草蓯蓉多糖對過氧化氫致HL-7702細胞損傷的保護作用
2021-11-17 09:24:44金愛花常慧張釗葛乃嘉全吉淑尹學哲
食品與發酵工業 2021年21期
金愛花,常慧,張釗,葛乃嘉,全吉淑*,尹學哲*
1(延邊大學附屬醫院,吉林 延吉,133000)2(延邊大學 醫學院,吉林 延吉,133002)
肝損傷是指各種致病因素作用于肝組織而引起的肝細胞變性、壞死以及肝功能改變。活性氧(reactive oxygen species,ROS)自由基在不同形式的肝損傷過程中扮演了重要角色,而ROS引發的氧化應激是肝細胞損傷的重要因素,是包括肝損傷在內的多種肝臟疾病發生的病理基礎。當肝細胞內ROS過剩時,其引發的氧化應激容易使細胞內脂質、核酸、蛋白質等生物分子發生改變,導致肝細胞結構和功能的損傷,最終誘發肝細胞死亡[1-2]。H2O2是一類小分子活性氧,易溶于水和穿透細胞膜,是常見的細胞氧化損傷劑[3];其能夠誘導肝細胞DNA損傷,導致受損基因表達發生改變,同時還可誘發細胞內蛋白質和脂質損傷[4]。當肝細胞損傷較輕時不僅不會造成肝細胞死亡,在抗氧化劑作用下肝細胞的這些損傷還有可能被修復[4-5]。草蓯蓉多糖(Boschniakiarossicapolysaccharides,BRP)是草蓯蓉全草中最為豐富的重要活性成分,無毒,具有較強的抗氧化及免疫調節活性,在多種疾病的防治中發揮重要作用[6-8]。課題組研究發現,BRP對HepG2細胞氧化應激損傷具有抑制作用,且抑制HepG2細胞核轉錄因子-κB(nuclear factor-κB,NF-κB)的激活[9]。HepG2細胞雖具有肝實質細胞的部分特征,但畢竟是肝癌細胞,其生長規律和生物學功能與正常肝細胞可能有些差異[10-11]。因此,有可能不能完整體現正常肝實質細胞受到損傷時的實際情況。HL-7702細胞,又稱L02細胞,是永生化的人正常肝細胞,可連續傳代,并在多次傳代中仍能保持正常肝實質細胞原有生化和生理特征,是體外肝細胞研究的理想細胞系,常用于肝細胞毒性的細胞學研究中[12-13]。
本研究以H2O2為誘導劑,HL-7702人正常肝細胞株為研究對象,建立肝細胞氧化應激損傷模型,分別從增殖、凋亡以及相關信號通路等方面,探討BRP對H2O2誘導的HL-7702細胞損傷的保護作用以及相關作用機制,為其在保肝方面的應用提供科學依據。
1 試驗材料
1.1 細胞株與試劑
