睪丸酮叢毛單胞菌中多種蛋白與SDRx的生物學(xué)作用研究
2021-10-28 01:03:30張海鷗楊鑫蕾尉建嵇冶
張海鷗,楊鑫蕾,尉建,嵇冶
(長(zhǎng)春理工大學(xué) 生命科學(xué)技術(shù)學(xué)院,長(zhǎng)春 130022)
隨著工業(yè)的發(fā)展,工廠所排放的大量甾體類(lèi)化合物,對(duì)人類(lèi)和動(dòng)物的生長(zhǎng)發(fā)育帶來(lái)巨大的危害[1-2]:如引發(fā)人類(lèi)不孕不育、生殖疾病、腫瘤及生物變性等嚴(yán)重問(wèn)題[3]。近年科學(xué)家在自然界發(fā)現(xiàn)了屬于叢毛單胞菌屬的睪丸酮叢毛單胞菌(Comamonastestosteroni,CT)ATCC11996,它 是以甾體作為唯一碳源的革蘭氏陰性細(xì)菌,并以甾體激素作為碳源和能源物質(zhì),進(jìn)行代謝活動(dòng)。在之前的研究中已報(bào)道了CT的基因組序列,并描述和表征了幾種甾體代謝酶與甾體結(jié)合的反應(yīng)過(guò)程[4]。CT菌分解甾體激素的過(guò)程錯(cuò)綜復(fù)雜,一共有幾十種酶先后共同參與了它的代謝過(guò)程[5],SDR短鏈脫氫酶就是眾多代謝酶家族中的一員。
過(guò)去研究表明LuxR、LysR、TetR蛋白對(duì)睪丸酮叢毛單胞菌(CT菌)中的很多脫氫酶具有調(diào)控作用[6-8],它們可以與短鏈脫氫酶及甾體激素互相結(jié)合并對(duì)甾體激素進(jìn)行降解。因此研究CT菌中SDRx脫氫酶的調(diào)控作用具有重要的研究?jī)r(jià)值。本研究采用基因克隆、重組質(zhì)粒構(gòu)建、質(zhì)粒共轉(zhuǎn)化、ELISA、高效液相等方法,研究這三種蛋白對(duì)SDRx的調(diào)控作用,從而為更好的利用CT菌修復(fù)環(huán)境中甾體激素的污染的研究做貢獻(xiàn),可為睪丸酮叢毛單胞菌在自然界中甾體激素的生物降解及基因調(diào)控研究提供實(shí)驗(yàn)依據(jù)[9-12]。
1 實(shí)驗(yàn)材料
