馬鈴薯間作玉米對(duì)土壤細(xì)菌群落的影響研究
2021-11-11 06:44:06李光文崔慧珍伏云珍
農(nóng)業(yè)科學(xué)研究 2021年3期
關(guān)鍵詞:差異
李光文,李 倩,崔慧珍,伏云珍,馬 琨
(1.寧夏大學(xué) 農(nóng)學(xué)院,寧夏 銀川 750021;2.寧夏大學(xué)西北土地退化與生態(tài)恢復(fù)國(guó)家重點(diǎn)實(shí)驗(yàn)室培育基地,寧夏 銀川 750021)
間作是一種有效的土地利用方式,被廣泛應(yīng)用于農(nóng)業(yè)生產(chǎn)中[1]。它能通過(guò)作物種內(nèi)、種間的競(jìng)爭(zhēng)與互利關(guān)系,促進(jìn)田間植物群落多樣化,在實(shí)際生產(chǎn)中往往比單作更易獲取光、水和養(yǎng)分資源[2-3]。間作下土壤微生物能通過(guò)參與碳、氮循環(huán)來(lái)影響農(nóng)業(yè)生產(chǎn)系統(tǒng)的可持續(xù)性,通過(guò)改變土壤養(yǎng)分有效性來(lái)影響植物的生長(zhǎng)[4]。因此,植物、土壤和微生物之間的相互作用被認(rèn)為是農(nóng)業(yè)生態(tài)系統(tǒng)的主要驅(qū)動(dòng)力[5]。
目前,有關(guān)間、套作對(duì)土壤微生物影響的研究多集中在豆類、禾谷類作物之間。Bossio等研究認(rèn)為,土壤細(xì)菌群落的多樣性在作物生長(zhǎng)和產(chǎn)量形成中具有重要作用[6]。研究證實(shí),豆類-禾谷類間作、燕麥-紫云英間作可改變土壤細(xì)菌群落結(jié)構(gòu),提高細(xì)菌群落多樣性[7-8];影響土壤革蘭氏陽(yáng)性菌(G+)與革蘭氏陰性菌(G-)的比值[9];增加土壤短芽孢桿菌屬、芽孢桿菌屬、土芽孢桿菌屬和喜鹽芽孢桿菌屬等有益菌屬所占比例,降低鏈球菌屬、奈瑟菌屬、梭菌屬和支原體屬等潛在致病菌屬的比例[10]。Zhang等[11]發(fā)現(xiàn),玉米間作蠶豆明顯改變了土壤細(xì)菌群落結(jié)構(gòu),但沒(méi)有影響土壤細(xì)菌的多樣性。Taschen等[12]發(fā)現(xiàn),小麥間作豌豆誘導(dǎo)的作物種間協(xié)同作用能通過(guò)減少土壤α-變形菌門(mén)(Proteobacteria)和酸桿菌門(mén)(Acidobacteria)的表達(dá),增加放線菌門(mén)(Actinobacteria)的表達(dá),改變土壤細(xì)菌的群落結(jié)構(gòu),并認(rèn)為,土壤細(xì)菌群落結(jié)構(gòu)和多樣性變化是由單作或間作根際分泌物的數(shù)量、性質(zhì)所驅(qū)動(dòng)[12]。也有研究指出,玉米間作馬鈴薯[13-14]、馬鈴薯間作蠶豆和馬鈴薯間作蕎麥[15]也會(huì)影響土壤細(xì)菌的分布,引起土壤細(xì)菌群落組成的變化,在一定程度上能夠提高細(xì)菌多樣性,改變土壤微生物的群落結(jié)構(gòu)。土壤微生物對(duì)間作作物根系分泌物的選擇作用,是導(dǎo)致土壤微生物群落結(jié)構(gòu)存在差異的主要原因[16-17]。可見(jiàn),間作作物種類及種間關(guān)系差異會(huì)導(dǎo)致不同的研究結(jié)果。