BRP,由延邊大學醫學院生物化學與分子生物學教研室制備,多糖含量為97.4%,主要為分子質量為4.9×104Da的中性多糖[6]。HL-7702細胞株,江蘇凱基生物技術股份有限公司;DMEM高糖培養基、胎牛血清,以色列BI公司;胰蛋白酶,美國西格瑪公司;CCK-8試劑盒,北京莊盟國際生物基因科技有限公司;乳酸脫氫酶(lactate dehydrogenase,LDH)測試盒、丙二醛(malondialdelyde,MDA)測試盒,南京建成生物科技有限公司;Hoechst33342染色液、原位末端標記(TdT-mediated dUTP nick end labeling,TUNEL)細胞凋亡測試盒,上海碧云天生物技術有限公司;小鼠p53、多聚ADP核糖聚合酶[poly(ADP-ribose)polymerase,PARP]、細胞外信號調節蛋白激酶(extracellular signal-regulated kinase,ERK)抗體以及兔Bcl-2、Bax、Bak、細胞色素c(cytochrome c,Cyto c)、Caspase-3、c-Jun N末端激酶(c-Jun N-terminal kinase,JNK)、p-JNK、p38絲裂原活化蛋白激酶(p38 mitogen-activated protein kinase,p38)、p-p38、p-ERK、NF-κB抗體,美國賽信通公司;兔β-actin抗體、兔抗小鼠二抗、山羊抗兔二抗,美國西格瑪公司。
1.2 儀器與設備
CO2細胞培養箱,美國Shellab公司;超聲波細胞粉碎機,寧波新芝生物科技有限公司;酶標分析儀,深圳雷杜公司;冷凍離心機,德國艾本德公司;垂直板電泳儀、轉印槽,美國伯樂公司;全自動化學成像分析系統,上海培清科技有限公司。
2 實驗方法
2.1 BRP無菌液的配制
將BRP用無血清的DMEM培養基配成質量濃度為400 mg/L的儲液,過濾除菌,貯于4 ℃備用。
2.2 HL-7702細胞的培養及傳代
HL-7702細胞用DMEM培養基(含10%胎牛血清、100 μg/mL鏈霉素、100 U/mL青霉素)在體積分數5% CO2、飽和濕度條件下常規培養[9,13]。待HL-7702細胞達到80%以上融合度時,用胰酶消化并傳代[9,13]。
2.3 BRP安全劑量的選定
取HL-7702細胞接種于96孔板中,每孔接種密度為2×105個/mL[9,13]。放置24 h待HL-7702細胞貼壁后,換成含BRP培養液,使BRP質量濃度分別為0、25、50、100、200和400 mg/L,繼續培養12 h。用CCK-8測試盒測定HL-7702細胞存活率,以細胞存活率≥95%為BRP安全使用劑量[14]。每組設6個復孔,實驗重復3次。

