不同水質調控方式對海參池塘沉積物酶活性的影響
2021-11-19 05:37:14王祖峰李東萍魯曉倩畢麗仙周瑋
水產學雜志 2021年5期
關鍵詞:水質
王祖峰,李東萍,魯曉倩,畢麗仙,周瑋
(1.全國水產技術推廣總站,中國水產學會,北京 100125;2.中國農業大學,北京 100083;3.上海安譜實驗科技股份有限公司,上海 200000;4.大連海洋大學,遼寧 大連 116023)
池塘沉積物是水體沉降作用在池底表面形成的活性物質[1,2],是水生生態系統物質遷移轉化的最重要組分之一[3]。沉積物中的酶是物質循環和能量代謝的主要參與者,在養殖水體的有機質降解、營養元素的循環和轉換中發揮重要作用[1,4-13]。常用脲酶活性評價沉積物向水體供應無機氮的能力。相關研究認為,養殖池塘沉積物的脲酶活性與水體中NH4+-N 含量呈顯著負相關[14]。張宇等[15]報道,富營養化程度較為嚴重的大通湖脲酶活性(0.41 mg/g)顯著高于富營養化程度輕的湖泊脲酶活性[0.29~0.32 mg/(g·d)]。脫氫酶活性多用于分析沉積物活性微生物量及其對有機物降解活性的能力。伏小勇等[16]利用蚯蚓處理污泥時發現,脫氫酶活性和有機質存在顯著正相關性(P<0.01),當微生物量提高80%時加速了有機質的降解,后期脫氫酶穩定在1.03mg/(g·h)。磷酸酶活性通常反映沉積物有機磷的分解能力,或體現沉積物供磷能力。田秀平等[17]報道,兩種池塘中酸性磷酸酶活性與中等活性有機磷之間均呈顯著正相關,兩池塘相關系數分別為0.8559 和0.8269。李越蜀等[18]比較了不同養殖模式池塘,認為水質好的池塘沉積物中脲酶、脫氫酶、磷酸酶活性明顯較低。因此,在不同的沉積物有機質環境下,脲酶、脫氫酶、磷酸酶與微生物活性密切相關且反應靈敏,是反映環境質量,指導水產養殖生產的有效指標。酶活性與沉積物氮、磷和有機物代謝關系密切。
海參養殖是我國北方海水養殖的支柱產業之一,2019 年全國養殖面積已達24.67×104hm2,產量17.17×104t[19],其中池塘養殖是人工養殖主要方式,在產業發展中占據重要地位。利用潮汐水位變化,自然納潮換水是傳統池塘養殖唯一的水質調控方式,引發了生產波動,甚至海參死亡事件時有發生[20]。長期以來,研發有效的水質調控技術,擺脫靠天吃飯的管理困境是海參養殖穩產高產的關鍵。2007 年劉勃、蔣國春等[21,22]從提高溶解氧的角度開發了微孔曝氣水質調控技術,用管道向池塘底部輸入高壓空氣,可將水體溶氧含量提高2~3 mg/L,改善水質效果顯著,但該技術應用后氣孔堵塞而無法維護等問題嚴重限制了微孔曝氣技術的應用[20]。本團隊在研究溫鹽躍層導致海參死亡課題中,研發了養水機[23]水質調控技術,旨在強制池塘上下層水體交換打破躍層,同時利用益生菌作用,實現水質調控。前期研究顯示,在養水機調控下,溫鹽躍層現象有效減少,沉積物有機質量降低,微生物多樣性增加,促進營養鹽釋放,生產上海參成活率可提高到95%,單產可提高30%[24]。
本試驗通過測定自然納潮、微孔曝氣、養水機三種水質調控方式下的海參池塘沉積物中脲酶、脫氫酶、酸性磷酸酶、微生物活性和有機質含量的周年變化,比較三種水質調控方式的效果,進一步分析養水機調控水質的機理,以期為養水機的推廣應用提供科學依據。