間、套作對(duì)土壤細(xì)菌群落影響的研究雖然已有許多報(bào)道[7-8,10,12-17],但由于土壤類型、作物種類、種間關(guān)系等差異,有關(guān)馬鈴薯間作玉米對(duì)土壤細(xì)菌群落多樣性的影響仍然缺乏[9-10]。本研究以連續(xù)4年馬鈴薯、玉米單作及間作的農(nóng)田土壤為對(duì)象,利用TRFLP和磷脂脂肪酸的方法,分析持續(xù)馬鈴薯、玉米單作及間作下的土壤細(xì)菌群落組成、多樣性以及土壤主要微生物群落組成的變化規(guī)律,以期為合理間、套作栽培及作物選擇提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)地概況
試驗(yàn)地位于寧夏隆德縣沙塘鎮(zhèn)和平村,(106°0′37″E,35°35′11″N),海拔1 883 m,年均降水量502 mm,年均蒸發(fā)量1 370 mm。土壤類型為黑壚土,成土母質(zhì)為黃土。供試土壤基礎(chǔ)理化性狀為:全氮1.65 g/kg,全磷1.35 g/kg,有機(jī)質(zhì)33.28 g/kg,堿解氮70.00 mg/kg,速效磷49.24 mg/kg,速效鉀380.00 mg/kg,pH值為8.37(水土比5∶1)。
1.2 試驗(yàn)設(shè)計(jì)
試驗(yàn)開(kāi)始于2014年4月,前茬作物為馬鈴薯。2014—2017年,采用單因素隨機(jī)區(qū)組設(shè)計(jì),利用田間定位栽培,設(shè)3個(gè)處理,4次重復(fù):馬鈴薯單作(P)、玉米單作(M)和馬鈴薯間作玉米(M||P),小區(qū)面積為4 m×4 m=16 m2。每小區(qū)種4幅,幅寬為1 m,玉米采用寬窄行覆膜種植,寬行60 cm、窄行40 cm、株距35 cm,每幅種3行。間作馬鈴薯采用平作種植,行距50 cm、株距40 cm,馬鈴薯與玉米間的行距為25 cm。間作下的馬鈴薯、玉米各種2幅,條帶比為2∶3。單作馬鈴薯每小區(qū)種4幅,每幅種兩行,行距50 cm、株距40 cm。供試玉米品種為長(zhǎng)城706,馬鈴薯為青薯9號(hào)。基施尿素98.44 kg/hm2,磷酸氫二銨325 kg/hm2,全生育期不再追肥。
1.3 樣品采集及處理
2015年和2017年,均采用5點(diǎn)取樣法,在作物收獲前采集作物行間0~20 cm土層土壤樣品,間作處理采集馬鈴薯與玉米相鄰行間的土壤。本試驗(yàn)共采集12個(gè)土壤樣本,各樣本獨(dú)立,低溫保存帶回實(shí)驗(yàn)室后過(guò)1 mm篩,將過(guò)篩后的土壤樣本各分為2份,一份保存在-40℃冰箱,用于土壤微生物群落結(jié)構(gòu)分析;一份保存在-80℃冰箱,用于土壤樣品的長(zhǎng)期保存。2017年的一部分土壤樣本用于土壤細(xì)菌群落多樣性分析;一部分土樣自然風(fēng)干,用于土壤理化性狀分析。
1.4 測(cè)試項(xiàng)目與方法
1.4.1 土壤理化性質(zhì)的測(cè)定 土壤全氮(TN)采用半微量凱氏法;有機(jī)質(zhì)(OM)采用重鉻酸鉀氧化外加熱法;全磷采用HClO4-H2SO4消煮,鉬銻抗比色法;堿解氮采用堿解擴(kuò)散法;速效磷采用NaHCO3浸提-鉬銻抗比色法;速效鉀采用NH4OAc浸提-火焰光度法;土壤pH采用酸度計(jì)測(cè)定,(水土比5∶1)[18]。
1.4.2 土壤細(xì)菌群落組成及多樣性測(cè)定 土壤微生物脂肪酸的測(cè)定采用PLFAs分析,通過(guò)Agilent 7860型氣相色譜儀,MIDI Sherlock脂肪酸圖譜微生物鑒定系統(tǒng)分析待測(cè)樣品[19]。樣品提取主要通過(guò)前提黑暗培養(yǎng)、濃縮、固相萃取、酯化等過(guò)程。
