羅氏沼蝦及蔬菜在日光溫室內不同養殖系統中生產性能的比較研究
2021-11-19 05:37:42王大賀陳旺蔡鵬飛劉少貞王偉偉劉青
水產學雜志 2021年5期
王大賀,陳旺,蔡鵬飛,劉少貞,王偉偉,劉青
(山西農業大學動物科學學院,山西 太谷 030801)
當前天然漁業資源量不斷減少,而人們對水產品的需求量卻不斷增加,水產養殖在漁業領域中的貢獻比率日益增大[1]。傳統池塘高密度養蝦模式的發展多是以消耗自然資源、污染環境為代價,工業化農業生產對能源密集型化肥生產和大量水資源的依賴引發了可持續性的大問題。在社會生活中,生產中污水處理、農業灌溉、畜牧業等無不將矛頭指向水資源的合理使用[2]。當前,全國范圍內正在限制和取締污染高的“小棚”養蝦模式,縱觀歷史,農業的生產以各種形式融入并適應社會發展,特別是近現代,城市的快速發展,城市通過貿易和創新的中心地位來激勵農業的發展[3],集約、創新的農業生產形式的生存必須依賴創造更有價值的產品。蝦菜共生系統可以很好地解決污染外排的問題,提供更健康的產品和可持續發展的健康水產養殖業。蝦菜共生體系將蝦的循環水養殖和水耕栽培立體結合,以實現節能減排、節約土地、充分利用養殖空間[4]。系統中的物質和能量良性循環使物質能量朝著蝦菜雙方流動,而不產生外排污染物,是典型的生態循環農業經濟模式[5-7]。對魚菜共生的研究相對較多,而蝦菜共生,尤其是日光溫室內的蝦菜共生系統研究卻鮮見報道。本試驗比較研究了日光溫室內的蝦菜共生系統、土工膜覆蓋池及室內循環水養殖系統的生產效能,以期為推進該綜合種養模式的應用提供參考。
1 材料與方法
1.1 試驗場所、設施和運行
本試驗中,蝦菜綜合種養系統和土工膜覆蓋池在相同的日光大棚內進行,循環水養殖系統在室內進行。日光大棚配置有卷簾、遮陽網、照明燈等環境控制設備和在線水質監測器(購于幫邦水產)實時監測水溫、溶解氧和pH 變動。綜合種養系統由自行設計定制的軸長6 m×3 m 的橢圓形PVC 框架魚池組成。