冬蟲夏草調控PERK-eIF2α信號通路抑制香煙煙霧提取物刺激下的人支氣管上皮細胞凋亡研究*
2021-11-08 02:24:40祝慶華李敏芳唐明文
中國中醫(yī)急癥 2021年10期
關鍵詞:檢測
劉 玉 謝 緯 祝慶華 李敏芳 唐明文 陳 生
(廣東省深圳市中醫(yī)院,廣州中醫(yī)藥大學第四臨床醫(yī)學院,廣東 深圳 518003)
慢性阻塞性肺疾病(COPD)是一種持續(xù)進展的以氣流受限為特征的疾病,嚴重危害人類健康,與接觸有毒顆粒和氣體的顯著暴露引起的氣道和肺泡異常有關,致殘率和死亡率極高,社會經濟負擔重,已成為一個重要的公共衛(wèi)生問題[1]。COPD的發(fā)病機制復雜,目前臨床用于治療COPD的藥物難以取得滿意的療效。
香煙煙霧刺激引起肺內氧化應激是吸煙導致COPD的始動環(huán)節(jié),而內質網應激(ERS)是造成肺組織細胞凋亡促進COPD發(fā)生發(fā)展的重要機制[7]。冬蟲夏草是我國傳統(tǒng)藥用真菌,具有補肺益腎、止血化痰等功效,主要用于治療久咳虛喘、勞嗽咯血、陽痿遺精、腰膝酸痛等癥[2]。現代藥理研究證實,冬蟲夏草主要成分含有腺苷、多種氨基酸、甘露醇、麥角醇、糖及多種維生素、微量元素等,具有抗氧化、抗炎、免疫調節(jié)等藥理作用[3-4]。大量臨床研究也顯示,冬蟲夏草治療肺部疾病具有確切臨床療效[5-6]。有研究發(fā)現,冬蟲夏草可干預糖尿病、腎病動物模型ERS發(fā)展[8-9],但冬蟲夏草是否能通過抵抗ERS誘導的肺泡上皮細胞凋亡而發(fā)揮治療COPD尚不完全明確。本實驗采用香煙煙霧提取物(CSE)誘導A549細胞方法建立體外COPD細胞模型,通過冬蟲夏草干預COPD細胞模型,觀察細胞凋亡和內質網應激信號通路p-PERK-eIF2α相關蛋白水平的變化,探討冬蟲夏草調控內質網應激信號通路PERK-eIF2α抑制香煙煙霧提取物刺激下的人支氣管上皮細胞凋亡,為冬蟲夏草的合理應用提供實驗基礎。
1 材料與方法
1.1 細胞株 人肺泡上皮細胞(A549細胞,廣州華韻生物科技有限公司)。
1.2 試劑與儀器 DMEM培養(yǎng)基、胎牛血清、胰蛋白酶(0.25%)(美國Gibco公司,批號分別為11966025、10099141、25200056);磷酸鹽緩沖(PBS,美國Hyclone公司,批號:SH3025601);Annexin V-FITC/PI細胞凋亡檢測試劑盒(Solarbio,批號:CA1020);Hoechst 33342染色液(中國碧云天公司,批號分別為C0017、C1022);中強度RIPA裂解液(弗德生物,批號:SBJ-0226);BAC蛋白定量試劑盒(弗德生物,批號:FD2001);PERK抗體(affinity,批號:AF5304);PERK抗體(affinity,批號:DF7576);eIF2α(affinity,批號:AF6087);p-eIF2α(af?finity,批號:AF3087)。1500型全波長酶標儀、371型CO2細胞培養(yǎng)箱,美國Thermo Scientific公司;超凈工作臺,蘇州精華安泰技術有限公司;倒置顯微鏡,日本Olympus公司;恒溫培養(yǎng)振蕩器,天津市歐諾儀器儀表有限公司;蛋白電泳及轉膜設備,北京六一生物科技有限公司;凝膠成像儀,海門市其林貝爾儀器制造有限公司。
1.3 蟲草菌液的準備 對質量濃度0.99 g/mL的蟲草菌液(購自杭州中美華東制藥有限公司)進行過濾除菌,細胞干預之前用含10%胎牛血清的H-DMEM培養(yǎng)基將蟲草菌液稀釋成10、50、100、200 mg/L[10-11]。