土壤細(xì)菌群落結(jié)構(gòu)及多樣性采用T-RFLP的方法。土壤總DNA提取采用OMEGA公司的試劑盒,按照操作說(shuō)明進(jìn)行。細(xì)菌通用引物序列為:前引物8F(5′-AGAGTTTGATCCTGGCTCAG-3′),后引物926R(5′-CCGTCAATTCCTTTRAGTTT-3′)。擴(kuò)增產(chǎn)物經(jīng)SanPrep柱式DNA膠回收試劑盒純化回收,分別用HaeⅢ和MspⅠ兩種限制性內(nèi)切酶(上海生工)進(jìn)行酶切,擴(kuò)增體系、程序以及酶切體系參見(jiàn)秦越等的研究方法[20]。熒光標(biāo)記(FAM)引物由上海生工合成,擴(kuò)增所用的Taq Plus DNA聚合酶為天根生化科技有限公司生產(chǎn),酶切產(chǎn)物低溫避光委托上海生工經(jīng)毛細(xì)管電泳分離,在ABI自動(dòng)測(cè)序分析儀上記錄樣品片段信息,比較細(xì)菌群落差異。
采用香農(nóng)-維納(Shannon-Weiner)指數(shù)(H)和均勻度指數(shù)(E)描述土壤細(xì)菌群落結(jié)構(gòu)多樣性,計(jì)算公式參見(jiàn)Dunbar J的方法[21]。
H0=-∑pi lnpi,其中,pi為限制性內(nèi)切酶酶切的特定長(zhǎng)度末端片段相對(duì)峰面積(i峰的峰面積/所有峰面積的總和);群落的均勻度E=H/Hmax,其中Hmax=log2(S)表示對(duì)應(yīng)群落中所有分類群都同樣豐富時(shí)的群落多樣性。
1.5 數(shù)據(jù)處理
選擇熒光強(qiáng)度大于50個(gè)相對(duì)熒光單位(RFU)且長(zhǎng)度在50~500 bp之間的末端片段(T-RFs)作為有效片段進(jìn)行后期數(shù)據(jù)處理[22];采用Biodap軟件計(jì)算土壤細(xì)菌群落的Shannon-Weiner指數(shù)及Evenness指數(shù)。為減少實(shí)驗(yàn)誤差,T-RFLP方法測(cè)定的數(shù)據(jù)均為HaeⅢ和MspⅠ兩種內(nèi)切酶的均值;DPS 7.05軟件進(jìn)行單因素方差分析的LSD法多重比較(P<0.05),Canoco 5.0軟件進(jìn)行RDA分析。
2 結(jié)果與分析
2.1 馬鈴薯玉米單作及間作栽培對(duì)土壤細(xì)菌群落的影響
由圖1可見(jiàn),2015年收獲期,持續(xù)馬鈴薯單作、玉米單作及間作栽培下土壤細(xì)菌、G+和G-脂肪酸含量無(wú)顯著差異,而馬鈴薯單作的G+/G-比值與玉米單作及間作栽培間差異顯著(P<0.05)。2017年收獲期,除對(duì)應(yīng)土壤G-脂肪酸含量在玉米單作和馬鈴薯單作間有顯著差異外(P<0.05),土壤細(xì)菌、G+和G+/G-在不同處理間均無(wú)顯著差異。試驗(yàn)中,隨單作及間作栽培時(shí)間的延長(zhǎng),2017年度,以PLFAs表征的土壤細(xì)菌、G+和G-生物量較2015年有一定幅度的下降。其中,玉米單作、馬鈴薯單作以及馬鈴薯間作玉米體系下土壤細(xì)菌的脂肪酸含量分別降低了20.14%、39.02%和42.53%;土壤G+的脂肪酸含量分別下降了24.90%、21.60%和32.47%;土壤G-的脂肪酸含量分別下降了16.99%、46.82%和47.14%。2017年較2015年相比,除馬鈴薯單作的土壤細(xì)菌和G-外,其他處理的土壤細(xì)菌和G-脂肪酸含量均有顯著差異(P<0.05),且G-在間作栽培的年度間有極顯著差異(P<0.01)。除間作栽培下G+和G+/G-有顯著差異外(P<0.05),其他處理的年度間G+和G+/G-均無(wú)顯著差異。
