三聯藥業有限公司,批號:170104D5);戊巴比妥鈉(國藥集團化學試劑有限公司,批號:WS20140104);義齒基托聚合物(上海貝瓊齒材有限公司,批號:170308);義齒基托樹脂(上海新世紀齒科材料有限公司,批號:20170408)。
1.3儀器與設備 大鼠獨立通氣籠IVC(型號:RU25H5,蘇州市蘇杭科技器材有限公司),分析天平(型號:MS105,梅特勒-托利多儀器上海有限公司,感量:0.01 mg),大鼠曠場實驗箱(100 cm×100 cm×40 cm,木制),標準腦立體定位儀(型號:51600,美國Stoelting公司),在體多通道神經記錄分析系統(型號:OPX-32D,美國Plexon公司),在體單軸微操作儀(型號:IVM-1000,Microelettrica Scientifica China)。
1.4在體多通道記錄電極的埋植 選取SD雄性大鼠20只,進行電極埋植手術前,所有大鼠禁食8 h,不禁水。戊巴比妥鈉(40 mg·kg-1)腹腔注射麻醉大鼠后,將其固定在標準腦立體定位儀上。根據《George Paxinos & Charles Watson大鼠腦立體定位圖譜》(第3版),定位BLA區位置(坐標AP 2.5~2.8 mm,ML 4.5~4.8 mm,DV 7.5 mm),將16(4×4)通道在體記錄電極(鉑銥合金,電極絲外周涂有絕緣層,直徑35 μm)埋置于大鼠的BLA區域,并用義齒基托樹脂將電極部分固定在動物顱頂。術后動物恢復4~5 d,于有機玻璃籠內單獨飼養。
1.5動物的分組及給藥 將已植入電極并經過術后恢復的大鼠經隨機數字表法分為模型對照組與炒酸棗仁組,每組10只。炒酸棗仁組每日灌胃給予大鼠炒酸棗仁水煎液(17.5 g·kg-1),模型對照組給予同等體積純化水,每日1次,連續7 d。
1.6模型的制備及行為學測試 利用曠場實驗復制大鼠焦慮模型,每日給藥0.5 h后,將各組大鼠放入曠場箱內底面中心,任其自由探索,利用動物行為學采集和分析系統,記錄10 min內大鼠在曠場中的行為學表現,每日1次,連續7 d。測試指標為跨格次數、站立次數、進入中央格次數、中央格運動時間(s)和總運動時間(s),并以中央格運動時間除以總運動時間計算中央格/總運動時間(%)。
1.7神經元在體放電信號的采集與處理 每日給藥0.5 h后,利用在體多通道神經記錄分析系統,先采集各組大鼠在日常飼養環境中BLA區神經元基礎放電信號(10 min),再在曠場實驗的同時采集曠場環境中的BLA區神經元曠場放電信號(10 min),每日1次,連續7 d。利用Offline Sorter軟件對采集到的神經元放電信號進行處理,過濾低頻放電和環境干擾放電,設置信噪比>3:1的信號為所需的動作電位信號,運用主成分分析(principal component analysis,PCA)技術對提取出的動作電位進行分類處理,并計算每一類神經元動作電位的放電頻率。將每只大鼠的每一類神經元動作電位在曠場中的放電頻率減去其基礎放電頻率,得出的值定義為神經元動作電位放電頻率增加值,以此值進行兩組大鼠在曠場中神經元動作電位放電頻率變化的對比。
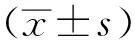
2 結果