魚池由鋼架吊裝固定,上面固定六根相通且串聯并列的種植管道(內徑9.3 cm,長3.03 m),管道向上面打孔作栽培孔,左側3 根管道中放有火山巖和陶粒,右3 根則無基質[8-10],管道從左到右編號1~6。框架魚池內配備潛水泵(開40 min/h,關20 min/h)、過濾器、增氧機及水氣輸送管等裝置。蝦菜共生系統定時間隔循環水運行[11],潛水泵將養殖水提升至高位過濾桶(濾袋、火山巖、陶粒、生物球、過濾棉等材料)后,自流輸送至栽培管道內經蔬菜根部吸收后流回養殖池內。采用高壓渦輪風機(開20 min/h,關40 min/h)和配套氣管路通過微孔增氧管曝氣。高溫期以遮陽網控制光照強度并配合通風降溫。試驗過程中不排換水,僅補充自來水。整套系統水流量為1.5 m3/h,循環率為2.5 次/d,栽培區水流速約為3.68 m/min[12]。
土工膜覆蓋池均為漁業用的HDPE 土工膜全部覆蓋的土池,長寬深尺寸為10 m×5 m×1.5 m,也設于同一日光溫室大棚內,根據水質情況排換水,池內微孔底部增氧與種養系統同步啟閉。
溫室大棚操作室內循環水養殖系統由上下三層共12 個獨立進排水的40 cm×30 cm×30 cm 養殖箱和100 cm×40 cm×30 cm 的過濾箱組成,自然光照,循環過濾箱內有循環過濾、生物凈化、增氧、溫控等設備。
1.2 試驗方法
購于浙江市場的體質量0.0083 g 的羅氏沼蝦苗按表2 密度投放于大棚內蝦菜共生系統、覆膜池以及室內循環水養殖系統中。試驗期間,羅氏沼蝦的日投餌率為4%~6%,根據蝦攝食情況、天氣、水質調整,每日記錄實際投喂量;每周測量水體的氨氮、亞硝酸鹽、磷酸鹽、硫化物和溶解氧含量、pH 及溫度[13]。每隔15 d 測量羅氏沼蝦體質量、體長和蔬菜的重量、根上植株高。蔬菜苗種采用基質育苗盤自培,種類有小白菜、早熟大白菜、苣Lactuca sativa Linn.、芫荽Coriandrum sativum Linn.、雍菜Ipomoea aquatica Forsskal 等,蔬菜的種植密度為26 株/m2[14],采用防蟲網以及藍板和黃板粘蟲,防治蔬菜蟲害。