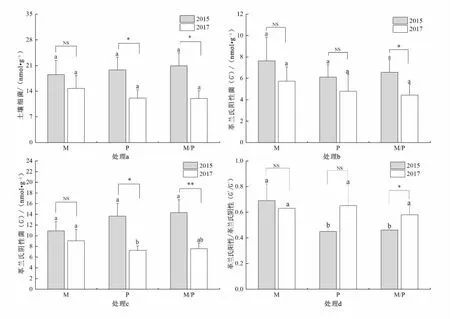
圖1 馬鈴薯、玉米單作及間作栽培對(duì)土壤細(xì)菌群落的影響
總體上,在同一年內(nèi),各處理的土壤細(xì)菌脂肪酸含量波動(dòng)幅度很小,但年際間,馬鈴薯間作玉米栽培模式下的土壤細(xì)菌、G-和G+的脂肪酸含量以及G+/G-比值均有顯著性差異,以PLFAs表征的G-生物量變化對(duì)栽培模式和時(shí)間的長(zhǎng)短比G+更敏感,年際間的差異較年度內(nèi)的變化更明顯。
2.2 間作栽培對(duì)土壤細(xì)菌群落多樣性的影響
由圖2可見(jiàn),與單作馬鈴薯、單作玉米相比,連續(xù)馬鈴薯玉米間作4年后,土壤細(xì)菌群落的香農(nóng)-維納指數(shù)和均勻度指數(shù)分別提高了6.25%~8.97%和2%。但馬鈴薯間作玉米及單作栽培模式下的土壤細(xì)菌群落多樣性及均勻度指數(shù)間均無(wú)顯著差異。可見(jiàn),連續(xù)馬鈴薯、玉米單作及間作在短期內(nèi)并沒(méi)有改變土壤細(xì)菌群落的多樣性。
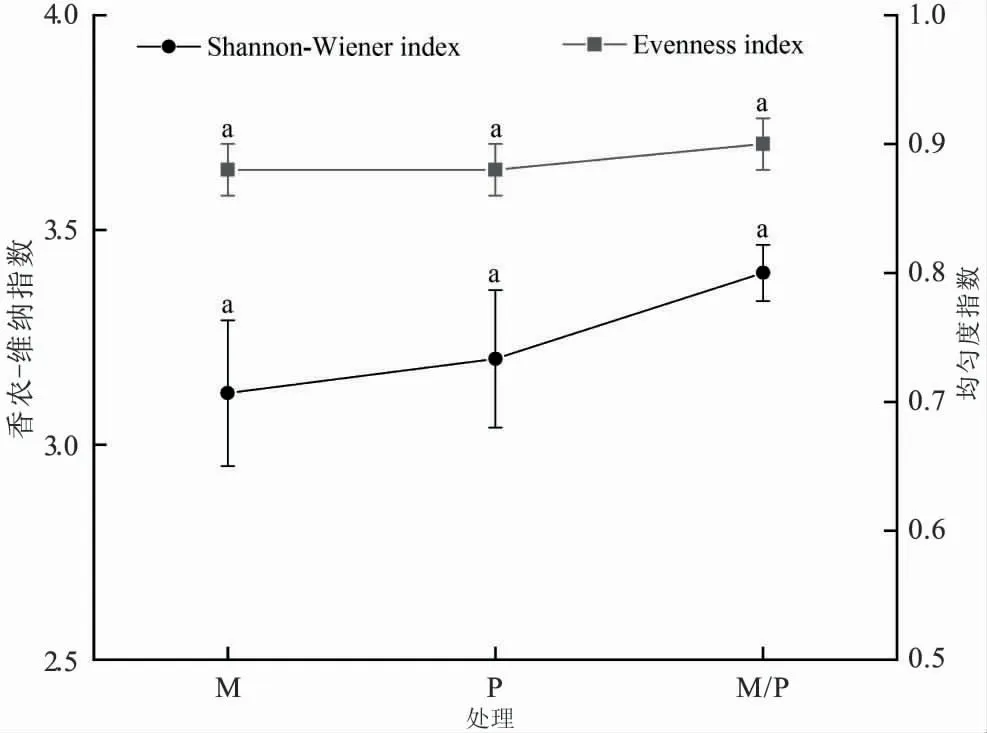
圖2 馬鈴薯、玉米單作及間作栽培對(duì)土壤細(xì)菌多樣性的影響
2.3 馬鈴薯玉米間作及單作栽培對(duì)細(xì)菌菌群結(jié)構(gòu)組成的影響
試驗(yàn)中將T-RFs末端片段與細(xì)菌16S rRNA核糖體數(shù)據(jù)庫(kù)進(jìn)行比對(duì),結(jié)果表明,馬鈴薯單作、玉米單作及間作下所鑒定出的土壤細(xì)菌分屬20門(mén),其中單作玉米土壤細(xì)菌分屬14門(mén),單作馬鈴薯土壤細(xì)菌分屬16個(gè)門(mén),馬鈴薯間作玉米細(xì)菌分屬17門(mén)。相對(duì)豐度百分比較高的前10門(mén)細(xì)菌,土壤細(xì)菌厚壁菌門(mén)(Firmicutes)、變形菌門(mén)(Proteobacteria)和擬桿菌門(mén)(Bacteroidetes)為不同栽培模式下的土壤細(xì)菌優(yōu)勢(shì)菌門(mén)(圖3),且不同處理間均無(wú)顯著差異。連續(xù)玉米單作下,對(duì)應(yīng)土壤厚壁菌門(mén)的相對(duì)豐度百分比達(dá)到37.3%,較馬鈴薯單作和馬鈴薯間作玉米分別高出22.69%和34.17%;土壤變形菌門(mén)的相對(duì)豐度百分比較單作馬鈴薯、馬鈴薯間作玉米也分別高出19.25%和7.63%;擬桿菌門(mén)的相對(duì)豐度則分別高出27.16%和40.4%。然而,整個(gè)土壤細(xì)菌群落中,相對(duì)豐度百分比較低的柔膜菌門(mén)(Tenericutes)、疣微菌門(mén)(Verrucomicrobia)、浮霉菌門(mén)(Planctomycetes)卻表現(xiàn)為馬鈴薯間作玉米均高于單作處理,但各處理間均無(wú)顯著差異。可見(jiàn),不同栽培模式下,土壤細(xì)菌群落組成在門(mén)水平的相對(duì)豐度百分比上存在差異,持續(xù)馬鈴薯間作玉米降低了土壤中厚壁菌門(mén)細(xì)菌的相對(duì)豐度百分比,影響了土壤細(xì)菌門(mén)水平上的群落組成。
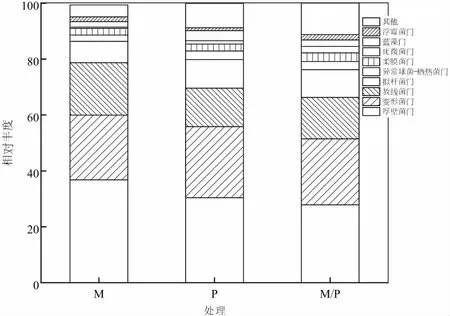