2.1炒酸棗仁對焦慮大鼠行為學動態變化的影響 隨著測試天數的增加,大鼠跨格次數、站立次數及總運動時間逐漸下降,進入中央格次數變化平穩,中央格運動時間、中央格/總運動時間(%)逐漸上升。與模型對照組比較,炒酸棗仁組大鼠跨格次數在第5天明顯減少(P<0.05);站立次數在第2,4,5,7天明顯減少(P<0.05或P<0.01);總運動時間1~7 d均無明顯變化(P>0.05);進入中央格次數在第1天明顯增加(P<0.01);中央格運動時間在第1~第3,第6天明顯增加(P<0.05或P<0.01);中央格/總運動時間(%)在第1,第3、第5~第7天明顯增加(P<0.05或P<0.01)。見圖1。
①與模型對照組比較,P<0.05;②與模型對照組比較,P<0.01。
2.2炒酸棗仁對焦慮大鼠BLA區神經元信息編碼動態變化的影響
2.2.1大鼠BLA區神經元動作電位的分類及特征圖 在大鼠BLA區記錄到的神經元動作電位波形特征為:波長較長,波峰后曲線下降緩慢。按照放電特征的不同,把在BLA區記錄到的神經元分為Ⅰ、Ⅱ兩類(以下稱BLA區Ⅰ類神經元為NeuronⅠ,Ⅱ類神經元為NeuronⅡ)。NeuronⅠ的時長較長、振幅稍低,自相關圖中出現典型的尖峰、峰值后表現出快速的指數型下降;NeuronⅡ的時長較短、振幅較高,自相關圖中峰值后下降緩慢。兩類神經元動作電位放電模式的特征圖見圖2。

A.NeuronⅠ動作電位;B.NeuronⅡ動作電位;a.波形圖;b.放電間隔直方圖;c.自相關圖。
2.2.2焦慮大鼠BLA區神經元動作電位放電頻率的動態變化 與自身的基礎放電頻率相比,模型對照組和炒酸棗仁組大鼠在曠場中NeuronⅠ、NeuronⅡ動作電位放電頻率在1~7 d均有所增加;NeuronⅠ、NeuronⅡ在曠場中動作電位放電頻率在1~2 d均有所增加,然后在2~7 d逐漸下降。NeuronⅠ與NeuronⅡ的放電數比例約為2:1。各組放電頻率直方圖見圖3。

底部未畫黑線部分為大鼠處于日常飼養環境中的基礎放電,畫黑線部分為大鼠處于曠場環境中的曠場放電。
2.2.3炒酸棗仁對焦慮大鼠BLA區神經元動作電位放電頻率動態變化的影響 由于與自身的基礎放電頻率相比,每只大鼠在曠場中的NeuronⅠ、NeuronⅡ動作電位放電頻率在1~7 d均有所增加,以神經元動作電位放電頻率增加值進行兩組大鼠在曠場中動作電位放電頻率變化的對比。實驗結果表明,與模型對照組比較,炒酸棗仁組大鼠BLA區NeuronⅠ的動作電位放電頻率增加值在第1,3~6天明顯減少(P<0.05或P<0.01),NeuronⅡ的動作電位放電頻率增加值在第1,3,4,6,7天明顯減少((P<0.05或P<0.01)。見圖4。
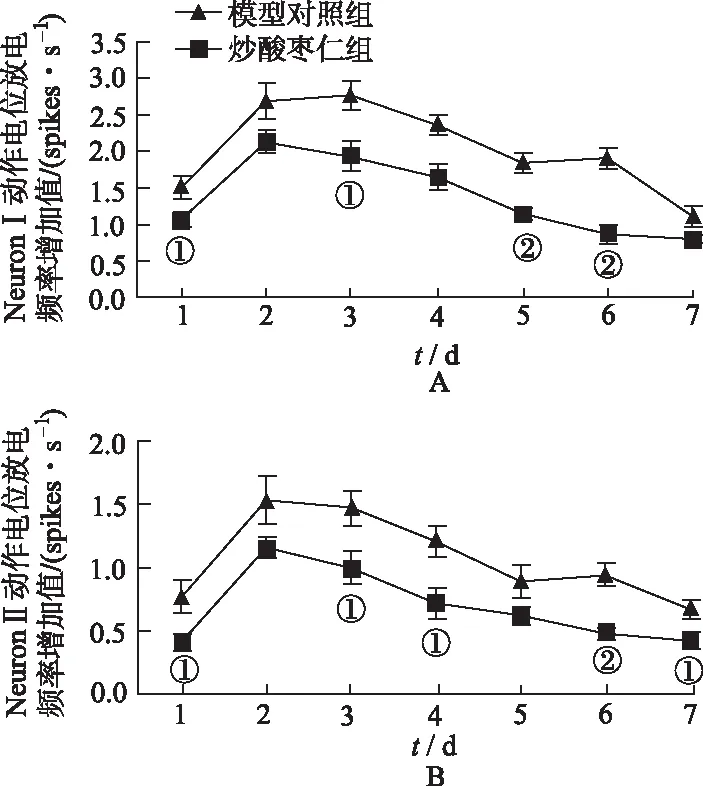
①與模型對照組比較,P<0.05;②與模型對照組比較,P<0.01。
3 討論
嚙齒動物面對曠場提供的新環境時,同時產生對空曠環境的恐懼和對新奇環境的探索欲[14],曠場實驗利用這種矛盾沖突心理,在造成動物焦慮情緒的同時,進行運動性和焦慮水平的評估[15-16]。本文利用曠場實驗制備大鼠焦慮模型,考察了連續7 d大鼠焦慮狀態下的行為學動態變化。曠場行為學結果顯示,大鼠的運動性相關指標,包括跨格次數、站立次數及總運動時間,隨著測試時間的增加均逐漸減少。這一現象反映,隨實驗時間的增加,大鼠對曠場環境逐漸適應從而減弱它們對曠場的好奇心,降低大鼠的探索行為。
曠場環境的中央格區域對動物意味著潛在威脅,周邊格相較而言更安全,動物在中央格區域的探究行為越多,代表動物的焦慮程度越低[17]。連續7 d的行為學實驗結果顯示,大鼠的焦慮相關指標,包括中央格運動時間及中央格運動時間/總運動時間(%),隨著測試時間的增加而逐漸增加,表明動物的焦慮程度逐漸減輕。大鼠的焦慮樣行為在初次接觸曠場時表現得更為強烈,而焦慮水平隨著重復接觸曠場而逐漸降低。這些觀察結果證明,動物在反復暴露于曠場環境后表現出習慣化行為,與文獻[18]報道一致。HSIAO等[19]報道,連續進行4 d的曠場測試,動物在中心格區域運動時間會逐漸減少,即反復接觸曠場后焦慮水平會增加,與本文的實驗結果有所差異,可能是因為每天動物進行50 min的曠場實驗,而本文每日只測試10 min。因此,推測重復暴露于致焦慮環境中的時間過長不會建立習慣化行為,反而會造成焦慮程度增加。