1.4 香煙煙霧提取物的制備 將3支去過濾嘴香煙依次連于自制的抽吸裝置[12]中點燃,負壓吸引,吸入的煙霧經過入口3 mL的PBS中制成懸液,密閉搖勻,經0.22濾膜濾器過濾除菌。為確保每次制備及干預所用CSE質量濃度相同,在初次制備CSE時,使用分光光度儀在CSE最佳吸光波長(270~280 nm)范圍內確定其初始吸光度值,保證每次制備的CSE吸光度值相同。制備好的CSE原液用含10%胎牛血清RPMI-MI-1640培養(yǎng)基按濃度0.5%、1%、2%、5%稀釋,為確保CSE中有效成分,每次干預前30 min制備,采用0.22 μm過濾器過濾。
1.5 MTT法測定CSE干預濃度和時間 分別接種A549細胞于5個96孔板中培養(yǎng),每個培養(yǎng)板每孔分別加入100 μL用含10%胎牛血清培養(yǎng)基稀釋的0.5%、1%、2%、5%CSE溶液,每個干預孔均設置5個復孔,同時設空白孔(不含細胞和CSE的培養(yǎng)基)和對照孔(含細胞的培養(yǎng)基,無CSE)。5個培養(yǎng)板分別培養(yǎng)6、10、16、24、48 h,PBS沖洗,每孔加10 μL MTT(5 mg/mL),于培養(yǎng)箱中繼續(xù)避光培養(yǎng)4 h,酶標儀震蕩,吸去上清,每孔加100 μL DMSO,用錫紙包裹避光,吸光度570 nm處測量各孔的吸光度值并記錄。計算細胞存活率(%)=(實驗孔-空白組)/(對照組-空白組)×100%。根據存活率確認后續(xù)實驗最佳CSE質量濃度和作用時間。
1.6 LDH法檢測冬蟲夏草菌液對A549細胞的損傷接種細胞至96孔板,每孔100 μL,密度大約為2×105個/mL,置培養(yǎng)箱預培養(yǎng)24 h,分組設對照組(含細胞的培養(yǎng)基)、樣品最大酶活性對照組(用于后續(xù)裂解的細胞孔)和冬蟲夏草組(蟲草菌液濃度0、10、50、100、200 mg/L),各組細胞置于細胞培養(yǎng)箱培養(yǎng)24 h,在檢測時點前1 h,在樣品最大酶活性對照孔中加入20 μL LDH釋放劑,培養(yǎng)箱中孵育1 h,離心,每孔取上清液120 μL至新的96孔板,按照LDH檢測說明書操作,測定490 nm波長處吸光度值(A490值)。LDH活力(%)=(A試驗組-A對照組)/(A細胞最大酶活性對照-A對照組)×100%。
1.7 MTT法檢測蟲草菌液和CSE對A549細胞活力的影響 接種細胞至96孔板,每孔100 μL,密度大約為2×105個/mL。細胞分組設空白組(不含細胞的培養(yǎng)基)、對照組(含細胞的培養(yǎng)基)、CSE組、冬蟲夏草組(蟲草菌液含量10、50、100 mg/L)。CSE組加入100 μL 2%CSE,冬蟲夏草組加入100 μL蟲草菌液和100 μL 2%CSE,孵育24 h后,PBS沖洗,每孔加入10 μL MTT(5 mg/mL),避光培養(yǎng)4 h,吸去上清,每孔加入100 μL DMSO,酶標儀震蕩,吸光度570 nm處測量各孔的吸光度值并記錄。計算細胞存活率(%)=(實驗組-空白組)/(對照組-空白組)×100%。
