早老素在日本血吸蟲生殖發育中的作用
2021-11-09 11:59:24楊旭晗沈際佳
安徽醫科大學學報 2021年10期
楊旭晗,楊 菲,沈際佳,劉 淼
血吸蟲病是熱帶地區高發的寄生蟲病之一,蟲卵是其傳播和致病最重要的因素。血吸蟲雌蟲性器官的發育成熟和產卵必須通過雌雄蟲合抱方可完成。可能原因為雌雄合抱在蟲體內啟動了特殊基因的表達,這些基因可能參與了調節血吸蟲雌蟲的性成熟和產卵過程。但是,雌雄合抱對雌性生殖系統發育的影響及分子機制尚未完全確定。
早老素 (presenilin, PS) 在多種動物體內均可表達,具有多種生物學作用。但其在血吸蟲蟲體生殖發育中的作用尚未有報道。該研究首先運用熒光定量PCR技術分析早老素基因在血吸蟲發育各個階段的表達情況,隨后通過RNA干擾(RNA interference,RNAi)成蟲的早老素基因,初步證實早老素在日本血吸蟲生殖系統發育中的作用。
1 材料與方法
1.1 實驗動物
雌性6周齡昆明鼠由安徽省實驗動物中心提供,陽性釘螺(含日本血吸蟲尾蚴)購自湖南省血吸蟲病防治研究所。1.2 主要試劑
Tween-20及DEPC購自美國Solarbio 公司; TRIzol購自美國Invitrogen 公司; ProteinaseK、LipofectamineRNAiMAX Reagent、熒光定量PCR試劑盒(SYBR Green)、10×PCR buffer、dNTPmix及逆轉錄試劑盒(PrimeScriptRT reagent Kit)購自日本TaKaRa&Clontech公司;三抗及引物委托上海生工生物公司合成; RPMI1640、無添加劑的M199培養基、OPTI-MEM及胎牛血清(FBS)、糖原購自美國Gibco公司;其余試劑均為國產分析純。1.3 日本血吸蟲雌雄蟲的收集
準備加滿純凈水的10 ml錐形瓶,并將感染日本血吸蟲的釘螺放置于瓶中,于 25 ℃、通風條件下光照4 h,然后收集從釘螺中逸出浮于水面的血吸蟲尾蚴。每只昆明鼠裸露腹部皮膚,給予80只左右的尾蚴經腹部皮膚感染,昆明鼠分別在感染血吸蟲后16、18、24、28 d處死,收集蟲體則使用肝門靜脈灌注法,用PBS充分沖洗蟲體3次,將蟲體上黏附的其他組織清潔干凈,分離雌蟲及雄蟲,并分別儲存在-80 ℃、無RNA酶環境中備用。1.4 日本血吸蟲雌蟲卵巢分離
取50~60條28 d的雌性血吸蟲放入離心管里,用PBS清洗,加入表皮消化劑,在37 ℃水平搖床消化5 min,棄去消化劑,消化4次,然后加入彈性蛋白酶,當雌蟲整個蟲體呈現松弛的團狀時終止,將蟲體倒入平皿中,加入PBS,移液器吹打使卵巢從蟲體釋放。洗滌去卵巢上的附著組織,將卵巢吸取收集,儲存在-80 ℃備用。1.5 日本血吸蟲總RNA的提取
利用TRIzol試劑分別提取感染不同時期的血吸蟲的總RNA。1 mg蟲體加入1 ml TRIzol試劑,使用常規酚氯仿法對總RNA進行抽提。RNA濃度用NanoDrop2000測定并記錄,按照TaKaRa公司的逆轉錄試劑盒說明書的步驟進行cDNA合成。利用Primer Premier5軟件設計熒光定量PCR引物,設計好的引物由上海生工生物公司進行合成。設計合成的引物序列如表1。加樣操作按照熒光定量PCR試劑盒(SYBR Green)的說明進行,而后PCR過程按95 ℃ 10 s,95 ℃ 15 s、60 ℃ 40 s(40 個循環),95 ℃ 15 s、60 ℃ 1 min、95 ℃ 15 s完成,結果分析通過ABI StepOne software進行。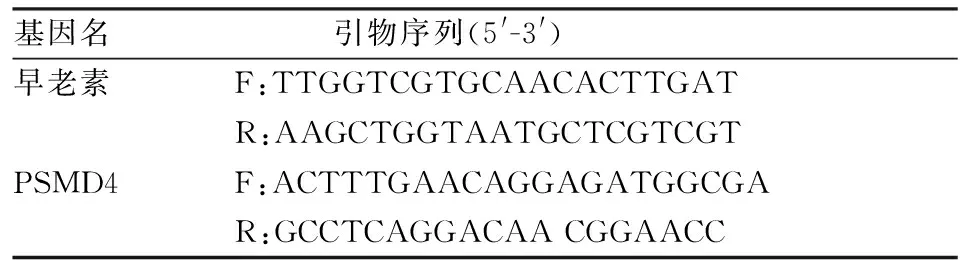