本實驗在利用曠場實驗制備大鼠焦慮模型的同時,記錄連續7 d大鼠BLA區在體神經元的基礎放電及在曠場環境中曠場放電情況。本文采集到的BLA區神經元動作電位的波形特征為波長較長,波峰后曲線下降緩慢,與文獻[20]報道結果一致。將在BLA區收集到的神經元信號按動作電位的波形和放電特征分為Ⅰ、Ⅱ兩類,與基礎放電頻率相比,所有大鼠的NeuronⅠ、NeuronⅡ在連續7 d的曠場環境中其動作電位放電頻率均增加。這一現象表明,曠場環境改變大鼠BLA區的神經元信息編碼,NeuronⅠ、NeuronⅡ均參與了曠場對動物的焦慮加工。
通過對BLA區神經元動作電位放電頻率增加值變化的觀察,發現NeuronⅠ、NeuronⅡ的動作電位放電頻率在1~2 d均有所增加,然后在2~7 d持續下降。文獻[20]報道小鼠在連續3 d在曠場環境中BLA區神經元動作電位放電頻率持續增加,本研究在1~2 d的結果與上述報道相似。這一現象提示,反復暴露于曠場環境中可能會引發BLA區神經元突觸可塑性的短期增強,提高BLA區神經元的興奮性。本研究結果顯示從第2天開始BLA區神經元動作電位放電頻率呈下降趨勢,與WANG等[20]研究存在差異,推測造成此差異的原因與在曠場中的暴露時間有關。WANG等[20]研究中動物每日進行30 min的曠場實驗,而本文每日只測試10 min,提示重復接觸致焦慮的曠場環境的時間過長不會建立習慣化行為,反而會造成動物焦慮水平的逐漸增加,這與行為學結果的分析相同,但仍需進一步的研究確定。本文第2~7天的結果證明,由于測試次數的不斷增加,大鼠建立習慣性行為,逐漸減輕在曠場中的焦慮程度,從而降低了BLA區神經元的放電頻率,與行為學測試的結果一致。
本研究在觀察焦慮大鼠行為學和神經元在體活動動態變化的同時,給予炒酸棗仁水煎液干預,探討炒酸棗仁對其的影響作用。行為學結果顯示,炒酸棗仁減少大鼠在第5,7天的跨格次數,并減少了第2,4,5,7天的站立次數。這一現象表明炒酸棗仁能降低重復暴露于曠場環境中大鼠的運動性和探索行為。同時,炒酸棗仁增加大鼠第1天的進入中央格次數,增加第1~3,6天的中央格運動時間,并增加了第1,3,5~7天的中央格/總運動時間,表明炒酸棗仁能減輕曠場對大鼠造成的焦慮情緒。由于重復暴露于曠場環境中,大鼠建立了習慣化行為,焦慮水平隨著測試次數增加而降低,然而炒酸棗仁仍然能降低重復測試中動物的焦慮水平,提示炒酸棗仁的抗焦慮效果遠遠超過了動物自身的適應能力。
神經元在體電活動實驗結果顯示,炒酸棗仁在第1,3~6天減少了大鼠NeuronⅠ動作電位放電頻率增加值,在第1,3,4,6,7天減少了大鼠NeuronⅡ動作電位放電頻率增加值。表明炒酸棗仁通過抑制BLA區神經元的過度放電改變BLA區神經元信息編碼,拮抗曠場對大鼠造成的焦慮狀態。此外,在某些測試時間,炒酸棗仁能明顯減少BLA區神經元電活動,但動物的焦慮樣行為并沒有顯著變化。推測是由于炒酸棗仁的抗焦慮作用并不是由某一腦區的一類神經元介導,可能是多個腦區或多類神經元共同作用的結果,具體原因還需進一步研究。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