1 材料與方法
1.1 試驗池塘
試驗在遼寧省莊河市大連寶發海珍品有限公司長605 m、寬85 m、深1.2~2.0 m 的9 口海參池塘中進行。池塘泥沙底質,均勻鋪設海參網礁,南北各有一進、排水閘門,每月大潮期間(初一、十五)換水3~5 d。各池塘內海參放養規格、密度相同,試驗期間不投餌、不投藥,統一管理。試驗池塘采用三種水質調控方式:自然納水;池底鋪設微孔曝氣微裝置,當池塘缺氧時,0.15 W/m2空壓機開始工作;池塘最深處安置功率750 W 的養水機,每天從21:00~次日9:00 工作12 h。每種水質調控方式池塘各3 個。
1.2 養水機結構與原理
養水機[CN200620006141.5]主要分為四部分:進水組件[CN2016213645 76.7]、生物包、水動力裝置和養殖池塘專用噴頭[CN201410415402.8][23]。
1.3 試驗方法
2017 年1 月—12 月,每月大潮前3~5 d 在池塘南端最深處采樣。
沉積物采集:參照《海洋調查規范》(GB/T12763-2007)第6 部分規定的方法采樣。在采樣點用直徑5 cm 圓形柱狀采泥器采集5 cm 底泥3 份,裝入滅菌自封袋,4℃冷藏帶回實驗室,去除樣品中根系、貝殼等雜質,風干至恒重后研磨、過篩絹(100 目)、混勻裝袋,4℃下保存待測。
酶活性測定:按照關松蔭[25]的方法,采用比色法測定脲酶活性、氯化三苯基四氮唑(TTC)比色法測定脫氫酶活性、磷酸苯二鈉比色法測定酸性磷酸酶活性。按照金相燦[3]的方法,采用熒光素雙醋酸酯法測定微生物活性。按照重鉻酸鉀還原—容量法(GB17378.5-2007)測定有機質含量。相關試劑材料購自上海索橋生物科技有限公司。
數據處理:取每種水質調控的3 個平行池塘的平均數。使用Excel 2013 和SPSS17.0 對試驗數據進行處理和方差分析,以皮爾森系數表示相關性,P<0.05 為相關,P<0.01 為極相關。
2 結果與分析
2.1 不同水質調控方式池塘沉積物中脲酶活性的周年變化
由圖1 可知,三種池塘沉積物脲酶活性全年變化趨勢較一致,整體活性范圍在95.00~279.42 U/g之間,1—4 月脲酶活性(180.66~279.42 U/g)高于5—12 月(95.00~249.70 U/g)。自然納水池塘以1月、2 月、3 月、4 月和9 月為代表8 次處于最高水平,10 月、12 月處于最低水平;微孔曝氣池塘以2月、3 月、4 月、9 月為代表8 次處于中間水平,6 月、7 月、10 月、12 月4 次處于最高水平;養水機池塘10 次處于最低水平,僅10 月、12 月處于中間水平。變化趨勢表明,3 月微孔曝氣池塘和養水機池塘顯著大跌至223.48 U/g 和195.51 U/g,7 月微孔曝氣池塘和自然納潮池塘顯著上漲到187.65 U/g 和171.04 U/g,9 月自然納潮池塘顯著增長到249.70 U/g,10月下降到年較低值122.10 U/g。三種水質調控方式下池塘沉積物脲酶活性波動極差值由高至低依次為:自然納潮(181.79 U/g)>微孔曝氣(147.71 U/g)>養水機(131.10 U/g)。