圖3 馬鈴薯、玉米間作栽培影響下的土壤細(xì)菌門(mén)水平分類學(xué)組成及分布
2.4 馬鈴薯間作玉米對(duì)土壤細(xì)菌功能菌群的影響
由表1可見(jiàn),間作模式下參與土壤氮循環(huán)、碳循環(huán)、纖維素降解以及硫循環(huán)過(guò)程的T-RFs所對(duì)應(yīng)功能菌屬的豐度百分?jǐn)?shù),整體高于玉米單作和馬鈴薯單作處理所對(duì)應(yīng)的土壤。長(zhǎng)度為89、159、150 bp的T-RFs片段,歸屬為土壤亞硝化單胞菌屬、節(jié)桿菌屬和固氮螺絲菌屬,均與土壤氮循環(huán)有關(guān),為馬鈴薯間作玉米模式下所特有;不同處理下,長(zhǎng)度為125、128 bp的T-RFs片段所對(duì)應(yīng)的中慢生根瘤菌屬細(xì)菌相對(duì)豐度百分比也以馬鈴薯間作玉米栽培模式最高。與土壤碳循環(huán)有關(guān)的T-RFs片段分別為125、141、142、159、160、169、195、200、203 bp,其對(duì)應(yīng)的土壤細(xì)菌優(yōu)勢(shì)菌群歸類為鏈霉菌屬,其豐度百分比較玉米單作、馬鈴薯單作分別高出2.78倍、1.84倍。此外,參與碳循環(huán),并且歸屬為醋桿菌屬130 bp的TRFs片段和擬桿菌屬93、95、171 bp的T-RFs片段的細(xì)菌相對(duì)豐度百分比也以間作處理較高。
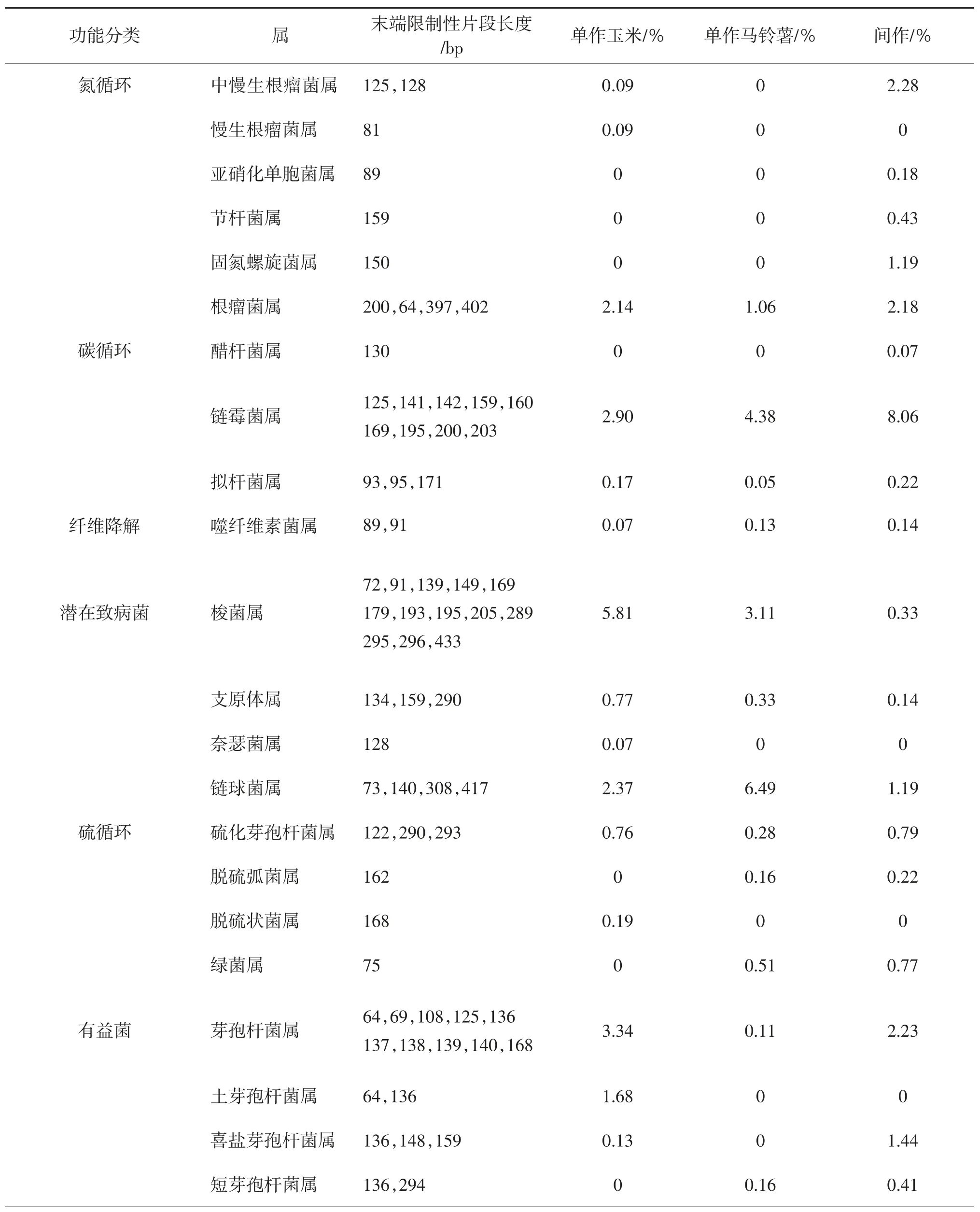
表1 土壤細(xì)菌T-RFs對(duì)應(yīng)的功能菌群相對(duì)豐度百分比
間作模式下,T-RFs片段長(zhǎng)度為200、364、397、402 bp的根瘤菌屬相對(duì)豐度百分比較單作玉米高出了1.87%,是單作馬鈴薯的2.06倍;中慢生根瘤菌屬的豐度百分比為單作玉米的25.33倍,高出單作馬鈴薯2.28%。間作處理中有益菌屬的喜鹽芽孢桿菌屬、短芽孢桿菌屬豐度百分比較單作玉米、單作馬鈴薯也有顯著增加。馬鈴薯間作玉米下土壤潛在致病菌的梭菌屬、支原體屬、鏈球菌屬的豐富百分比均低于單作玉米和單作馬鈴薯;其相對(duì)豐度百分比僅相當(dāng)于對(duì)應(yīng)微生物菌屬的5.68%~10.61%、18.18%~42.42%、18.34%~50.21%;奈瑟菌屬僅出現(xiàn)在單作玉米處理。
可見(jiàn),間作能通過(guò)影響與碳、氮、硫循環(huán)有關(guān)的土壤細(xì)菌群落豐度百分比而影響細(xì)菌群落的組成。間作在一定程度上降低了潛在致病菌所占比例。
2.5 馬鈴薯間作玉米下土壤細(xì)菌群落組成與土壤理化性狀間的相互關(guān)系
RDA分析表明(圖4),全部排序軸能夠在累積變量74.24%上揭示單作及間作體系對(duì)土壤細(xì)菌門(mén)水平分類學(xué)組成差異的影響,其中第一排序軸解釋變量為56.81%,第二排序軸解釋變量為17.43%。厚壁菌門(mén)相對(duì)豐度的改變與土壤全氮呈正相關(guān),而變形菌門(mén)、棲熱菌門(mén)、擬桿菌門(mén)、疣微菌門(mén)相對(duì)豐度的改變與土壤有機(jī)質(zhì)和速效鉀呈正相關(guān),放線菌門(mén)相對(duì)豐度的改變與土壤全氮、全磷呈正相關(guān),與土壤有機(jī)質(zhì)和速效鉀呈負(fù)相關(guān)。但蒙特卡羅檢驗(yàn)表明,馬鈴薯單作、玉米單作及馬鈴薯間作玉米栽培4年后的土壤各理化性狀指標(biāo)與土壤細(xì)菌群落組成相對(duì)豐度間的相關(guān)性均不顯著(P>0.05)。本試驗(yàn)條件下,土壤理化性質(zhì)的差異并不是驅(qū)動(dòng)土壤細(xì)菌群落組成中各主要微生物類群相對(duì)豐度差異的主要原因。