CT菌,pCR2.1-TOPO自殺質(zhì)粒由德國(guó)基爾大學(xué)熊光明教授惠贈(zèng)。質(zhì)粒小提試劑盒來(lái)自上海Songon公司,DNA凝膠回收試劑盒來(lái)自北京TIAN GEN公司,睪丸酮、孕酮、雌酮、17a-乙炔基雌二醇、甲睪酮均來(lái)自德國(guó)Dr.Ehrenstorfer公司,羊抗鼠抗體來(lái)自上海Chemicon公司。大腸桿菌DH5α為實(shí)驗(yàn)室保存,pK18、pUC19質(zhì)粒均為實(shí)驗(yàn)室所有;重組質(zhì)粒pUC19-LuxR為實(shí)驗(yàn)室保存。所用化學(xué)試劑均為分析純。
2 實(shí)驗(yàn)方法
2.1 SDRx基因位點(diǎn)及其引物設(shè)計(jì)
SDRx基因?qū)儆?-氧酰基-(酰基載體蛋白)還原酶(3-oxoacyl-[acyl-carrier-protein]-reduc‐tase)一員,以NAD+或NADP+為受體、作用于供體CH-OH基團(tuán)上的氧化還原酶,如圖1所示,它位于 glutathione-dependent formaldehy-de-activating下游344 kb處,在CT菌基因組中具有相同的轉(zhuǎn)錄方向。作為SDR超家族的一員,該SDR基因包含SDR的共同序列基序,N末端Gly-XXX-Gly-XGly輔因子結(jié)合基序和Tyr-XXX-Lys區(qū)段。
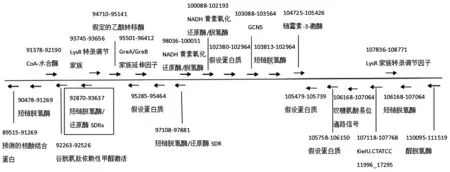
圖1 SDRx基因位點(diǎn)
2.2 SDRx敲除菌株構(gòu)建及活性測(cè)定
參照張昊等人[13-14]的方法,設(shè)計(jì)SDRx基因201-500bp中心部位的敲除引物時(shí),在上游第三、四堿基之間加入G(P1)并在下游末端引入終止子TAA(P2),引物列表如表1所示,并進(jìn)行PCR擴(kuò)增,將PCR片段克隆至pCR2.1-TOPO中,得到pCR2.1-TOPO-SDR300,電轉(zhuǎn)到CT菌感受態(tài)細(xì)胞中,27℃,180 rpm條件下置于Amp+/Kan+雙抗平板,得到多個(gè)SDRx敲除株,利用P1和TOPO質(zhì)粒的敲除通用驗(yàn)證引物(P3),以敲除株基因組為模板進(jìn)行PCR驗(yàn)證。利用基因同源重組將目的基因?qū)隒T菌基因組中,由于TOPO質(zhì)粒的自殺特性和引物加G進(jìn)行移碼突變從而達(dá)到基因敲除目的。