(1)
2.4 H2O2誘導的HL-7702細胞氧化損傷模型的建立
取HL-7702細胞接種于96孔板中,每孔接種密度為2×105個/mL[9,13]。培養24 h待HL-7702細胞貼壁。換入含H2O2(摩爾濃度分別為75、150、300、600和1 200 μmol/L)的無血清培養液,繼續培養6 h[9,13]。按測試盒操作步驟和方法,用CCK-8法檢測HL-7702細胞存活率。每組設6個復孔,實驗重復3次。
2.5 BRP對H2O2誘導HL-7702細胞氧化損傷的影響
取HL-7702細胞接種于96孔板中,每孔接種密度為2×105個/mL[9,13]。培養24 h待HL-7702細胞貼壁。隨機分為正常組、模型組、BRP低劑量組和高劑量組(分別為BRP50組和BRP100組)。模型組HL-7702細胞先常規培養24 h待細胞貼壁后,于無血清培養液中培養12 h,加H2O2使其終摩爾濃度為300 μmol/L,繼續培養6 h;BRP低劑量組和高劑量組HL-7702細胞常規培養24 h,分別用含50和100 mg/L BRP的無血清培養液繼續培養12 h,再用終摩爾濃度為300 μmol/L的H2O2損傷6 h;正常組則常規培養24 h,用無血清培養液繼續培養12 h,再換無血清培養液培養6 h[9,13]。按測試盒操作步驟和方法,用CCK-8法檢測HL-7702細胞存活率,用微板法檢測細胞及培養液中LDH活性并計算LDH釋放率[13]。每組設6個復孔,實驗重復3次。
(2)
2.6 BRP對H2O2誘導HL-7702細胞MDA水平的影響
取HL-7702細胞接種于6孔板中,分組和處理同2.5。按測試盒操作步驟和方法,用硫代巴比妥酸法檢測細胞中MDA生成量[13,15]。
2.7 BRP對H2O2誘導HL-7702細胞凋亡的影響
細胞凋亡的觀察采用Hoechst33342染色法及TUNEL染色法。取HL-7702細胞接種于6孔板中,分組和處理同2.5。細胞固定,分別加Hoechst33342染色液及TUNEL工作液孵育,封片后在熒光顯微鏡下觀察染色結果[16]。
2.8 BRP對H2O2誘導HL-7702細胞凋亡相關蛋白表達的影響
取HL-7702細胞接種于100 mm培養皿中,分組和處理同2.5。分別提取細胞總蛋白、胞漿及核蛋白,并在SDS-PAGE上進行電泳、分離蛋白質,并轉移到PVDF膜上,封閉,敷一抗、二抗,ECL顯色,采集圖像并進行灰度分析[9,15]。
2.9 數據分析
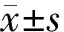