3 討論
3.1 間作栽培對(duì)土壤細(xì)菌主要類群生物量的影響
試驗(yàn)中,馬鈴薯間作玉米下土壤細(xì)菌PLFAs與同一年限馬鈴薯單作、玉米單作均沒(méi)有差異,這與Li[23]對(duì)玉米間作花生的研究結(jié)論一致。在相同栽培年限下,以PLFAs表征的G+生物量在馬鈴薯單作、玉米單作及馬鈴薯間作玉米間無(wú)顯著差異,但G-的脂肪酸含量在單作玉米與單作馬鈴薯間有顯著性差異。Esperschütz等[24]、Buyer等[25]研究表明,相對(duì)G+,G-對(duì)共生營(yíng)養(yǎng)條件較敏感,往往更加依賴根系分泌的碳化合物,能主導(dǎo)枯落物碳的同化[26]。分析認(rèn)為,玉米根系與馬鈴薯根系的分泌物都能為G-提供更豐富的可利用碳源,但單作玉米地上部生物量遠(yuǎn)大于單作馬鈴薯,這可能有助于更多的玉米枯落物進(jìn)入土壤;由于G-優(yōu)先利用輸入土壤系統(tǒng)的新鮮植物枯落物和根系分泌的低分子量有機(jī)碳化合物,而G+被認(rèn)為偏好于較老的以及經(jīng)微生物轉(zhuǎn)化的土壤有機(jī)質(zhì)為碳源[27-28]。因此,這可能就是馬鈴薯單作與玉米單作間,以PLFAs表征的G-生物量有顯著差異的主要原因。
試驗(yàn)中,相比2015年,2017年各處理下以PLFAs表征的細(xì)菌、G+和G-生物量呈現(xiàn)下降趨勢(shì),且間作栽培下降最為顯著,G-較G+脂肪酸含量在年際間的差異較年度內(nèi)的變化更明顯;G+/G-比值除單作玉米外,其他處理在年際間有上升的趨勢(shì),且馬鈴薯間作玉米顯著上升。楊敏等[29]發(fā)現(xiàn),魔芋連作會(huì)使根際土壤中細(xì)菌數(shù)量明顯增多,這與本試驗(yàn)結(jié)果相反;而劉瑩瑩等[30]在黃瓜連作中發(fā)現(xiàn),隨著栽培年限的增加,土壤細(xì)菌、G-細(xì)菌PLFAs量呈降低趨勢(shì),這與本試驗(yàn)結(jié)果一致。可見(jiàn),隨著栽培年限增加,不同作物對(duì)土壤細(xì)菌、G+和G-生物量的影響不同,而間作栽培由兩種作物共同參與,更有益于土壤細(xì)菌、G+和G-生物量的調(diào)節(jié)。土壤中G+/G-比值能反應(yīng)農(nóng)田生態(tài)系統(tǒng)的穩(wěn)定性,G+/G-比值越大,土壤生態(tài)系統(tǒng)越穩(wěn)定[31];G+/G-比值的高低也能表明腐殖化碳源的多少[32]。可見(jiàn),相比2015年,2017年馬鈴薯間作玉米的G+/G-比值顯著上升的主要原因是馬鈴薯間作玉米栽培更有利于土壤生態(tài)系統(tǒng)的穩(wěn)定,并能腐殖化更高的碳源。
3.2 間作栽培對(duì)土壤細(xì)菌群落組成、多樣性及功能的影響