用終濃度為 0.5 μmol/L的睪丸酮(T)、孕酮(P)、雌酮(ES)、雌二醇(E2)、甲睪酮(ME),對(duì)野生型、敲除株進(jìn)行誘導(dǎo)培養(yǎng),并設(shè)置空白對(duì)照組(相同培養(yǎng)基體系下只加入等濃度的激素不加菌液)。每種激素在27℃,180 rpm/min條件下誘導(dǎo)16 h,用等體積的氯仿萃取,重復(fù)3次,收集氯仿后通風(fēng)櫥中晾干,分別溶于1 mL甲醇作為樣品,島津液相色譜儀進(jìn)行HPLC分析。檢測(cè)波長(zhǎng)分別為雌二醇280 nm;睪丸酮280 nm;孕酮280 nm;甲睪酮208 nm;雌酮241 nm;流動(dòng)相水與甲醇的比為20∶80;每次進(jìn)樣50 μL;色譜柱的溫度為40℃。
2.3 SDRx脫氫酶表達(dá)載體構(gòu)建及目的活性蛋白的表達(dá)純化
以野生型CT菌基因組為模板進(jìn)行PCR得到目的基因片段SDRx,擴(kuò)增引物P4、P5(見(jiàn)表1)。連接至pET-15b載體上,構(gòu)建重組表達(dá)質(zhì)粒pETSDRx,轉(zhuǎn)入至表達(dá)菌株 BL21(DE3)中,得到疑似陽(yáng)性克隆子并進(jìn)行測(cè)序及雙酶切驗(yàn)證,無(wú)突變成功構(gòu)建表達(dá)工程菌。

表1 引物設(shè)計(jì)
對(duì)pET-15b和pET-SDRx進(jìn)行大量表達(dá),在IPTG終濃度0.8 mmol/L條件下進(jìn)行誘導(dǎo),16℃,110 rpm,恒溫震蕩培養(yǎng)24 h。分別收取菌液后,PBS重懸3次,凍融超聲,鎳柱純化獲得目的蛋白,將所得蛋白進(jìn)行SDS-PAGE鑒定。
2.4 SDRx蛋白多克隆抗體的制備及效價(jià)測(cè)定
取得小鼠陰性血清后,對(duì)其進(jìn)行特異性免疫。將上述工程蛋白過(guò)濾,分三次進(jìn)行免疫,第一次免疫將工程蛋白與完全弗氏佐劑1∶1混合,第二、三次免疫將工程蛋白與不完全弗氏佐劑1∶1混合,每次免疫間隔時(shí)間為一周,完成第三次免疫后,進(jìn)行眼球取血,得到陽(yáng)性血清。
ELISA法測(cè)定SDRx小鼠多克隆抗體效價(jià),加入陰性血清為陰性對(duì)照(N),加入抗原作為陽(yáng)性標(biāo)準(zhǔn)(P),P/N>2.1時(shí)該比例為抗體效價(jià)。