1.8 細胞培養(yǎng)和分組 將A549細胞接種于含10%的DMEM培養(yǎng)液(含10%胎牛血清,100 U/mL青霉素和鏈霉素),于37℃、5%CO2培養(yǎng)箱中培養(yǎng),每隔2~3 d換液1次,倒置顯微鏡下觀察細胞貼壁達到80%~90%時,加入0.25%胰蛋白酶消化、傳代。將A549細胞分為空白對照組、CSE組、冬蟲夏草組。空白組加入培養(yǎng)基,CSE組加入2%CSE,冬蟲夏草組加入蟲草菌液(100 mg/L)和2%CSE。各組均孵育24 h。
1.9 Hoechst 33342染色檢測細胞凋亡 各組細胞根據以上方法進行處理后,吸棄原有培養(yǎng)基,加入100 μL Hoechst 33342染色液。將細胞置于37℃、5%二氧化碳培養(yǎng)箱中繼續(xù)培養(yǎng)20~30 min,吸棄染色液,用PBS或培養(yǎng)液洗滌2~3次后用熒光顯微鏡進行觀察。A549細胞表現為染色質聚集,核固縮和凋亡小體的出現被認為是凋亡細胞。各組細胞隨機挑選3個視野進行凋亡細胞及總細胞計數,實驗重復3次。凋亡率=凋亡細胞數/總細胞數×100%。
1.10 流式細胞術儀檢測細胞凋亡 根據Annexin V-FITC/PI凋亡檢測試劑盒說明書操作。將處理后的各組細胞進行消化、離心、洗滌后置于流式管中,使用 400 μL 1×Annexin V 結合液重懸細胞,加入 5 μL Annexin V染液,混勻,2~8℃避光孵育15 min;加入10 μL PI染液,2~8℃避光孵育5 min。使用流式細胞儀檢測凋亡。
1.11 Western blotting檢測內質網應激通路蛋白表達 分組的各組細胞進行以上干預后,PBS洗滌3遍細胞裂解液,置于冰上30 min。離心取上清液,BCA法測定蛋白濃度并定量,經聚丙烯酰胺變性凝膠電泳分離蛋白,用濕轉法將蛋白轉移至聚偏氟乙烯膜,50 mol/L脫脂牛奶封閉,分別加入 p-PERK、PERK、p-eIF2α、eIF2α、BAX、Bcl抗體(1∶2 000),4 ℃過夜,TBST洗3次,加對應1∶5 000稀釋的辣根過氧化物酶標記的二抗,室溫孵育2 h,TBST洗3次,使用ECL-plus試劑盒發(fā)光,通過生化檢測系統(tǒng)成像并分析結果,以β-actin作為內參照,檢測目的蛋白表達情況。
1.12 統(tǒng)計學處理 應用SPSS22.0統(tǒng)計軟件。實驗數據表示為(SD),多組間差異使用單因素方差分析進行評估,LSD檢驗比較兩組間的差異。P<0.05為差異有統(tǒng)計學意義。
2 結果
2.1 CSE對A549細胞存活率的影響 用不同濃度的CSE處理A549細胞不同時間,細胞的存活率均呈下降趨勢,且CSE干預的時間越長、濃度越大,細胞存活率下降越明顯。根據MTT法測定的細胞存活率繪制生存曲線(圖1),參照IC50和曲線下降趨勢,確定2%CSE干預24 h用于后續(xù)的實驗。
2.2 冬蟲夏草液對正常A549細胞的毒性作用 見表1。細胞上清LDH檢測結果顯示,200 mg/L蟲草菌液對正常培養(yǎng)的A549細胞可能存在毒性,其他質量濃度的蟲草菌液(10~100 mg/L)對正常培養(yǎng)的A549細胞無明顯毒性。
表1 各組細胞上清LDH活性比較(U/L,±s)
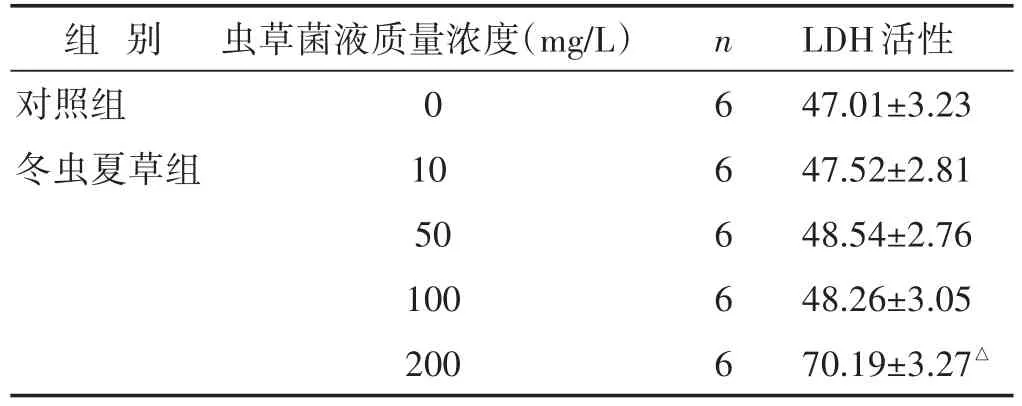
表1 各組細胞上清LDH活性比較(U/L,±s)
注:與對照組比較,△P<0.05。
組別對照組冬蟲夏草組蟲草菌液質量濃度(mg/L)0 10 50 100 200 n 666 6 6 LDH活性47.01±3.23 47.52±2.81 48.54±2.76 48.26±3.05 70.19±3.27△