單作馬鈴薯、單作玉米及馬鈴薯間作玉米下土壤厚壁菌門(mén)、變形菌門(mén)和擬桿菌門(mén)為土壤細(xì)菌優(yōu)勢(shì)菌門(mén);馬鈴薯間作玉米表現(xiàn)出較高的相對(duì)豐度,這與Zheng等[33]的研究結(jié)果一致。相關(guān)研究認(rèn)為,間作作物類型不同,改變了菌門(mén)相對(duì)豐度的百分比,卻未改變優(yōu)勢(shì)菌門(mén)的類型,這可能是處于相同生長(zhǎng)環(huán)境中的土壤微生物類群具有相似性[34]。趙雅效等[35]在紫花苜蓿間作小黑麥中發(fā)現(xiàn)土壤細(xì)菌的優(yōu)勢(shì)菌門(mén)是變形菌門(mén)和擬桿菌門(mén),并發(fā)現(xiàn)間作能改變土壤變形菌門(mén)和擬桿菌門(mén)的相對(duì)豐度,提高土壤細(xì)菌豐富度和多樣性,這與Zhang等[36]桑樹(shù)間作紫花苜蓿的研究結(jié)果一致。同時(shí),變形菌門(mén)在土壤細(xì)菌群落中具有較高的相對(duì)豐度[37],并且是堿性土壤的優(yōu)勢(shì)種群[38-39]。本試驗(yàn)中,厚壁菌門(mén)、變形菌門(mén)和擬桿菌門(mén)相對(duì)豐度的改變,可能是種植模式和作物不同導(dǎo)致的[40],其改變了土壤根際微環(huán)境,改善了細(xì)菌菌門(mén)多樣性,使相對(duì)豐度發(fā)生了變化。也可能受土壤類型的影響[41],存在對(duì)土壤微環(huán)境敏感性強(qiáng)的細(xì)菌,能刺激特定菌門(mén)細(xì)菌的生長(zhǎng),并成為優(yōu)勢(shì)菌門(mén),具體原因有待進(jìn)一步研究。
本研究表明,與單作相比,馬鈴薯間作玉米下土壤細(xì)菌群落的多樣性和均勻度指數(shù)間無(wú)差異。這同樊曉剛等[42]土壤微生物群落通常具有比較穩(wěn)定的多樣性特征的研究結(jié)論一致。但Li等[23]、Latati等[43]研究認(rèn)為,間作作物地下交互作用導(dǎo)致根際土壤微生物群落的物種組成具有特異性。實(shí)際上,在間作條件下,植物根系不是完全孤立而是相互交織在一起,通過(guò)作物根系活動(dòng)、分泌物、根際沉積特征的影響以及豆科作物根際土壤特異性細(xì)菌,刺激作物根際的過(guò)程,導(dǎo)致根際化學(xué)特性的變化,是間作下土壤細(xì)菌多樣性產(chǎn)生明顯差異的主要原因。然而,試驗(yàn)中,馬鈴薯間作玉米下土壤微生物群落多樣性與前人研究結(jié)果的差異表明,間作體系對(duì)土壤微生物群落的調(diào)控是一個(gè)緩慢的過(guò)程,在多種因子綜合作用下,隨種植年限延長(zhǎng),間作與單作對(duì)土壤微生物多樣性的影響差異,才有可能逐漸表現(xiàn)出來(lái)。
試驗(yàn)中,間作下參與碳循環(huán)的擬桿菌屬細(xì)菌相對(duì)豐度顯著增加,相關(guān)研究也證實(shí),土壤厚壁菌門(mén)、變形菌門(mén)和擬桿菌門(mén)細(xì)菌與土壤碳的降解有關(guān)[44]。因此,這也間接說(shuō)明土壤擬桿菌屬細(xì)菌是重要的有機(jī)物分解者。間作模式下,馬鈴薯、玉米地下種間作用可能有利于提升土壤細(xì)菌的分解能力。有研究證實(shí),細(xì)菌群落結(jié)構(gòu)的變化顯著影響了由專性微生物介導(dǎo)的固氮和硝化作用等功能過(guò)程[45]。Song等[46]的研究結(jié)果表明:禾谷類作物和豆科作物間作,能改變固氮菌的優(yōu)勢(shì)種,影響土壤微生物的群落組成,進(jìn)而促進(jìn)豆類作物結(jié)瘤固氮。玉米花生間作,較單作顯著增加了與固氮有關(guān)的土壤細(xì)菌的相對(duì)豐度,是因?yàn)橛衩谆ㄉg作減少了與反硝化作用有關(guān)的基因拷貝數(shù),顯著緩解了化學(xué)氮肥對(duì)結(jié)瘤和N2固定的抑制效應(yīng)[17]。