將抗原稀釋至2 μg/mL 37℃包被1 h,洗滌液清洗3次后,每孔加入100 μL(1∶8 000),37 ℃包被1 h;洗滌3次,每孔加入100 μL Chemicon公司購(gòu)得的二抗羊抗鼠(1∶4 000)37 ℃包被1 h,洗滌液清洗5次,去離子水洗滌2次,加入TMB應(yīng)用液作為顯色液避光包被30 min,加入終止液后,酶標(biāo)儀450 nm波長(zhǎng)測(cè)OD值,空白對(duì)照為490 nm。制備標(biāo)準(zhǔn)曲線并得到線性回歸方程y=0.068 1 ln(x)+0.270 7,R2=0.999 5,得到最終抗體效價(jià)為1∶800 0。
2.5 三種調(diào)控蛋白對(duì)SDRx脫氫酶調(diào)控作用的研究
參照熊光明的實(shí)驗(yàn)方法[15],引物設(shè)計(jì)如表1(P6-P11)所示,將LysR、TetR的PCR片段分別轉(zhuǎn)入pUC19;SDRx與在其前端107bp的預(yù)測(cè)啟動(dòng)子區(qū)域(107SDRx)片段轉(zhuǎn)入pK18,活化實(shí)驗(yàn)室保存的pUC19-LuxR菌種。向HB101感受態(tài)細(xì)胞中加入等濃度pK18-107SDRx質(zhì)粒與pUC19-LysR質(zhì)粒,重組為L(zhǎng)ysR共轉(zhuǎn)株,命名“pLysR”;pK18-107SDRx質(zhì)粒與pUC19-TetR質(zhì)粒,重組為T(mén)etR共轉(zhuǎn) 株 ,命 名“pTetR”;pK18-107SDRx質(zhì) 粒 與pUC19-LuxR質(zhì)粒,進(jìn)行共轉(zhuǎn)化,重組為L(zhǎng)uxR共轉(zhuǎn)株,命名“pLuxR”。Amp+/Kan+雙抗平板篩選出多個(gè)陽(yáng)性克隆子,并以其基因組為模板進(jìn)行PCR驗(yàn)證,成功構(gòu)建共轉(zhuǎn)化菌株。以睪丸酮為誘導(dǎo)激素,取上述陽(yáng)性克隆子活化擴(kuò)大后,加入終濃度 0.5 μmol/L睪丸酮,37℃ 180 rpm條件下誘導(dǎo)14 h,提取總蛋白,設(shè)置對(duì)照組與平行試驗(yàn)組各三組,對(duì)照組為相同體系下無(wú)水乙醇進(jìn)行誘導(dǎo)的重組菌株,平行試驗(yàn)組分別為相同體系下終濃度為0.5 μmol/L的孕酮、睪丸酮進(jìn)行誘導(dǎo)的重組菌株,分別提取總蛋白,方法參照2.3部分。