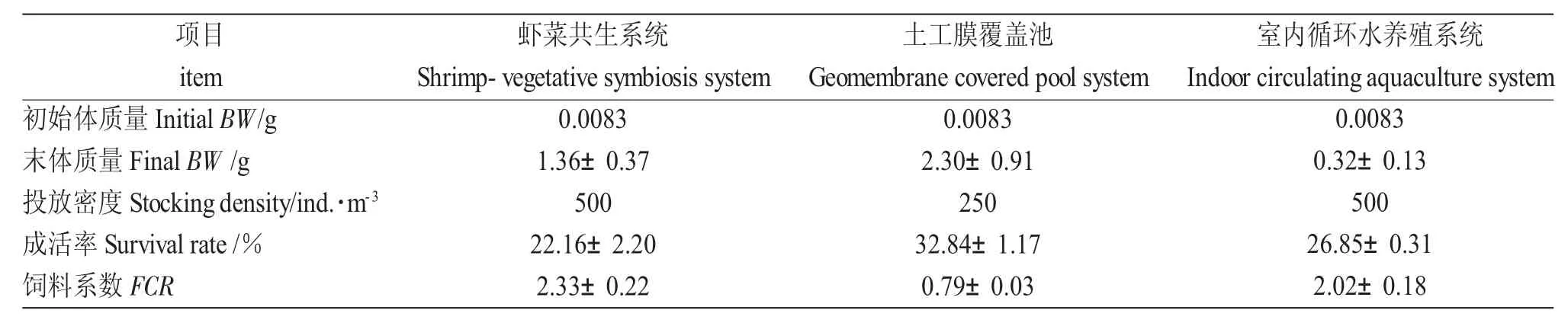
表2 各養殖系統中羅氏沼蝦試驗前后的體質量、投放密度、成活率及餌料系數Tab.2 Body weight(BW),stocking density,survival rate and food conversion ratio(FCR)of giant river prawn at the beginning and end of the experiment
1.3 數據采集與處理
蝦的生長期為75 d,第一批蔬菜小白菜和早熟大白菜的生長期是45 d,第二批蔬菜小白菜、生菜、香菜受溫度限制生長期是30 d,測量周期是15 d/次。測重時用吸水紙拭干蝦和蔬菜表面水分,用游標卡尺(精度為0.1 mm)和電子天平(精度為0.01 g)測定。蝦和蔬菜迅速測量完后立即放回水中。水質檢測方法參考文獻《養殖水環境化學實驗》[15]。
蝦和菜15 d 增重率和特定生長率的計算公式:
增重率WGR(%)=100×(Wt-Wt-1)/Wt-1;
特定生長率SGR(%/d)=100×(lnWt-lnWt-1)/D,其中Wt指第t 次測量蝦的平均體質量,Wt-1 指第t-1 次蝦的平均體質量,D 為飼養時間(d)。
利用SPSS 22.0 統計軟件進行數據處理與分析,試驗數據采用單因素方差分析、LSD 多重比較以及一般線性模型等方法進行分析,差異顯著水平設為P<0.05,利用GraphPad Prism5 軟件作圖。
2 結果與分析
2.1 基礎水質情況
試驗期間,蝦菜共生系統和室內循環水系統只補充自來水,不換水。土工膜覆蓋池水質的某項指標出現較大變動時每次更換1/4 的水體,以保證羅氏沼蝦的正常生[16,17]。由表1 可知:試驗期間各系統的氨氮、亞硝酸鹽、硫化物、磷酸鹽含量均處于較低水平,符合養殖水質指標,其中蝦菜共生系統中氨氮含量更低、更穩定,水質均偏堿性;土工膜覆蓋池中藻類繁殖旺盛,pH 過高,共換水4 次;室內循環水pH 穩定(圖1),室內循環水硝酸鹽含量較另外兩系統高,且差異顯著(P<0.05)。