2.3 冬蟲夏草液有效干預質量濃度篩選 見表2。MTT法檢測結果顯示,與空白組比較,CSE干預組細胞活力顯著降低(P<0.01);與CSE干預組比較,蟲草50、100 mg/L可顯著提高細胞存活率(P<0.01),其中100 mg/L最為顯著,而10 mg/L無明顯影響,故后續(xù)實驗采用冬蟲夏草100 mg/L進行。
表2 各組細胞存活率比較(%,±s)

表2 各組細胞存活率比較(%,±s)
注:與空白組比較,△P<0.05;與CSE組比較,*P<0.05。下同。
組別空白組CSE組冬蟲夏草組質量濃度(mg/L)--1 0 50 100 n66666細胞存活率103.00±3.01 51.49±3.87△55.23±2.93△81.52±3.76*95.37±4.22*
2.4 Hoechst 33342染色檢測冬蟲夏草對細胞凋亡的影響 見圖2,表3。結果顯示,空白組的細胞較少有染色質凝集,核萎縮現象,凋亡小體較少;與空白組相比,CSE組細胞出現核凝聚,細胞皺縮,Hoechst 33342染色明顯加深等,凋亡小體較多(圖2b),且凋亡率顯著升高,與空白組的凋亡率比較,差異具有統(tǒng)計學意義(P<0.01);與CSE組相比,冬蟲夏草組細胞皺縮和凝集明顯減輕,凋亡小體也明顯減少(圖2c),與CSE組的凋亡率比較,具有統(tǒng)計學差異(P<0.01)。
表3 各組Hoechst 33342染色檢測A549細胞凋亡率比較(%,±s)

表3 各組Hoechst 33342染色檢測A549細胞凋亡率比較(%,±s)
組別空白組CSE組冬蟲夏草組質量濃度(mg/L)-2%100 n 666凋亡率4.26±0.97 54.21±1.13△10.33±1.02*

圖2 各組Hoechst 33342染色檢測A549細胞凋亡情況(200倍)
2.5 流式細胞檢測冬蟲夏草對細胞凋亡的影響 見圖3,表4。流式細胞檢測結果顯示,與正常對照組比較,CSE組處理使細胞凋亡率升高至12.7%(P<0.05);與CSE組比較,冬蟲夏草干預可使細胞凋亡率降低至3.8%,兩組細胞凋亡率相比具有統(tǒng)計學差異(P<0.05)。
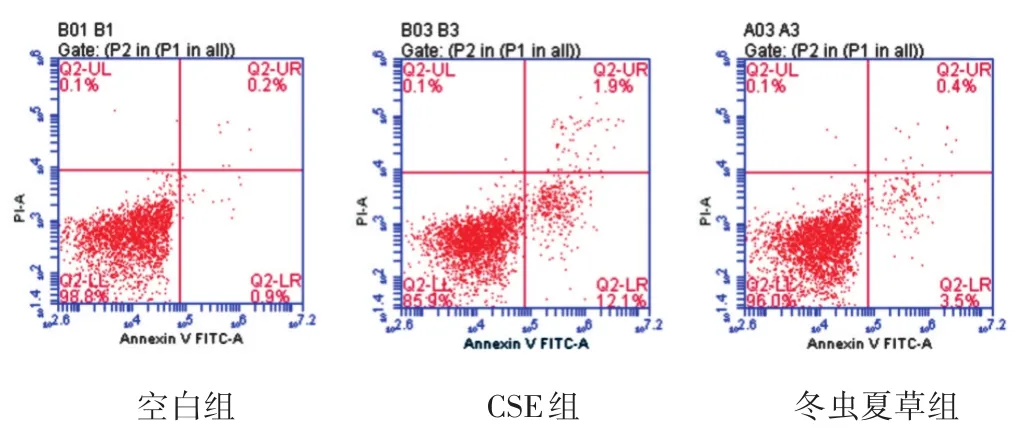
圖3 各組流式細胞檢測A549細胞凋亡情況
表4 各組流式細胞檢測A549細胞凋亡率比較(%,±s)

表4 各組流式細胞檢測A549細胞凋亡率比較(%,±s)
組別空白組CSE組冬蟲夏草組質量濃度(mg/L)-2%100 n 6 6 6凋亡率1.00±0.22 12.70±0.81△3.80±0.64*
2.6 冬蟲夏草對內質網應激分子表達的影響 見圖4,表5。Western blotting結果顯示,與正常組相比,CSE組 p-eIF2α、p-PERK、Bax蛋白表達水平上調(P<0.05),Bcl-2表達下調,Bax/Bcl-2比值升高(P<0.05);與CSE組比較,冬蟲夏草組p-eIF2α、Bax蛋白表達水平下調(P<0.05),Bcl-2表達上調(P<0.05),Bax/Bcl-2比值降低(P<0.05)。
表5 各組A549細胞PERK、p-PERK、eIF2α、p-eIF2α、Bax、Bcl-2表達水平比較(%,±s)

表5 各組A549細胞PERK、p-PERK、eIF2α、p-eIF2α、Bax、Bcl-2表達水平比較(%,±s)