將所得蛋白稀釋至1 mg/mL進(jìn)行包被,野生型總蛋白稀釋為2 mg/mL進(jìn)行包被。上述抗體以1∶8 000的效價(jià)進(jìn)行一抗包被。ELISA方法詳細(xì)步驟參照上述2.4.實(shí)驗(yàn)部分。將抗原稀釋至2 μg/mL 37℃包被1 h,洗滌液清洗3次后,每孔加入 100 μL(1∶8 000),37 ℃包被 1 h;洗滌 3次,每孔加入100 μL Chemicon公司購(gòu)得的二抗羊抗鼠(1∶4 000)37℃包被1 h,洗滌液清洗5次,去離子水洗滌2次,加入TMB應(yīng)用液作為顯色液避光包被30 min,加入終止液后,酶標(biāo)儀450 nm波長(zhǎng)測(cè)OD值,空白對(duì)照為490 nm。利用t檢驗(yàn)及方差分析的統(tǒng)計(jì)學(xué)方法對(duì)三組數(shù)據(jù)進(jìn)行分析,將LysR、LuxR、TetR重組共轉(zhuǎn)質(zhì)粒與pUC19重組共轉(zhuǎn)質(zhì)粒分組后對(duì)其顯著性差異進(jìn)行分析。
3 結(jié)果與分析
3.1 SDRx基因突變株(M-CT)鑒定
如圖2原理圖所示,利用基因同源重組將目的基因?qū)隒T菌基因組中,由于TOPO質(zhì)粒的自殺特性和引物加G進(jìn)行移碼突變從而達(dá)到基因敲除目的。SDRx敲除株P(guān)CR鑒定如圖3可知,1道以M-CT基因組為模板,在530 bp處有明亮單一條帶,而相應(yīng)2道以野生型CT為模板并未出現(xiàn)對(duì)應(yīng)條帶,證明SDRx基因的敲除株M-CT構(gòu)建成功。
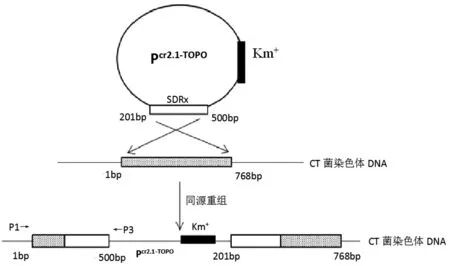
圖2 SDRx敲除原理圖
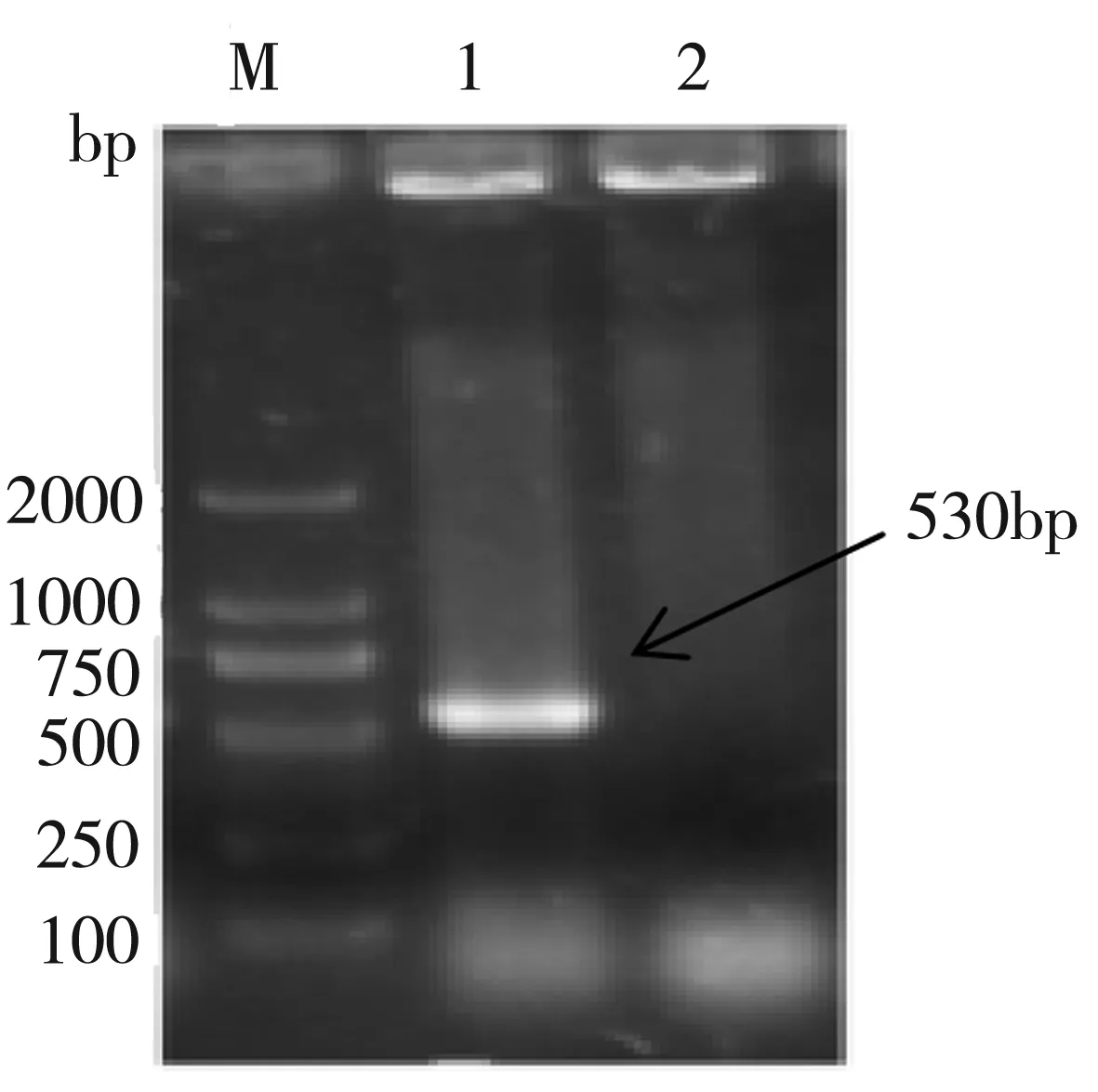
圖3 SDRx敲除株P(guān)CR鑒定圖
3.2 高效液相數(shù)據(jù)分析
高效液相結(jié)果如圖4可知,為野生型CT與敲除株M-CT的三組平行實(shí)驗(yàn)均值,加入終濃度為0.5 μmol/L的不同種類(lèi)甾體激素,其中敲除突變株在各激素培養(yǎng)條件下,降解量均明顯少于野生型,SDRx基因敲除后其降解激素的能力受到了很大的影響,柱狀圖顯示了CT菌中SDRx敲除后,對(duì)不同激素的降解能力影響程度為睪丸酮>甲睪酮>孕酮>雌酮>雌二醇。