相比而言,蝦菜共生系統中氨氮、硝酸鹽、pH、水溫、磷酸鹽等指標較穩定,溶氧處于較高水平(圖1~圖4)。在試驗期間水質條件可滿足羅氏沼蝦和蔬菜正常生長需求。
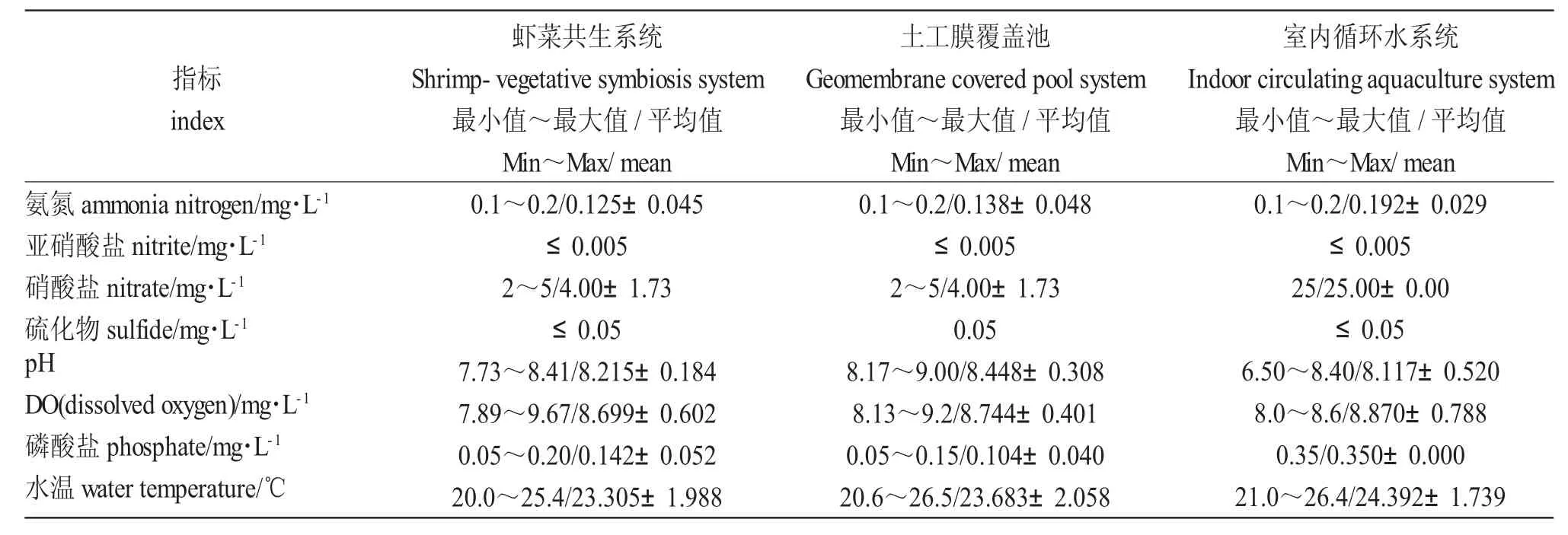
表1 各養殖系統中水質變化情況Tab.1 Changes in water quality in various aquaculture systems
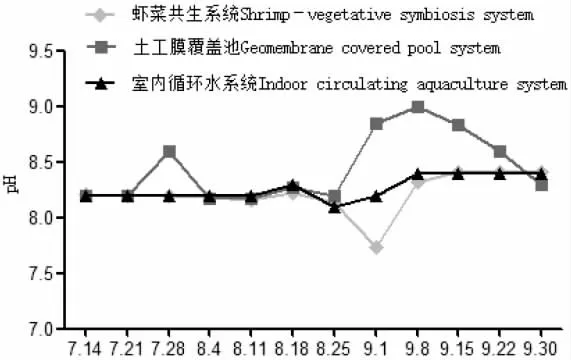
圖1 各養殖系統中pH 隨時間變化Fig.1 Change in pH level with time in various breeding systems
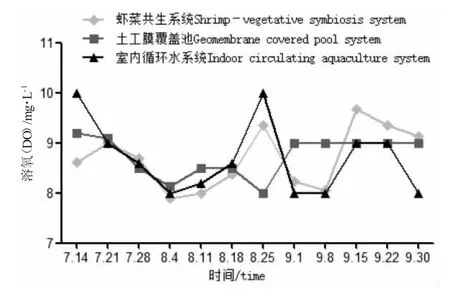
圖2 各養殖系統中溶解氧隨時間變化Fig.2 Change in dissolved oxygen level with time in various breeding systems
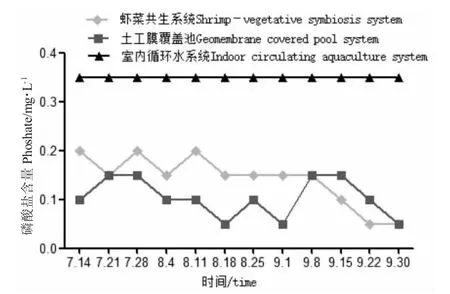
圖3 各養殖系統中磷酸鹽含量隨時間變化Fig.3 Change in phosphate content level with time in various breeding systems
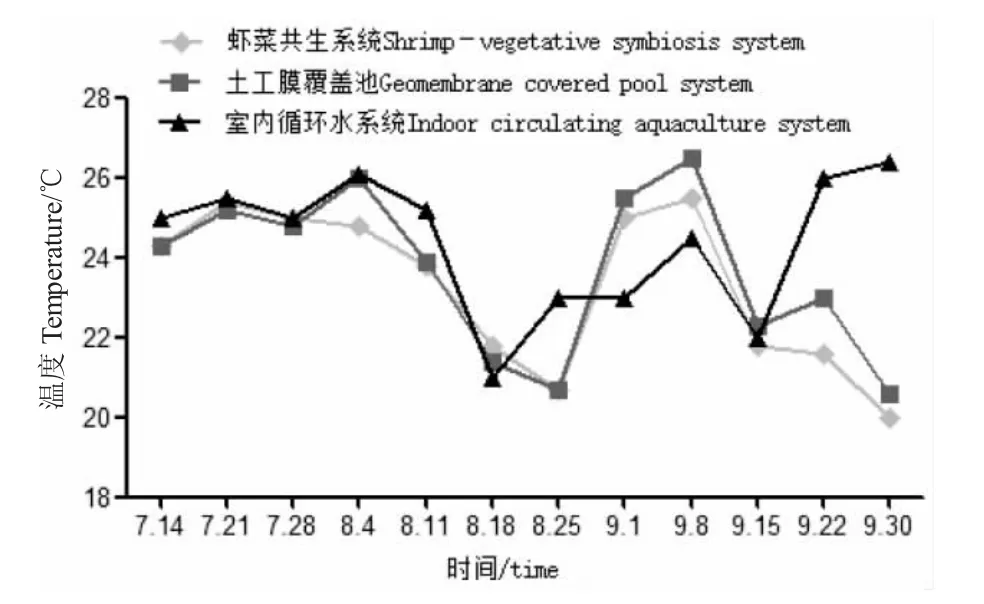
圖4 各養殖系統中溫度隨時間變化Fig.4 Change in temperature with time in various breeding systems
2.2 羅氏沼蝦的生長情況