表1 早老素及PSMD4的熒光定量PCR引物序列
1.6 RNA干擾
根據SjPS的mRNA,利用Thermo Fisher在線軟件(https://rnaidesigner.thermofisher.com)設計小干擾RNA(siRNAs)的序列,送至上海吉瑪基因公司合成,并合成1管小干擾RNA為陰性對照,序列見表2。首先處死感染血吸蟲尾蚴28 d后的昆明鼠,利用肝門靜脈灌注法取出血吸蟲蟲體并篩選出合抱的血吸蟲至完全巴赫培養基培養,首先使用3個不同的siRNA對體外培養的成蟲進行RNAi,同時設置DEPC水組(Control)、非特異性siRNA組(negative control, siNCTRL)。根據LipofectamineRNAiMAX Reagent說明書上的操作步驟轉染siRNAs,3 d后收集培養的血吸蟲蟲體并提取總RNA,各組的干擾效率通過熒光定量PCR技術計算并分析。使用同樣條件進行3次平行重復實驗后,確定干擾效率最高的SjPS siRNA1。打亂所選取的小干擾RNA序列,使用NCBI中的BLAST比對證實其無特異性后,送至上海吉瑪基因公司,合成錯配的siRNA,序列見表1。分別設置DEPC水組(Control)、SjPS SiRNA1組與siRNA1錯配組(Mock),并分別于體外作用成蟲10 d,每天進行半量換液,培養10 d后于顯微鏡下計算合抱與未合抱的蟲體數量和蟲卵數量,后收集所有培養的蟲體,一半提取總RNA,使用熒光定量PCR技術計算出siRNA的干擾效率,另一半蟲體使用多聚甲醛固定后,利用中性樹膠封片,使用激光共聚焦顯微鏡觀察干擾后血吸蟲蟲體內生殖系統的形態學變化。
表2 siRNA的序列
1.7 激光共聚焦觀測日本血吸蟲生殖系統
從培養基中挑出培養的血吸蟲完整蟲體,并使用PBS清洗,將雌蟲與雄蟲分離后,分別置入多聚甲醛固定液中固定,使用鹽酸卡紅將固定好的蟲體染色24 h,再脫色,待蟲體呈現透明狀態后,再轉移到載玻片上,使用中性樹膠進行封片。在反射模式下,將激光共聚焦掃描顯微鏡設置為470 nm波長的濾波器以及488 nm波長的He/Ne激光器,觀察日本血吸蟲兩性生殖系統結構形態是否發生變化。1.8 統計學處理
統計分析使用GraphPad Prism軟件(6.0版本)進行。所有試驗全部獨立重復 3 次及以上,組間比較采用單因素方差分析,事后兩兩樣本之間多重檢驗采用Tukey法,將各基因的相對表達量、蟲卵量以及雌雄合抱對數表示為均數±SEM,P
<0.05為差異有統計學意義。2 結果
2.1 早老素在合抱前期、初期及后期的血吸蟲雌蟲表達情況
應用熒光定量PCR技術分析比較早老素在血吸蟲感染16、18、24 d的雌性日本血吸蟲蟲體內的表達情況。選擇PSMD4為管家基因,標準化反應,運用2方法分析早老素mRNA的相對表達量,結果見圖1,從結果分析可知合抱后期的雌蟲體內早老素相對表達量(89.26±6.31)高于其他兩組,差異有統計學意義(F
=196.0,P
<0.001)。
圖1 早老素在合抱前期、初期及后期的雌蟲表達水平與16 d比較:***P<0.001;與18 d比較:###P<0.001