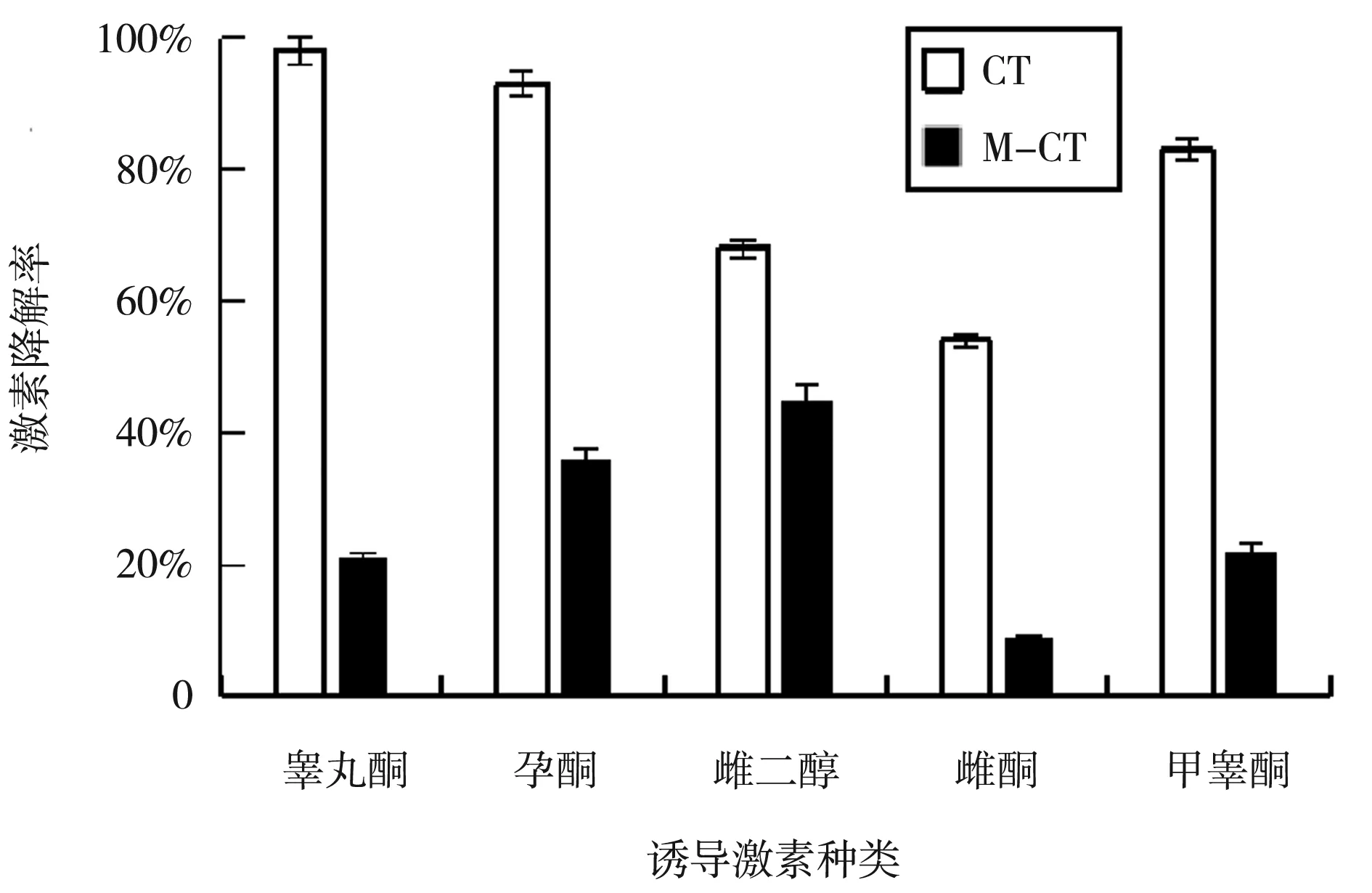
圖4 野生型及敲除株激素降解量數(shù)據(jù)圖
3.3 SDRx目的蛋白的表達(dá)純化結(jié)果圖
利用上述方法構(gòu)建SDRx表達(dá)載體,將疑似陽(yáng)性克隆子質(zhì)粒進(jìn)行雙酶切并送由庫(kù)美公司進(jìn)行測(cè)序鑒定,基因序列未發(fā)生突變,并將陽(yáng)性克隆子質(zhì)粒轉(zhuǎn)化到大腸桿菌BL21(DE3)p-LysS中,得到正確無(wú)突變的pET-SDRx-BL21(DE3)表達(dá)菌株。
經(jīng)軟件Protein Molecular Weight對(duì)SDRx的查詢計(jì)算,帶有組氨酸標(biāo)簽的重組蛋白分子量為29.2 kDa。如圖5所示,第10泳道為NPI-200下洗脫的目的蛋白,條帶清晰大小準(zhǔn)確為29.2 kDa,純化效果良好。
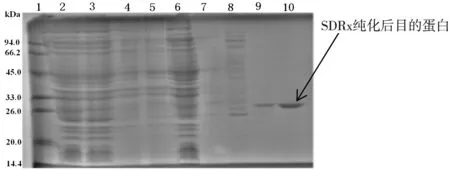
圖5 SDRx蛋白聚丙烯酰胺凝膠電泳表征結(jié)果圖
3.4 LysR,LuxR,TetR三種調(diào)控因子重組質(zhì)粒驗(yàn)證
圖6為pK18-107SDRx上啟動(dòng)子和SDRx基因的結(jié)構(gòu)示意圖。圖7為以上述構(gòu)建成功的pK18-107SDRx分別與pUC19-LysR/LuxR/TetR的共轉(zhuǎn)菌株基因組為模板的PCR驗(yàn)證結(jié)果,其中1道的LysR為912 bp;2道的LuxR為1 125 bp;3道的TetR為660 bp,均為理想數(shù)值,即表明成功構(gòu)建了三種共轉(zhuǎn)化質(zhì)粒。
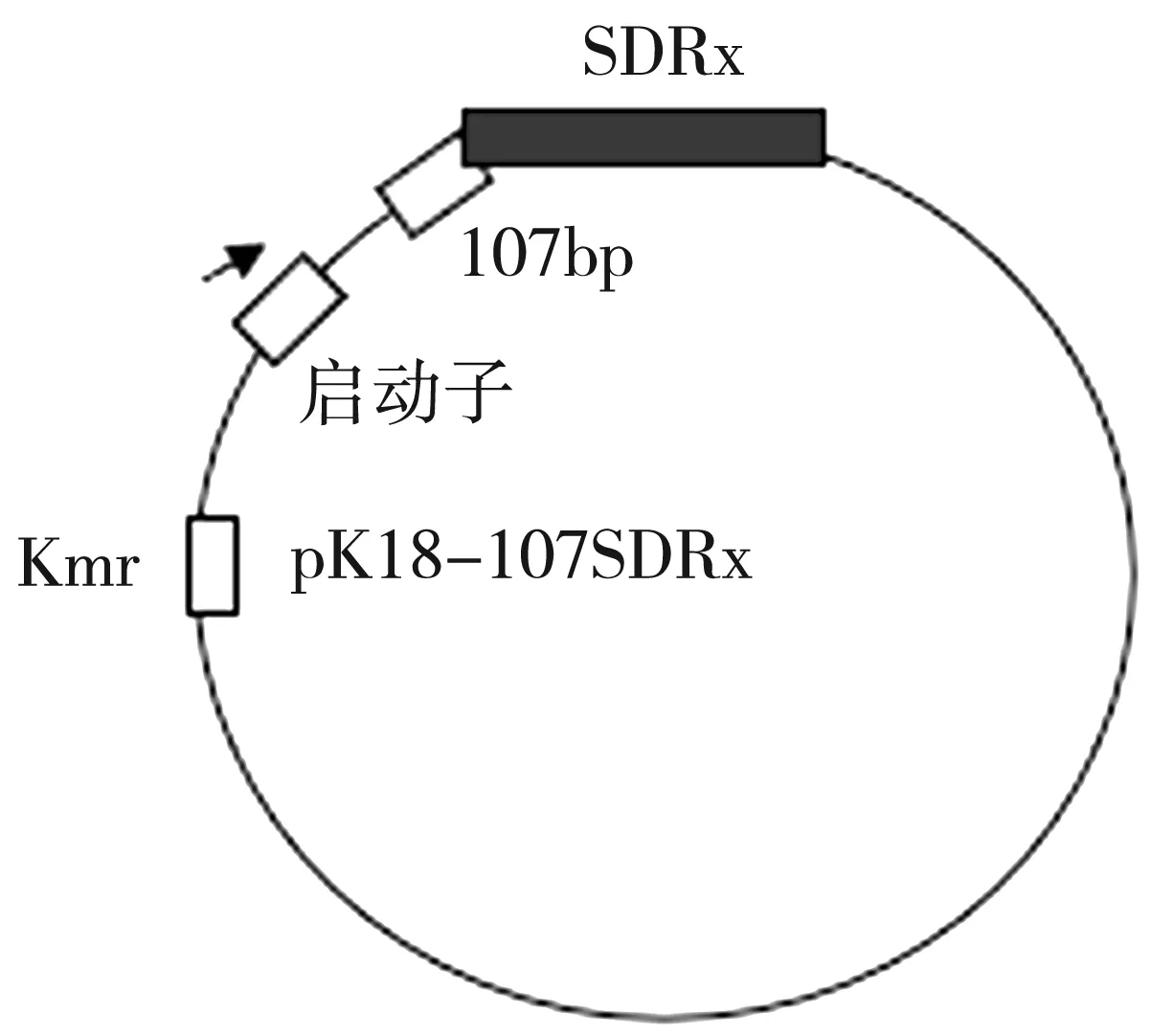
圖6 pK18-107SDRx上啟動(dòng)子和SDRx基因的結(jié)構(gòu)示意圖
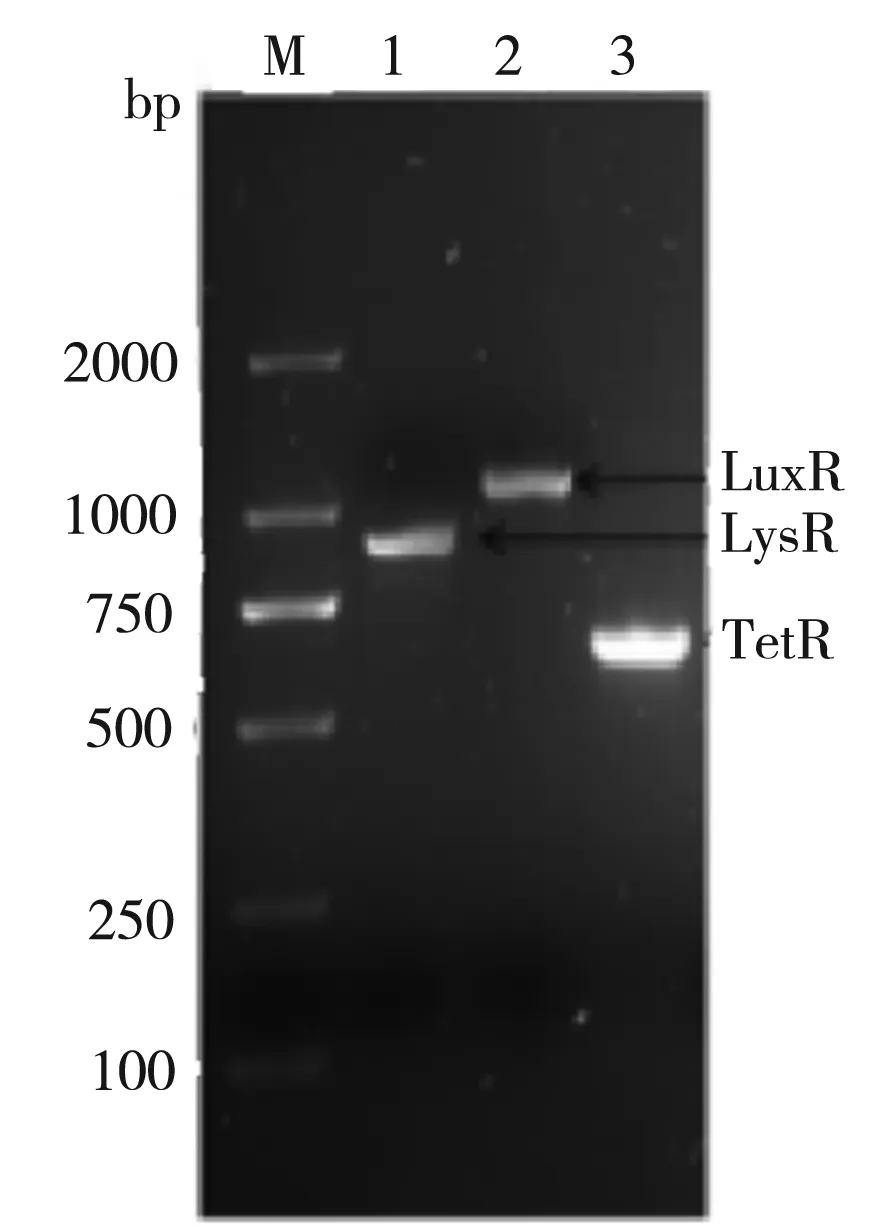
圖7 LysR,LuxR,TetR重組質(zhì)粒PCR鑒定
3.5 三種調(diào)控蛋白對(duì)SDRx脫氫酶調(diào)控作用的結(jié)果圖