組別空白組CSE組冬蟲夏草組n 6 6 6 p-PERK/PERK 0.05±0.03 0.45±0.04△0.39±0.05 p-eIF2α/eIF2α 0.07±0.05 0.99±0.07△0.51±0.06*Bax/β-actin 0.97±0.05 1.43±0.06△1.02±0.04*Bcl-2/β-actin 0.92±0.07 0.89±0.08 1.73±0.06*Bax/Bcl-2 0.92±0.06 1.41±0.07△0.43±0.05*

圖4 A549細胞PERK、p-PERK、eIF2α、p-eIF2α、Bax、Bcl-2的蛋白表達
3 討 論
ESR廣泛存在于應激的真核細胞內,當應激過于強烈或持續(xù)時間長,內質網穩(wěn)態(tài)不能恢復時,未折疊蛋白反應可激活細胞內凋亡信號上調凋亡分子表達,最終引起細胞內的級聯反應,誘導細胞凋亡[13-14]。ESR誘導細胞凋亡主要有IRE1、PERK、ATF6 3條通路,PERK是內質網應激反應時的首先活化途徑,主要促進細胞的自噬和凋亡。活化后PERK可磷酸化其下游分子eIF2α,抑制促存活蛋白Bcl-2和抗凋亡基因Bcl-2相關的X蛋白(Bax)表達,改變細胞的氧化狀態(tài),提高細胞對凋亡的敏感性[15]。細胞凋亡控制基因Bcl-2家族主要包含Bcl-2和Bax,研究表明,Bcl-2升高,抑制細胞凋亡,Bax增高,促進細胞凋亡,Bcl-2和Bax的比值決定著細胞受凋亡刺激后的生存能力[16-17]。大量研究表明ERS介導的細胞過度凋亡參與了COPD肺結構破壞[18-20],提示調節(jié)ERS信號通路抵制細胞凋亡或將是治療COPD的有效途徑。
本研究采用2%CSE處理A549細胞24 h,與空白組比較,細胞凋亡顯著增加,且ERS信號通路蛋白peIF2α、p-PERK的表達上調,抗凋亡蛋白Bcl-2與促凋亡蛋白Bax比值明顯降低,說明出現明顯ERS相關凋亡,和文獻[7,21]中CSE劑量和時間不完全一致,但結果具有一致性。
冬蟲夏草是中醫(yī)補益肺腎法治療COPD的一味名貴中藥,具有悠久的用藥歷史。中醫(yī)學認為冬蟲夏草性味甘、平,主要歸肺、腎兩經,功效主要有益肺腎,止咳嗽,補虛損,益精氣。冬蟲夏草被廣泛用于呼吸系統(tǒng)疾病治療,有研究提示其保護作用可能與內質網應激有關[22]。為了證實冬蟲夏草的保護機制是否與調節(jié)ESR誘導的細胞凋亡有關,本研究采用CSE誘導的A549細胞作為COPD細胞模型,結果發(fā)現,冬蟲夏草100 mg/L可逆轉CSE誘導的A549細胞內質網應激相關蛋白p-eIF2α、p-PERK的表達上調,抑制了內質網應激相關信號通路PERK-eIF2α的激活。
研究還發(fā)現,與空白對照組相比,CSE組促凋亡蛋白BAX明顯增加,抗凋亡蛋白Bcl-2降低,Bax/Bcl-2比值升高,流式細胞和Hoechst 33342染色顯示凋亡率增加,表明香煙引起肺泡上皮細胞內質網應激信號通路PERK-eIF2α活化,并進一步誘導了肺泡上皮細胞凋亡。與CSE組相比,蟲草組Bax/Bcl-2比值降低,流式細胞和Hoechst 33342染色顯示凋亡率下降,提示冬蟲夏草組可通過PERK-eIF2α通路,下調Bax/Bcl-2比值,減輕A549細胞凋亡。
綜上,本實驗初步證明冬蟲夏草通過抑制過度ERS,發(fā)揮抗人肺泡上皮細胞凋亡作用,其作用機制涉及PERK-eIF2α信號通路,表現為冬蟲夏草可以降低p-eIF2α、p-PERK的表達,從而減輕ERS凋亡信號通路激活,減少細胞凋亡。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