3 結果與分析
3.1 BRP安全使用劑量的確定
為了研究BRP劑量對HL-7702細胞生長的影響,并確定BRP的安全使用劑量,觀察了不同質量濃度BRP處理HL-7702細胞時細胞存活率的變化,結果如圖1所示。當BRP質量濃度在25~100 mg/L時,HL-7702細胞存活率均高于95%,對細胞生長的影響不明顯,可視為無顯著細胞毒性;而當BRP質量濃度在200~400 mg/L時,HL-7702細胞存活率低于95%,具有細胞毒性作用。因此,本實驗選定50和100 mg/L為后續研究中BRP干預的安全劑量。
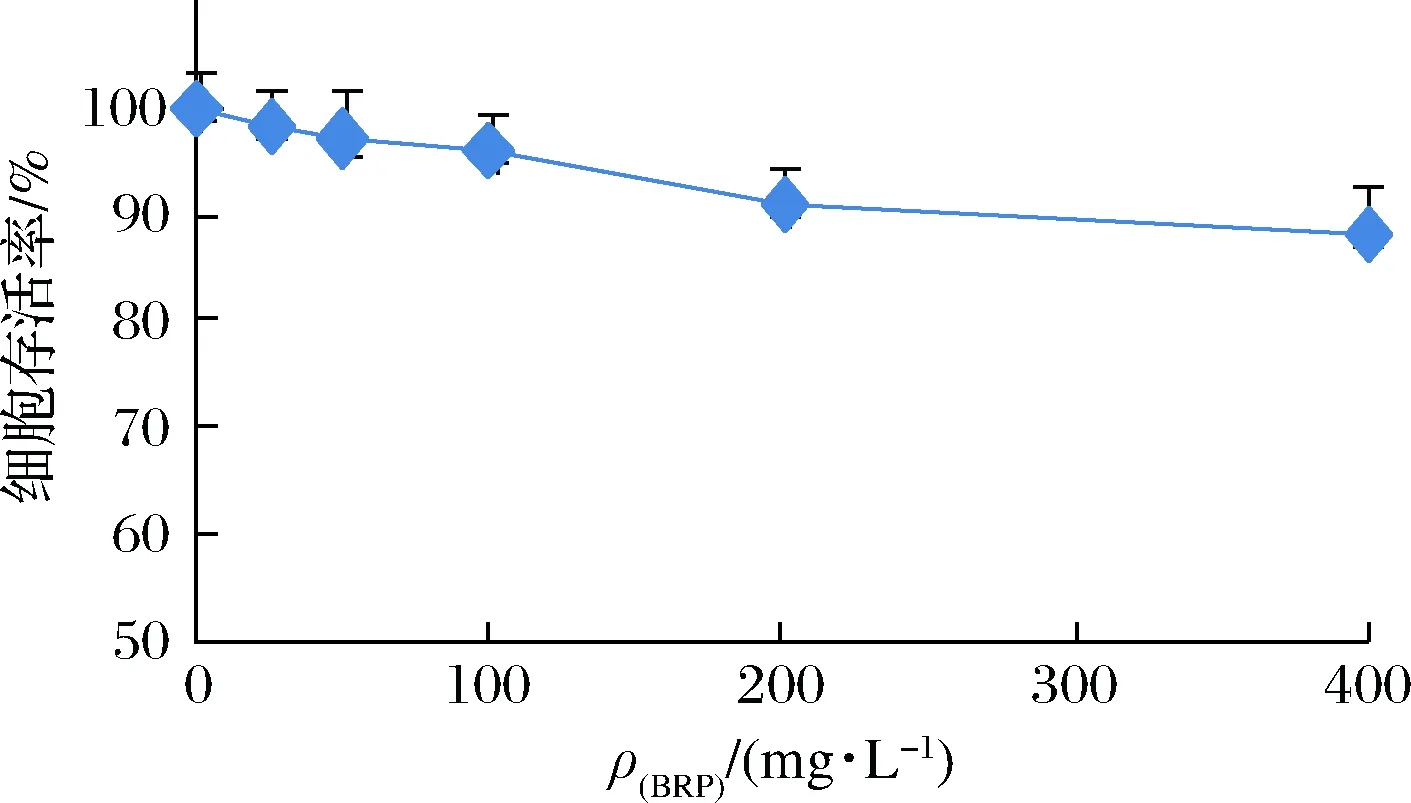
圖1 BRP對HL-7702細胞存活率的影響Fig.1 Effect of BRP on viabilities of HL-7702 cells
3.2 H2O2損傷濃度的確定
H2O2誘導的氧化應激是肝細胞損傷的主要機制之一,可使肝細胞生成過量ROS,造成肝細胞損傷甚至死亡[13,15]。為了研究H2O2抑制HL-7702細胞生長的作用規律,找出誘導HL-7702細胞損傷的最佳使用劑量,觀察了不同質量濃度H2O2處理HL-7702細胞時該細胞存活率的變化,結果如圖2所示。H2O2可顯著降低HL-7702細胞存活率,當H2O2質量濃度為300 μmol/L時,HL-7702細胞存活率約為42%,接近半數抑制率。因此,在后續的研究中,模型制備條件選定為300 μmol/L H2O2處理HL-7702細胞6 h。
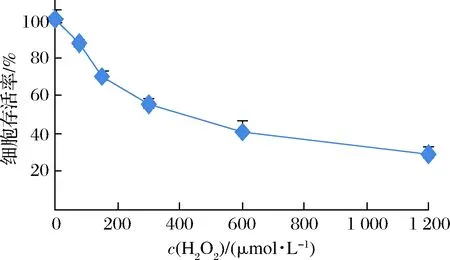
圖2 H2O2對HL-7702細胞存活率的影響Fig.2 Effect of H2O2 concentration on viabilities of HL-7702 cells
3.3 BRP對H2O2誘導的HL-7702細胞存活率影響
BRP對H2O2誘導的HL-7702細胞存活率的影響如圖3所示。BRP能夠提高H2O2損傷的HL-7702細胞存活率(P<0.05),當BRP質量濃度分別為50和100 mg/L時,HL-7702細胞存活率為80.3%和84.9%(P<0.05),比模型組增高26.2%和30.8%。但BRP低劑量組和高劑量組間差異卻無統計學意義(P>0.05)。
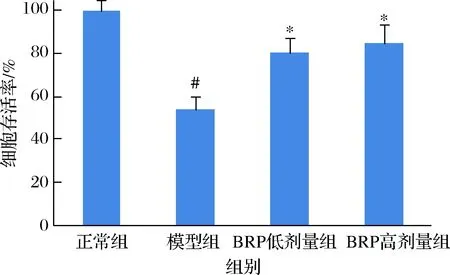