由3.2實(shí)驗(yàn)結(jié)果,即野生型CT菌對(duì)睪丸酮、甲睪酮及孕酮三種激素的降解能力較好,本實(shí)驗(yàn)選取了較有代表性的睪丸酮和孕酮作為底物,如圖8所示。利用t檢驗(yàn)及方差分析的統(tǒng)計(jì)學(xué)方法對(duì)三組數(shù)據(jù)進(jìn)行分析,將LysR、LuxR、TetR重組共轉(zhuǎn)質(zhì)粒與pUC19重組共轉(zhuǎn)質(zhì)粒分組后對(duì)其顯著性差異進(jìn)行分析。與無(wú)調(diào)控蛋白的共轉(zhuǎn)化菌株(pUC19空白對(duì)照組)數(shù)據(jù)對(duì)比,LysR調(diào)控蛋白在兩種激素條件下對(duì)SDRx脫氫酶的表達(dá)量基本不變;LuxR和TetR調(diào)控蛋白在兩種激素條件下對(duì)SDRx脫氫酶的表達(dá)起促進(jìn)作用,且睪丸酮誘導(dǎo)條件下的促進(jìn)作用尤為顯著,其中睪丸酮條件下TetR的促進(jìn)作用更明顯大于LuxR。
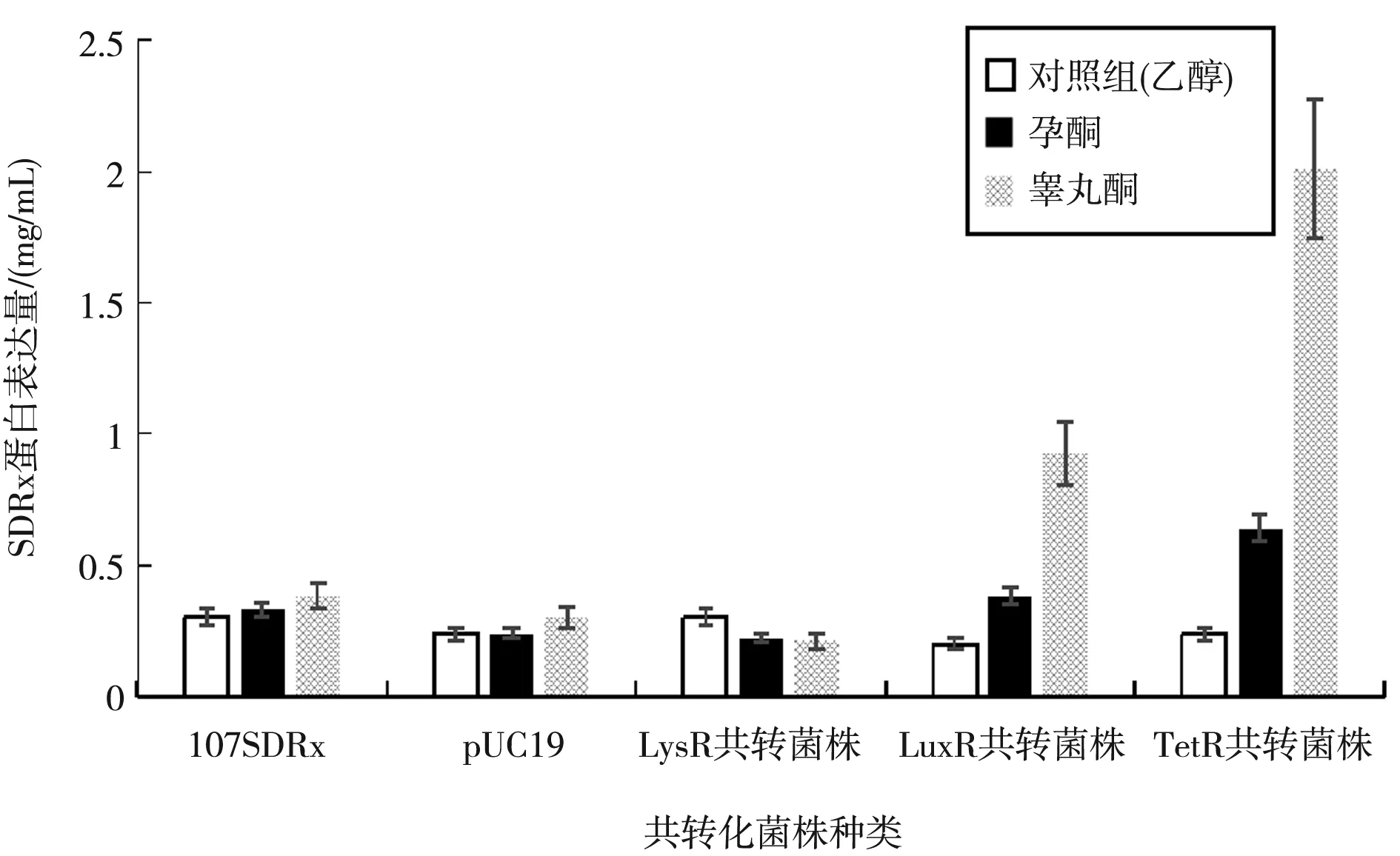
圖8 三種調(diào)控蛋白共轉(zhuǎn)菌株中SDRx蛋白的表達(dá)情況
4 結(jié)論與展望
本研究中的降解激素結(jié)果表明:敲除株對(duì)甾體類(lèi)激素的降解能力均呈現(xiàn)出不同程度的減弱而不是完全喪失,這與之前發(fā)表的BKR,AKR,LuxR等基因敲除株[16-17]出現(xiàn)的情況相類(lèi)似。可能是由于CT菌本身有一些同工酶或代償途徑,使得SDRx基因被敲除后,也會(huì)有其他途徑(或酶)來(lái)完成激素降解,表現(xiàn)為其降解能力的減弱而不是消失。
LysR,LuxR,TetR分別對(duì)SDRx的調(diào)控結(jié)果可以得出如下結(jié)論:(1)在誘導(dǎo)物為睪丸酮時(shí):與無(wú)調(diào)控蛋白的共轉(zhuǎn)菌株(pUC19空白對(duì)照組)相比,“pTetR”的SDRx表達(dá)量提高了5.3倍,明顯表現(xiàn)為正向促進(jìn)作用;“pLuxR”的SDRx表達(dá)量提高了2.1倍,增加并不明顯,但仍有正向促進(jìn)調(diào)控作用;而“pLysR”的SDRx表達(dá)量則降低了0.3倍,可忽略不計(jì),表明沒(méi)有調(diào)控作用;(2)當(dāng)誘導(dǎo)物為孕酮時(shí),與無(wú)調(diào)控蛋白的共轉(zhuǎn)菌株(pUC19空白對(duì)照組)相比,“pTetR”的SDRx表達(dá)量提高了1.6倍,表現(xiàn)為正向促進(jìn)作用,但效果明顯低于在睪丸酮的條件下;“pLuxR”、“pLysR”的 