2.2 早老素在血吸蟲雌性成蟲、雄性成蟲及雌性成蟲卵巢的表達情況
應用熒光定量PCR技術分析比較早老素mRNA在血吸蟲雌性成蟲、雄性成蟲及雌性成蟲卵巢的表達水平。結果發現,早老素蛋白在雌性血吸蟲的卵巢中相對表達量(3.00±0.19)高于雌性及雄性血吸蟲蟲體,并且與其他兩組的差異有統計學意義(F
=118.4,P
<0.001)。而雌性血吸蟲蟲體相對表達量(1.04±0.05)及雄性血吸蟲蟲體相對表達量(0.76±0.04)之間差異無統計學意義,見圖2。
圖2 早老素在雌性血吸蟲成蟲蟲體、雄性血吸蟲成蟲蟲體及雌性血吸蟲成蟲卵巢中的表達水平與雌性成蟲蟲體比較:***P<0.001;與雄性成蟲蟲體比較:###P<0.001
2.3 利用siRNA干擾血吸蟲成蟲早老素基因
利用siRNA干擾技術,對體外培養的4周齡血吸蟲成蟲的早老素基因進行干擾。首先,使用針對早老素表達設計的3個不同的siRNA對體外培養的血吸蟲成蟲進行干擾,干擾3 d后收集蟲體,利用熒光定量PCR技術計算出各組早老素的表達水平,從結果分析中能夠發現,在設計的3個siRNA中,SjPS siRNA1干擾后早老素的相對表達水平(0.34±0.028)遠低于其他兩組,差異有統計學意義(F
=45.18,P
<0.001),見圖3,故選擇干擾效率較高的SjPS siRNA1進行下一步實驗。然后利用SjPS siRNA1對體外培養28 d成蟲的早老素基因進行干擾,于10 d后收集蟲體,同時提取總RNA并逆轉錄后,通過熒光定量PCR技術,分析結果發現SjPS siRNA1組的效率與之前篩選實驗的結果一致; 同時發現SjPS siRNA1組的雌雄合抱數(10.40±0.51)低于Control組(12.80±0.73)和Mock組(12.80±0.66),差異有統計學意義(F
=4.645,P
<0.05),見圖3;蟲卵計數的結果顯示SjPS siRNA1組的產卵量(1 308.0±75.47)比Control組(2 872.0±101.7)和Mock組(3 043.0±149.7)均下降(F
=71.30,P
<0.001),見圖4。
圖3 體外篩選干擾效果最好的SjPS siRNAi1:Control組;2:siNCTRL組;3:SjPS siRNA1組;4: SjPS siRNA2組;5: SjPS siRNA3組; 與Control組比較:***P<0.001;與siNCTRL組比較:###P<0.001

圖4 SjPS siRNAi1體外干擾對血吸蟲產卵量及雌雄合抱的影響A:干擾后早老素基因相對表達水平;B:干擾后雌性成蟲產卵數量;C:干擾后雌雄成蟲合抱對數;與Control組比較:*P<0.05,***P<0.001;與Mock組比較:#P<0.05,###P<0.001
2.4 RNAi對日本血吸蟲生殖系統的影響
形態學分析顯示:經過siRNA干擾后的日本血吸蟲雌蟲與雄蟲的生殖系統形態均發生異常。Control組和Mock組均顯示雄性生殖系統由數個睪丸組成,且睪丸之間銜接平滑,同時睪丸內含有多個不同發育階段的雄性生殖細胞,并且比較飽滿,形態規則,呈圓形,包括精原細胞和精母細胞。同時睪丸周圍的基質結構緊湊。然而,SjPS siRNA1組的血吸蟲睪丸形態發生了明顯的變化。睪丸的尺寸變小,睪丸間的銜接處出現褶皺,睪丸內精原細胞和精母細胞的數量開始逐漸減少,且形態不規則,細胞間出現較明顯的間隙。除此之外,睪丸周圍的基質也出現較多的空洞結構。在Control組和Mock組雌蟲中,卵巢后部有大的成熟卵母細胞,前部有小的未成熟的卵母細胞。然而,SjPS siRNA1組血吸蟲雌蟲卵巢內成熟和未成熟卵母細胞界限被打破,卵母細胞分布雜亂,且數量減少,甚至在卵巢內部出現了空洞與裂隙。見圖5。