圖3 BRP對H2O2誘導的HL-7702細胞存活率的影響Fig.3 Effect of BRP on cell viabilities of H2O2-induced HL-7702 cells注:與正常組比較,#P<0.05;與模型組比較,*P<0.05(下同)
3.4 BRP對H2O2誘導的HL-7702細胞LDH釋放的影響
細胞損傷或死亡過程中,肝細胞內的重要胞內酶如LDH,可釋放到細胞培養液中,造成培養液中LDH活性增高,因此可以由培養液中的LDH釋放率評價細胞受損或死亡程度[17]。BRP對HL-7702細胞培養液中LDH釋放率的影響如圖4所示。BRP能夠降低H2O2誘導的HL-7702細胞培養液中LDH的釋放(P<0.05),當BRP質量濃度為50和100 mg/L時,培養液中LDH釋放率比模型組分別降低26.9%和32.8%,表明BRP能夠有效保護HL-7702細胞膜完整性。但BRP不同劑量組間的差異卻無統計學意義(P>0.05)。
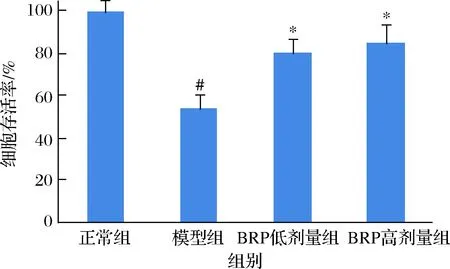
圖4 BRP對H2O2誘導的HL-7702細胞LDH 釋放的影響Fig.4 Effect of BRP on LDH release of H2O2-induced HL-7702 cells
3.5 BRP對H2O2誘導的HL-7702細胞MDA生成的影響
用H2O2損傷肝細胞可使肝細胞內產生大量ROS,并引發細胞膜脂質的過氧化反應,細胞內生成的MDA增多[18]。BRP對H2O2損傷的HL-7702細胞MDA水平的影響如圖5所示。BRP能夠降低H2O2誘導的HL-7702細胞MDA的生成(P<0.05),當BRP質量濃度為50和100 mg/L時,MDA生成量與模型組比較分別降低0.32和0.43 μmol/μg,提示BRP預處理可降低HL-7702細胞氧化應激水平。但BRP不同劑量組間的差異卻無統計學意義(P>0.05)。
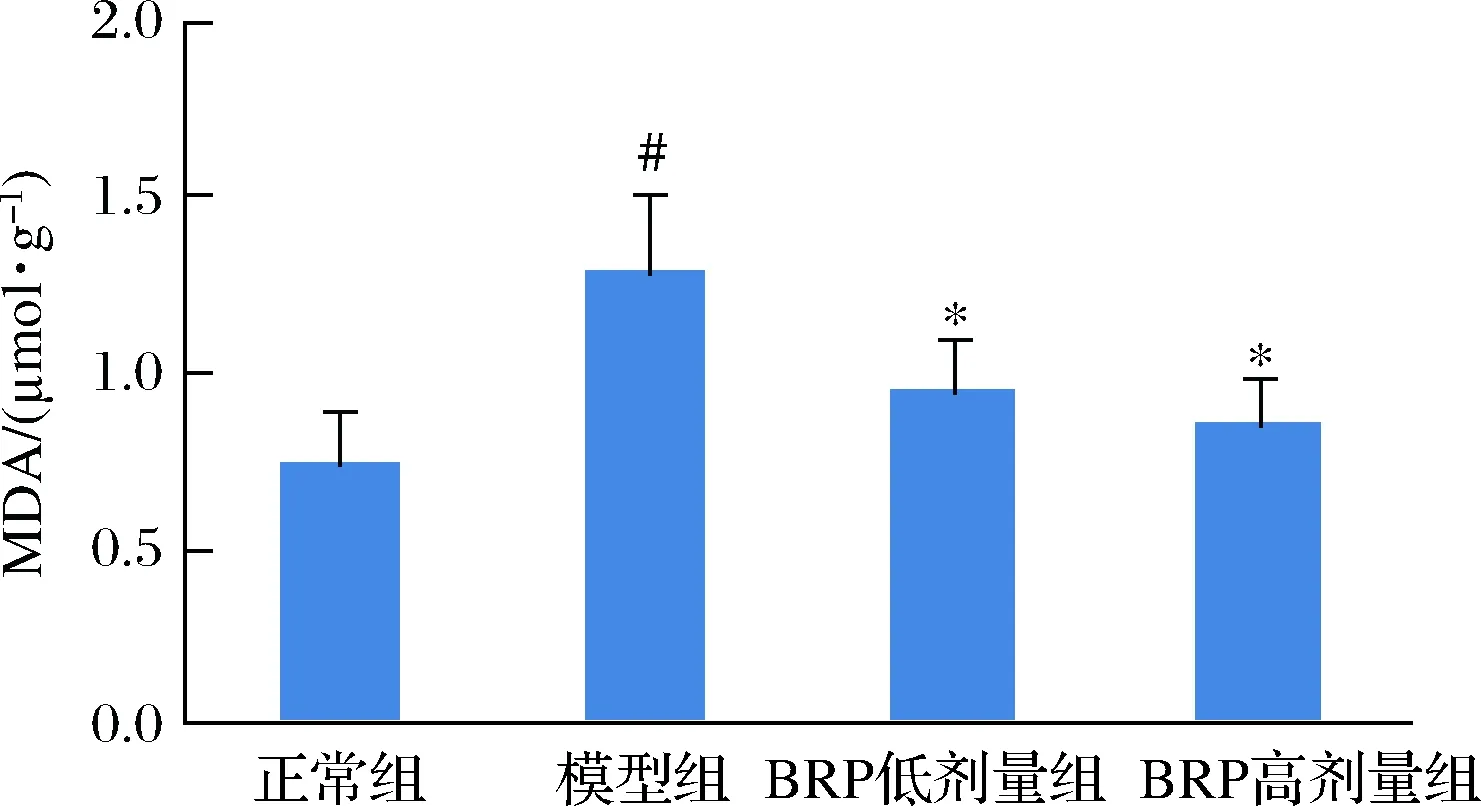
圖5 BRP對H2O2誘導的HL-7702細胞MDA水平的影響Fig.5 Effect of BRP on MDA levels in H2O2-induced HL-7702 cells
3.6 BRP對HL-7702細胞Hoechst33342染色的影響