ELI‐SA結(jié)果對(duì)比無(wú)調(diào)控蛋白的共轉(zhuǎn)菌株(pUC19空白對(duì)照組)分別為提高了的0.58倍、降低了0.08倍,數(shù)值較小可忽略不計(jì),沒(méi)有表現(xiàn)出調(diào)控作用。
在前人的工作中證明了CT菌中某些調(diào)控蛋白可以與誘導(dǎo)物、被調(diào)控基因三者之間的相互作用關(guān)系,并構(gòu)建了動(dòng)態(tài)調(diào)控模型[15],而本實(shí)驗(yàn)中又進(jìn)一步證明了TetR能夠?qū)DRx脫氫酶產(chǎn)生正調(diào)控。目前,對(duì)CT菌中甾體激素的全部代謝網(wǎng)絡(luò)尚處于逐漸了解的過(guò)程中,在今后的工作中,隨著研究的不斷深入,還有幾個(gè)問(wèn)題亟需解決:(1)TetR作為調(diào)控蛋白,如何結(jié)合到SDRx的調(diào)控區(qū)?(2)作為誘導(dǎo)物的睪丸酮如何結(jié)合到TetR蛋白上?結(jié)合位點(diǎn)在哪里?這個(gè)結(jié)合中心空間大小與睪丸酮(孕酮)分子是否匹配?(3)睪丸酮和孕酮在作為誘導(dǎo)物時(shí),三種調(diào)控蛋白分別與SDRx共轉(zhuǎn)化后的ELISA結(jié)果所表現(xiàn)出的差異,是否是由于其與誘導(dǎo)物結(jié)合中心的匹配效果不同所引起的?(4)用分子互做的方法來(lái)檢測(cè)TetR蛋白與睪丸酮分子的親和常數(shù)是多少?(5)當(dāng)TetR蛋白結(jié)合了睪丸酮后,它與SDRx基因的結(jié)合區(qū)又是否會(huì)產(chǎn)生變化?(6)CT菌中有大量的調(diào)控基因存在,TetR調(diào)控蛋白是否會(huì)與其他調(diào)控蛋白共同作用于SDR家族并對(duì)其產(chǎn)生調(diào)控?(7)所用的檢測(cè)方法是否可以應(yīng)用于環(huán)境中甾體類(lèi)激素的微量檢測(cè)等。隨著研究的深入,可以更好地了解分析CT菌的代謝作用機(jī)理,構(gòu)建出安全可靠、降解能力更強(qiáng)大的基因工程菌株。