2.2 不同水質調控方式池塘沉積物中脫氫酶活性的周年變化
由圖2 可知,三種池塘沉積物脫氫酶活性全年變化趨勢較一致,活性范圍在1.50~36.84 U/g 之間。7—9 月脫氫酶活性(20.33~36.84 U/g)高于1—6 月和10—12 月(1.50~25.17 U/g)。自然納潮池塘在1—7 月、10 月、12 月為代表9 次處于最高水平,8月處于中間水平,9 月、11 月處于最低水平;微孔曝氣池塘以1 月、3 月、5—7 月、9 月、10 月為代表7次處于中間水平,8 月1 次處于最高水平,2 月、4月、12 月3 次處于最低水平;養水機池塘1 月、3月、5—8 月、10 月、11 月8 次處于最低水平,2 月、4月、12 月3 次處于中間水平,9 月1 次處于最高水平。從變化趨勢比較上看,1—5 月自然納潮池塘顯著高于另外兩組。三種方式全年波動趨勢基本一致,波動極差值微孔曝氣(35.34 U/g)>養水機(26.67 U/g)>自然納潮(23.84 U/g)。
2.3 不同水質調控方式池塘沉積物中酸性磷酸酶活性的周年變化
由圖3 可知,三種池塘沉積物酸性磷酸酶活性全年變化趨勢不一致,活性范圍在87.27~973.38 nmol/(g·h)之間。自然納潮池塘沉積物中酸性磷酸酶活性年變化范圍為134.26~839.12 nmol/(g·h),極差為704.86 nmol/(g·h),平均值為470.47 nmol/(g·h);微孔曝氣池塘沉積物酸性磷酸酶活性年變化范圍為208.10~973.38 nmol/(g·h),極差為765.28 nmol/(g·h),平均值為564.80 nmol/(g·h);養水機池塘沉積物酸性磷酸酶活性年變化范圍為87.27~537.04 nmol/(g·h),極差為449.77 nmol/(g·h),平均值為303.14 nmol/(g·h)。三種調控方式相比,養水機池塘沉積物酸性磷酸酶活性年平均值最小,波動最小;微孔曝氣池塘沉積物酸性磷酸酶活性年平均值最大,波動最大。
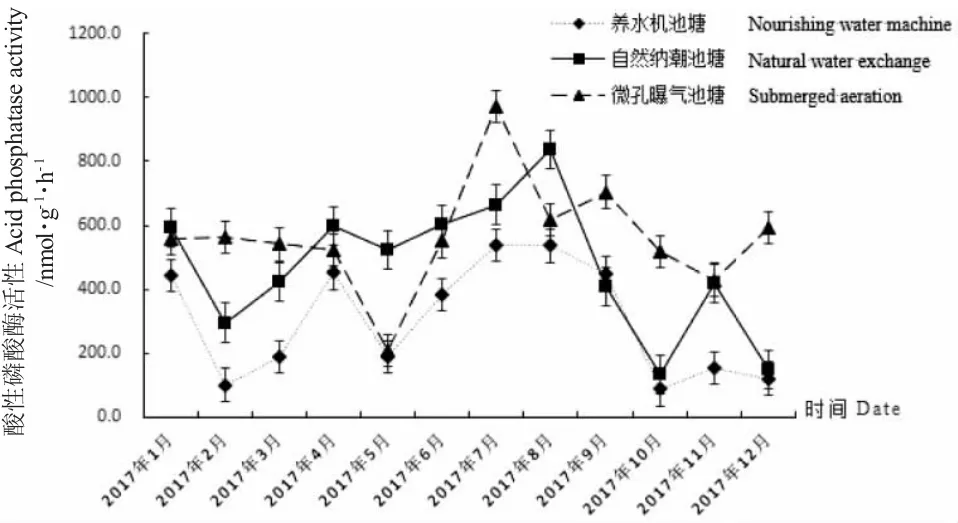
圖3 試驗池塘沉積物酸性磷酸酶活性周年變化Fig.3 The annual changes in acid phosphatase activity in sediments of the experimental ponds
2.4 不同水質調控方式池塘沉積物中微生物活性的周年變化
由圖4 可知,三種池塘沉積物中微生物活性全年變化趨勢大致一致,活性范圍在1.66~14.81 μg/(g·min)之間。自然納潮、微孔曝氣和養水機三種池塘沉積物微生物活性均在3 月達最大值,分別為8.71 μg/(g·min)、14.81 μg/(g·min)和7.68 μg/(g·min),10 月達最低值,分別為2.24 μg/(g·min)、3.16 μg/(g·min)和1.66 μg/(g·min)。自然納潮池塘沉積物微生物活性年變化極差為6.47 μg/(g·min),平均值為4.73 μg/(g·min);微孔曝氣池塘沉積物微生物活性年變化極差為11.65 μg/(g·min),平均值為7.38 μg/(g·min);養水機池塘沉積物微生物活性年變化極差為6.02 μg/(g·min),平均值4.60 μg/(g·min)。三種調控方式相比,養水機池塘沉積物微生物活性年平均值最小,波動最小;微孔曝氣池塘沉積物微生物活性年平均值最大,波動最大。