Hoechst染料是一種可穿過細胞膜的細胞核復染劑,與DNA結合后發出藍色熒光,常用于細胞凋亡檢測,以分辨凋亡細胞中固縮的細胞核[19]。Hoechst33342可使正常細胞核染上較淺的藍色熒光,而細胞發生凋亡時,其熒光強度會變強[20]。BRP對H2O2誘導的HL-7702細胞Hoechst33342染色的影響如圖6所示。正常組HL-7702細胞核染色均勻且較淺;與正常組比較,模型組HL-7702細胞核熒光染色增強,部分細胞核出現核固縮,這與前人報道一致[21];與模型組比較,BRP組HL-7702細胞核熒光染色減弱,凋亡細胞減少。
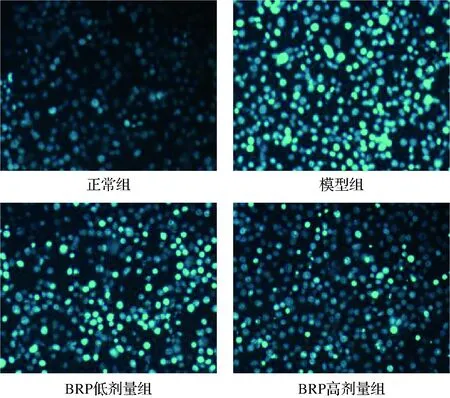
圖6 BRP對HL-7702細胞Hoechst33342染色的 影響(×100)Fig.6 Effect of BRP on Hoechst33342 staining of HL-7702 cells (×100)
3.7 BRP對H2O2誘導的HL-7702細胞凋亡的影響
TUNEL法為細胞凋亡檢測的“金標準”,可使凋亡細胞中斷裂DNA 3′-OH末端的被熒光素標記的脫氧尿苷酸發出綠色熒光[22]。BRP對H2O2誘導的HL-7702細胞凋亡的影響如圖7所示。與正常組比較,模型組發出綠色熒光的凋亡細胞明顯增多;與模型組比較,BRP組凋亡的HL-7702細胞明顯減少,提示BRP預處理可抑制H2O2誘導的HL-7702細胞的凋亡。
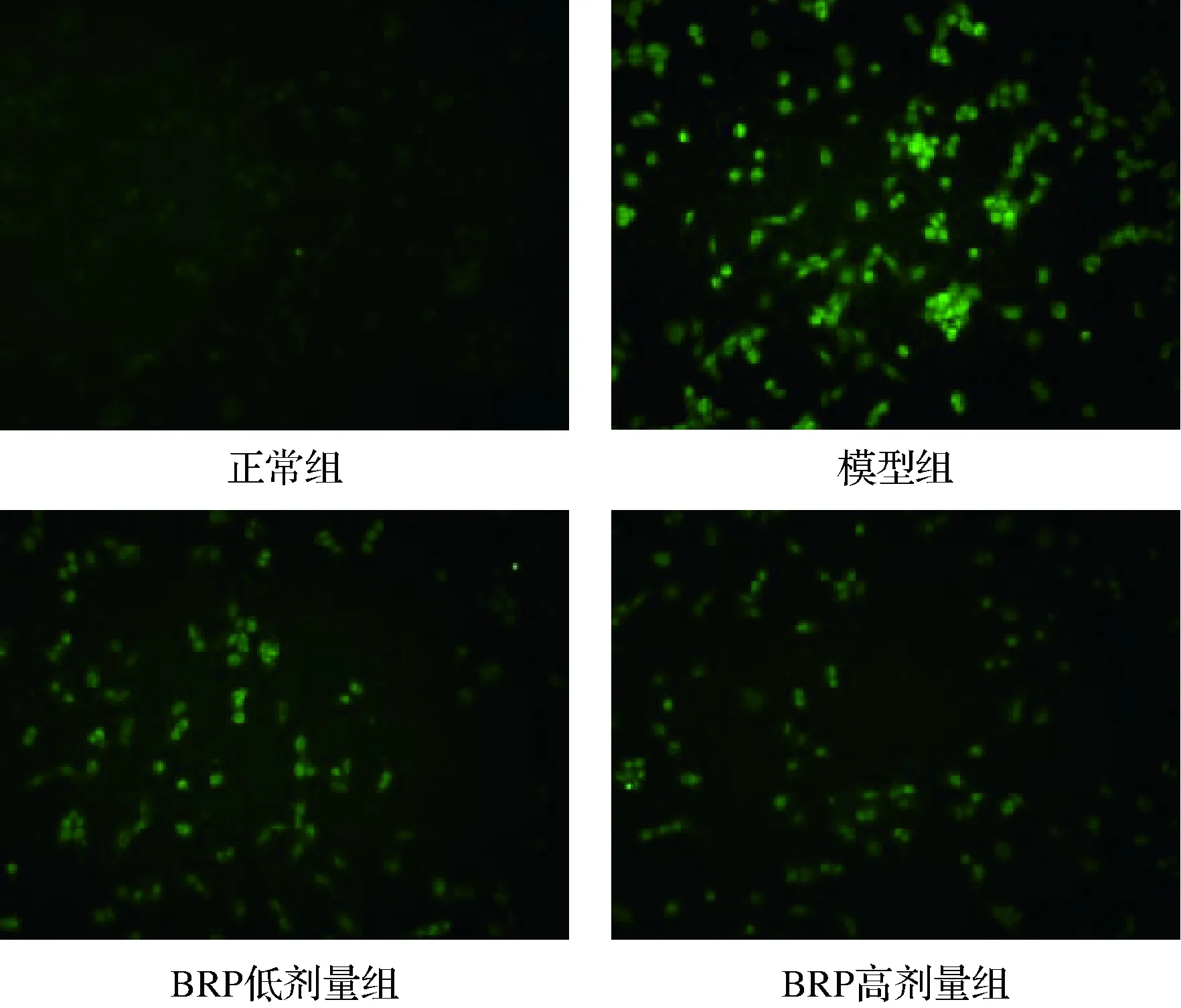
圖7 BRP對H2O2誘導的HL-7702細胞凋亡的 影響(×100)Fig.7 Effect of BRP on apoptosis of H2O2-induced HL-7702 cells (×100)
3.8 BRP對H2O2誘導的HL-7702細胞凋亡相關蛋白表達的影響
不同劑量BRP對損傷HL-7702細胞的保護作用,從細胞毒性、氧化應激以及細胞凋亡等多個指標上均不顯示顯著性差異,但在趨勢上高劑量組比低劑量組有優勢,因此在后續的機制研究中,選100 mg/L為BRP干預劑量。BRP對H2O2誘導HL-7702細胞凋亡相關蛋白表達的影響如圖8所示。與正常組比較,模型組HL-7702細胞漿Cyto c水平升高,細胞p53、Bak、Bax等促凋亡蛋白的表達上調(P<0.05),抗凋亡蛋白Bcl-2表達下調(P<0.05),C-casp3及C-PARP水平升高(P<0.05);與模型組比較,BRP組HL-7702細胞漿Cyto c水平降低(P<0.05),促凋亡蛋白p53、Bak、Bax表達下調(P<0.05),抗凋亡蛋白Bcl-2表達上調(P<0.05),C-casp3和C-PARP水平下降(P<0.05)。提示,BRP可能通過降低促凋亡蛋白和抗凋亡蛋白的比值而抑制H2O2誘導的肝細胞凋亡。