圖5 日本血吸蟲SjPS被特異性敲降后卵巢及睪丸的形態學變化A~C:雌蟲的卵巢;D~F:雄蟲的睪丸;A、D:Control組;B、E:Mock組;C、F:SjPS siRNA1組;OV:卵巢;TE:睪丸
3 討論
在我國主要流行的是日本血吸蟲病,其治療在很長一段時間內只能依靠吡喹酮藥物,而長期使用吡喹酮這種單一的藥物存在化療藥物抗性的風險。血吸蟲為雌雄異體。雌雄合抱、處于繁殖期的日本血吸蟲能夠持續不斷地產生大量的蟲卵。蟲卵沉積在肝臟和腸系膜靜脈,后者經腸黏膜損傷而排出體外。沉積在臟器的蟲卵能夠形成蟲卵肉芽腫,導致纖維化。
昆明鼠感染后16 d,蟲體發育成合抱前期,18 d為合抱初期,24 d為合抱后期。在前期實驗中,利用抑制性消減雜交技術篩選并鑒定日本血吸蟲雌蟲在合抱前期、合抱初期和合抱后期的差異表達基因,發現早老素在合抱后期表達量顯著升高,推測早老素可能在血吸蟲生殖發育中起到一定作用,值得進一步研究。
早老素在種屬之間高度保守,可分為早老素-1和早老素-2兩種異構體,但早老素在血吸蟲中沒有異構體。早老素主要通過構成γ-分泌酶在高等哺乳動物體內參與多種有關生殖發育的信號通路,包括與生殖腺卵泡的發育、排卵及黃體化等有關的Wnt通路,以及與胚胎發生和卵母細胞發生在內的多種發育過程有關的Notch通路。有研究表明,針對體外培養的曼氏血吸蟲,使用γ-分泌酶制劑DAPT處理后,可以觀察到其蟲卵發育被抑制,產卵量減少。
本研究證實早老素在血吸蟲合抱后期表達量急劇升高,說明雌雄合抱啟動了該基因的表達,同時證實該基因在卵巢高表達,提示早老素在血吸蟲的生殖發育中,尤其在雌蟲的生殖發育中起到重要作用。通過對早老素基因的干擾,早老素被敲降后,抑制了血吸蟲的雌雄合抱和產卵量,說明早老素對維持日本血吸蟲雌雄合抱和產卵起到重要的調控作用,為進一步研究其調控機制奠定了基礎。本研究顯示,利用siRNA干擾過日本血吸蟲早老素基因表達后,從中挑取培養10 d后仍然合抱的血吸蟲成蟲,將雌雄蟲分離后分別用卡紅染色,制作成玻片標本,然后在激光共聚焦顯微鏡下觀察發現,在雄蟲中,睪丸內精原細胞和精母細胞的數量顯著減少,睪丸的小葉明顯小于對照。在雌蟲中,卵巢內成熟和未成熟卵母細胞界限被打破,且數量減少。這些結果表明,早老素被特異性抑制后,導致雄性和雌性的生殖細胞分化與增殖受到抑制,而這些細胞的分化與增殖對于成熟生殖細胞的起始和連續產生具有至關重要的作用,在血吸蟲體內,若成熟生殖細胞減少,那么卵子的數量也將不可避免地減少,從而使蟲卵的發育和產生得到抑制。本研究同時證實RNA干擾組生殖系統相比于空白組雌蟲有明顯的形態學改變,多數早老素被敲降后的血吸蟲睪丸和卵巢中出現空洞與裂隙等異常結構,提示日本血吸蟲的生殖系統發育和分化受到了影響,從而抑制生殖細胞的增殖。本研究顯示,早老素受到抑制后雌蟲的產卵量也明顯下降。接下來將進一步研究以早老素為中心的調節生殖系統發育的信號傳導途徑,將有助于更好地了解日本血吸蟲的生殖生物學,發現調控日本血吸蟲生殖發育信號通路中的重要分子,為尋找可以抑制血吸蟲生殖發育的關鍵靶點奠定基礎。