本研究中,脫硫弧菌屬相對(duì)豐度的顯著增加,也表明馬鈴薯間作玉米能促進(jìn)土壤硫循環(huán),改善土壤理化性質(zhì)以及土壤硫元素的有效性,這同Muyzer和Stams[47]有關(guān)脫硫桿菌屬(Desulfobacterium)的描述類似。總體上,馬鈴薯間作玉米增加了與土壤碳、氮、硫循環(huán)以及纖維素分解有關(guān)細(xì)菌菌屬的相對(duì)豐度。增加了有益菌,降低了有害菌的豐度。但在禾本科(玉米)與茄科作物(馬鈴薯)的種間作用下為什么能產(chǎn)生與禾本科、豆科間作類似的微生物功能效應(yīng),還需要進(jìn)一步研究。
試驗(yàn)中,土壤厚壁菌門(mén)相對(duì)豐度與全氮呈正相關(guān),土壤變形菌門(mén)、擬桿菌門(mén)相對(duì)豐度與土壤速效鉀、有機(jī)質(zhì)呈正相關(guān)。這是因?yàn)楹癖诰T(mén)、擬桿菌門(mén)細(xì)菌能參與土壤養(yǎng)分的循環(huán),對(duì)土壤全氮、速效鉀和有機(jī)質(zhì)具有一定的消解作用,使其快速轉(zhuǎn)變?yōu)樽魑锷L(zhǎng)所需的營(yíng)養(yǎng)物質(zhì)[48]。此外,厚壁菌門(mén)是在作物根際土壤中存在的一種固氮菌,能很好地參與土壤氮素的轉(zhuǎn)化[49]。變形菌門(mén)屬于異養(yǎng)型細(xì)菌,是與植物共生的固氮細(xì)菌,能增強(qiáng)土壤的固氮能力,其豐富度的高低能在一定程度上反應(yīng)土壤有機(jī)質(zhì)含量的高低[34]。擬桿菌門(mén)是有機(jī)碳的主要礦化者[50],能為土壤細(xì)菌的活動(dòng)提供一定的碳源,這與本研究中變形菌門(mén)、擬桿菌門(mén)與有機(jī)質(zhì)呈正相關(guān)的結(jié)論相吻合。因此,土壤全氮、速效鉀和有機(jī)質(zhì)含量能影響相應(yīng)菌門(mén)相對(duì)豐度的變化。但本研究中,RDA檢驗(yàn)表明,土壤各理化性狀指標(biāo)與土壤細(xì)菌群落組成相對(duì)豐度間的相關(guān)性不顯著,故土壤理化差異不是驅(qū)動(dòng)土壤細(xì)菌群落組成變化的主要原因。可見(jiàn),驅(qū)動(dòng)間作土壤細(xì)菌變化是土壤環(huán)境整體變化的結(jié)果,它可能由種植模式、作物種類、氣候因子、土壤環(huán)境因子以及作物間的種競(jìng)爭(zhēng)互利關(guān)系等因素共同決定。
4 結(jié)論
馬鈴薯玉米間作及單作栽培在短期內(nèi)沒(méi)有影響土壤細(xì)菌群落的多樣性,但改變了土壤細(xì)菌群落組成,不同栽培模式下的土壤細(xì)菌群落相對(duì)豐度差異沒(méi)有受到土壤理化性質(zhì)差異的顯著影響。玉米單作下土壤革蘭氏陰性菌(G-)對(duì)土壤環(huán)境變化的影響比馬鈴薯單作和馬鈴薯間作玉米對(duì)土壤的影響更敏感。馬鈴薯間作玉米提高了土壤中有益細(xì)菌菌屬的豐度百分比、降低了潛在致病菌屬的豐度百分比。
猜你喜歡
英語(yǔ)世界(2023年10期)2023-11-17 09:19:16
汽車實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國(guó)非營(yíng)利評(píng)論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語(yǔ)文(2016年21期)2016-05-25 13:13:50