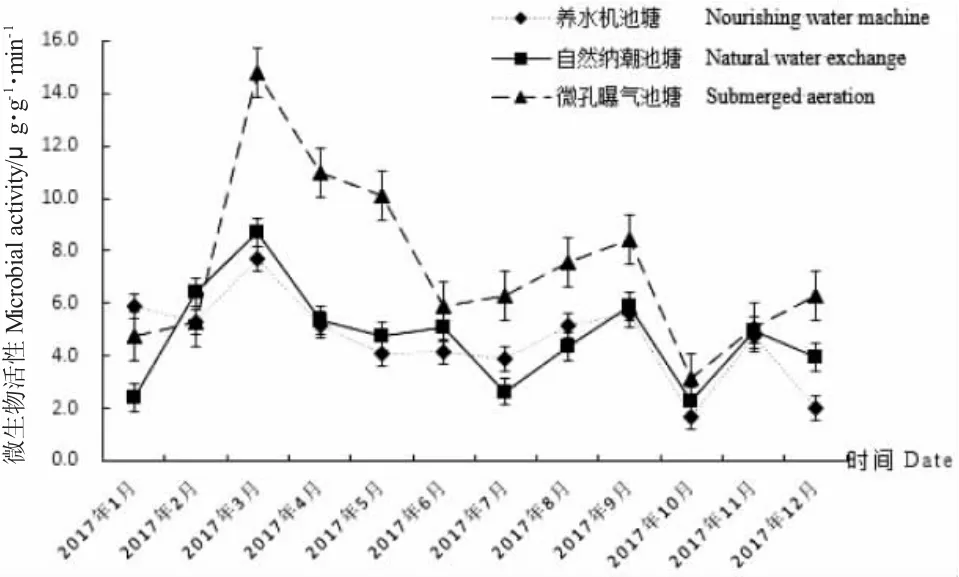
圖4 試驗池塘沉積物中微生物活性的周年變化Fig.4 The annual changes in microbial activity in sediments of the experimental ponds
2.5 不同水質調控方式池塘沉積物有機質含量周年變化
由圖5 可知,三種池塘沉積物中有機質全年變化趨勢不一致,含量范圍在0.07%~2.60%之間。自然納潮池塘沉積物有機質含量年變化范圍為1.05%~2.18%,極差為1.13%,平均值為1.54%;微孔曝氣池塘沉積物有機質含量年變化范圍為1.19%~2.60%,極差為1.41%,平均值為1.89%;養水機池塘沉積物有機質含量年變化范圍為0.07%~2.04%,極差為1.97%,平均值為1.07%。三種調控方式相比,養水機池塘沉積物有機質含量年平均值最小,微孔曝氣池塘沉積物有機質含量年平均值最大。
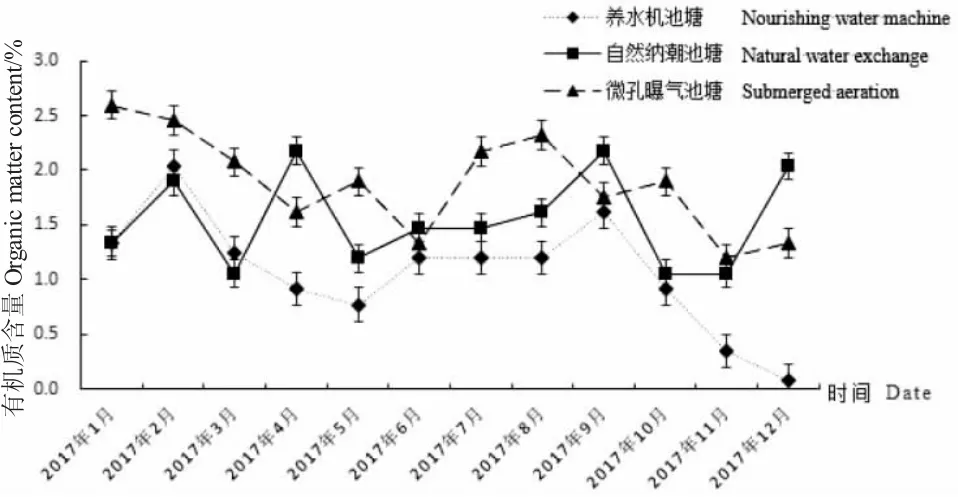
圖5 試驗池塘沉積物有機質含量周年變化Fig.5 The annual changes of organic matter contents in sediments of the experimental ponds
3 討論
3.1 三種水質調控方式池塘沉積物酶活性周年變化分析
本試驗三種水質調控方式池塘沉積物中脲酶、脫氫酶、酸性磷酸酶活性的周年變化趨勢與相關研究結論基本一致。相關研究認為,酶活性受溫度影響最大,水溫升高可促進沉積物微生物的活動,表現為酶活性的增大[26-28]。甘茂林等[29]認為,有機質也可以影響池塘沉積物中脲酶活性,大亞灣網箱養殖區表層沉積物有機質含量高于非養殖區35.2%,伴隨著細菌降解作用,脲酶活性明顯高出其他站位50.7%。本試驗中5—7 月,池塘沉積物微生物活性和有機質含量均處于較低水平,因而其脲酶活性明顯處于低位。劉梅等[30]、黃睿智[31]認為,植物或藻類對有機質降解也是影響池塘沉積物中脫氫酶活性的重要原因。有機物降解時,微生物量增加,脫氫酶活性緩慢增加。藻類和外源輸入因子的季節變化,產生的分泌物和外源營養物質向沉積物的輸入差異,可以帶來與本研究一致的池塘沉積物脫氫酶活性的季節差異。P 濃度也是影響沉積物酸性磷酸酶活性的重要原因[32-34],酸性磷酸酶可促進有機磷分解生成無機磷。沉積物磷濃度低時,酶作為催化劑使反應向正方向移動,酸性磷酸酶活性持續升高。本研究中沉積物微生物活性和有機質研究結果,較好地體現在沉積物脲酶、脫氫酶、酸性磷酸酶活性中,整體上形成了因果關系。但是,在有機質降解過程中三種酶與菌群結構的對應關系,菌群結構與有機質中營養元素的對應關系尚需要進一步研究。
3.2 三種水質調控方式池塘沉積物酶活性比較