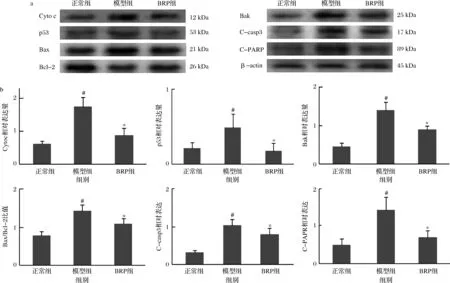
a-蛋白印跡結果;b-灰度比值結果圖8 BRP對HL-7702細胞凋亡相關蛋白表達的影響Fig.8 Effect of BRP on expression of apoptosis-related proteins in HL-7702 cells
3.9 BRP對H2O2誘導的HL-7702細胞MAPK、NF-κB活化的影響
ROS能夠引起MAPK、NF-κB通路的激活[20]。BRP對H2O2誘導HL-7702細胞MAPK、NF-κB活化的影響如圖9所示。H2O2對HL-7702細胞ERK、JNK、p38、NF-κB總蛋白表達以及p38磷酸化無顯著影響(P>0.05),但上調HL-7702細胞ERK、JNK磷酸化以及NF-κB p65核轉移(P<0.05);而BRP顯著降低ERK、JNK磷酸化水平以及NF-κB p65核轉移水平(P<0.05)。提示,BRP可能是通過ERK、JNK及NF-κB通路來調控凋亡相關蛋白的表達,從而抑制H2O2致肝細胞凋亡的。
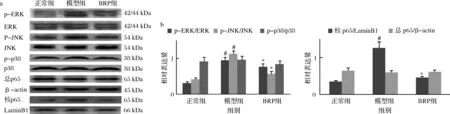
a-蛋白印跡結果;b-灰度比值結果圖9 BRP對HL-7702細胞MAPK、NF-κB激活的影響Fig.9 Effect of BRP on activation of MAPK and NF-κB in HL-7702 cells
4 結論與討論
肝細胞損傷是多種肝病的共同病理基礎,而ROS引發的氧化應激是造成肝細胞損傷的重要機制[4]。H2O2是實驗室常見的氧化應激損傷劑,極易透過細胞膜對肝細胞造成損傷、甚至死亡[3-4]。本研究H2O2誘導的HL-7702細胞損傷入手,運用現代分子生物學與分子藥理學方法,探討BRP對肝細胞氧化應激損傷的保護作用。結果表明,H2O2能夠降低HL-7702細胞存活率、升高LDH釋放率、升高細胞MDA水平,提示HL-7702細胞氧化損傷模型構建成功。而安全劑量的BRP預處理HL-7702細胞后,BRP組HL-7702細胞存活率與模型組比較顯著增高,LDH釋放率和細胞MDA生成量顯著下降,細胞凋亡程度顯著減少,表明BRP能夠保護H2O2誘導的HL-7702細胞損傷,抑制細胞凋亡。