海參養殖池塘環境會發生極大的變化,沉積物酶活性對環境變化敏感,活性變化可以及時反映出沉積物質量的變化。三種水質調控方式比較表明,養水機調控的池塘水體指標值均處于最低值,自然納潮調控方式的脲酶、脫氫酶兩項指標值的交替變化中略顯優勢,微孔曝氣方式的酸性磷酸酶活性、微生物活性、有機質含量指標值的交替變化中略顯優勢。根據甘茂林等[29]、劉梅等[30]、黃睿智[31]、蔡樹美等[34]觀點,在微生物對沉積物有機質的降解過程中,微生物指標受溫度和有機質水平的影響,影響了脲酶、脫氫酶、酸性磷酸酶的活性,可以在養水機調控方式相關觀測結果中得到系統印證。本研究中,微孔曝氣池塘沉積物初始有機質含量(2.60%)顯著高于另外兩種池塘(自然納潮池塘1.33%,養水機池塘1.33%,P<0.05),微孔曝氣調控方式在脲酶、脫氫酶、酸性磷酸酶、微生物活性等指標值的交替變化與沉積物中有機質含量波動有關。
3.3 三種水質調控方式對沉積物作用機理分析
在三種水質調控方式中,納潮換水方式海參池塘獲得的沉積物有機質量最上,在沉積物有機質自然沉降,但躍層現象會影響沉降效果;微孔曝氣方式是高壓空氣從池底充入,氣泡與水體接觸增加了水體溶解氧,也對水體形成了擾動;養水機方式是在動力作用下將表層海水由底層射出,強制形成水體對流,也對水體形成了一定的擾動[23,35,36],這兩種方式均能破壞水體躍層現象。皮坤等[37]、任貽超[38]、趙文等[39]研究表明,水體內部的擾動有助于懸浮顆粒物質的沉積,導致沉積有機質含量的增加。從這個層面上看,微孔曝氣方式和養水機方式都會導致池塘由納潮而引入池塘的有機質沉積量增加。然而,對上述三種水質調控池塘能量代謝周年比較發現,養水機池塘的N、P 代謝全年均處于由池底沉積物向上層水體釋放的狀態,而自然納潮方式和微孔曝氣方式在3 月、7 月、8 月則處于由上層水體向池底沉積物積累的狀態(另文報道)。由此可以解釋養水機沉積物有機質含量最低的原因。本試驗結果顯示,微孔爆氣池塘沉積物有機質含量最高,這與大部分學者的研究結果不同,分析原因除了微孔爆氣池塘初始有機質含量最高外,可能還由于試驗過程中,微孔爆氣過程不及時或時間較短,影響了對沉積物有機質的分解能力。
結論:養水機能明顯降低沉積物酶活性、微生物活性和沉積物有機質含量,周年變化相對穩定,有效降低水質富營養化水平,改善水質,為海參生長提供良好的環境。實際生產中,兼顧海參健康生長和降低養殖成本,應在海參最適生長時期堅持使用養水機,在春季冰層融化和秋季結冰前適當延長養水機工作時間,池塘結冰后可暫停使用。
猜你喜歡
環境(2023年5期)2023-06-30 01:20:01
小星星·閱讀100分(低年級)(2022年6期)2022-08-22 02:33:40
供水技術(2021年3期)2021-08-13 09:08:34
建材發展導向(2019年10期)2019-08-24 06:25:02
當代水產(2019年1期)2019-05-16 02:42:04
當代水產(2019年3期)2019-05-14 05:42:48
電子制作(2018年14期)2018-08-21 01:38:16
水利規劃與設計(2018年1期)2018-01-31 01:53:53
水利規劃與設計(2016年7期)2016-02-28 15:06:27
世界文學評論(2014年2期)2014-04-12 06:23:43