線粒體是細胞內最易受ROS攻擊的目標之一,因此在H2O2誘導的肝細胞凋亡中處于重要地位[23]。而Bcl-2家族在線粒體途徑細胞凋亡的發生中起著核心作用[24]。目前,Bcl-2家族蛋白中研究最多的主要有2種,一種是促凋亡蛋白,如Bak、Bax等,另外就是抗凋亡蛋白,如Bcl-2、Bcl-xL等,二者之間比例決定著損傷細胞走向生存或者死亡[24]。促凋亡蛋白和抗凋亡蛋白的比值增高則造成線粒體膜電位降低,促進Cyto c從線粒體釋放入胞質,繼而激活下游的Caspase家族,形成Caspase 3活化片段#C-casp3,導致一系列凋亡特征性形態變化和生化改變,并切割其特異性底物PARP形成C-PARP,最終導致細胞凋亡[24-25]。而p53能介導DNA損傷后的細胞應激反應,若損傷DNA修復失敗可與其他凋亡蛋白協同誘導細胞凋亡的發生[26]。研究表明,BRP預處理能夠降低HL-7702細胞漿Cyto c水平,下調促凋亡蛋白p53、Bak、Bax表達,上調抗凋亡蛋白Bcl-2表達,降低C-casp3和C-PARP水平。提示,BRP可能是通過調控p53以及Bcl-2家族蛋白來抑制H2O2致HL-7702細胞凋亡的。MAPK是肝細胞內促細胞增殖和傳遞應激信號的關鍵激酶,主要包括ERK、JNK、p38等3條途徑,分別調控細胞生存、凋亡以及炎癥等,且協同和拮抗共存,三者與肝細胞凋亡的發生均有著密切相關性,在上游活化子以及下游靶點之間存在著交叉和重疊[27]。NF-κB是近期研究較多的轉錄因子,調控多種與炎癥相關細胞因子的轉錄,在肝損傷的發生發展中也起重要作用[28]。因此,MAPK、NF-κB通路與肝細胞氧化應激損傷的關系一直是肝疾的研究熱點[27-29]。本研究表明,BRP預處理能夠降低ERK、JNK磷酸化以及NF-κB p65核轉移水平。提示,BRP可能是通過ERK、JNK以及NF-κB通路調控凋亡相關蛋白的表達從而抑制H2O2致HL-7702細胞凋亡的。綜上所述,BRP能夠有效保護H2O2誘導的HL-7702細胞損傷,并抑制細胞凋亡,此作用可能與BRP對ERK、JNK、NF-κB通路的調控有關。此外,NF-κB通路又是與炎癥密切相關的重要信號通路。BRP能否通過NF-κB通路影響肝細胞以及巨噬細胞的炎癥反應仍需進一步研究。
猜你喜歡
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中國塑料(2016年3期)2016-06-15 20:30:00
西南軍醫(2016年6期)2016-01-23 02:21:19
西南軍醫(2015年2期)2015-01-22 09:09:37
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