由圖5、圖6 可知:試驗期間,土工膜覆蓋池羅氏沼蝦的生長速度顯著高于另外兩種系統(P<0.05);蝦菜共生系統中蝦的生長速度高于室內循環水養殖系統,試驗前期生長差異不明顯,后期差異顯著(P<0.05);蝦菜共生系統的羅氏沼蝦增長率、增重率、特定增長率和特定增重率均先緩慢再上升,而另外兩種系統均呈下降趨勢,養殖生產后期蝦菜共生系統是否更有優勢還有待研究。
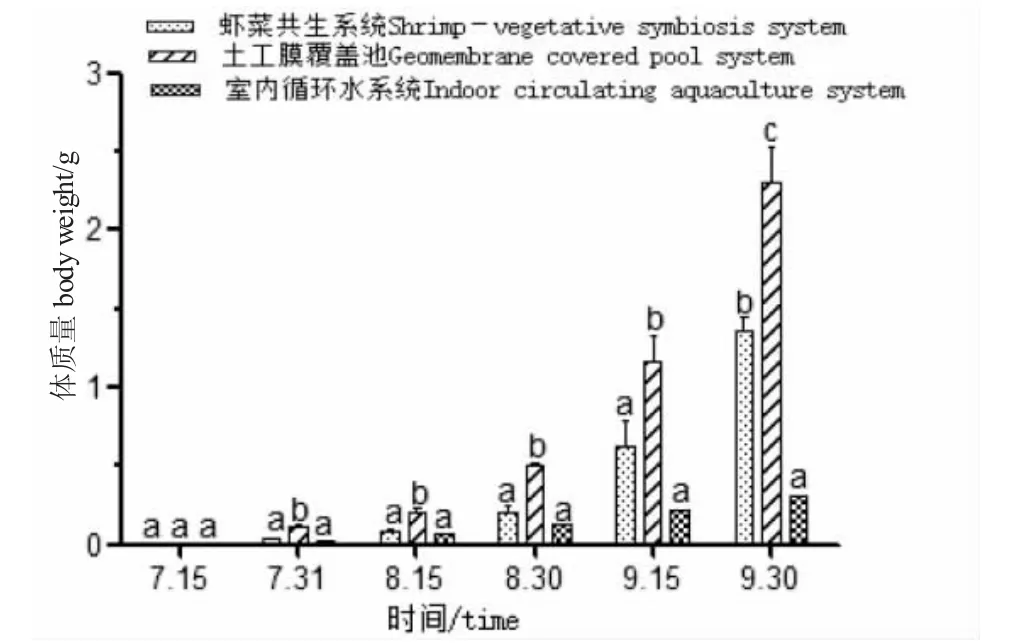
圖5 各養殖系統中羅氏沼蝦體質量隨時間變化Fig.5 Change in body weight of giant freshwater prawn with time in various breeding systems
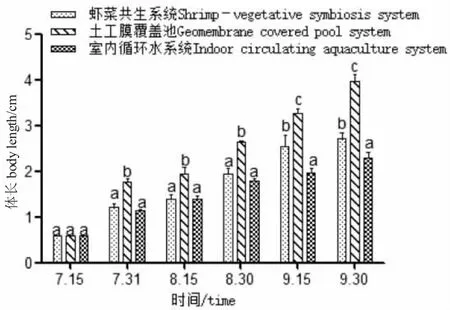
圖6 各養殖系統中羅氏沼蝦體長隨時間變化Fig.6 Change in body length of giant freshwater prawn with time in various breeding systems
2.3 共生蔬菜的生長情況
2.3.1 共生蔬菜的生長
試驗期間菜共生中,小白菜和早熟大白菜生長良好,后者的生長速度稍快于小白菜(圖7~圖9);45 d 內早熟大白菜平均增重31.01 g,小白菜平均增重16.26 g(表3);每個階段小白菜、早熟大白菜根上植株高的增長明顯不同,差異顯著(P<0.05),重量變化僅在后兩次測量時差異顯著(P<0.05);其增重率和特定增重率在各階段無明顯差異(P>0.05),而增長率和特定增長率最后一時間點有所降低,差異顯著(P<0.05)。小白菜成活率(94.8%)高于早熟大白菜(90.8%),這或與不同種類的蔬菜水培適應性有關[18,19]。
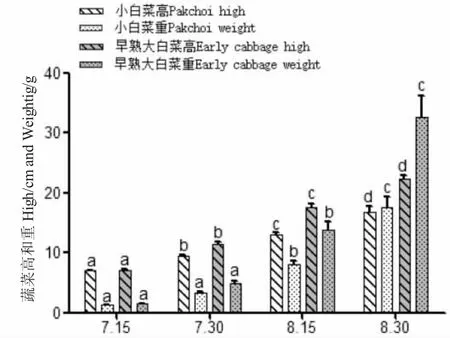
圖7 蝦菜共生系統中蔬菜高和重隨時間變化Fig.7 Change in vegetable high and weight with time in shrimp-vegetable symbiosis systems
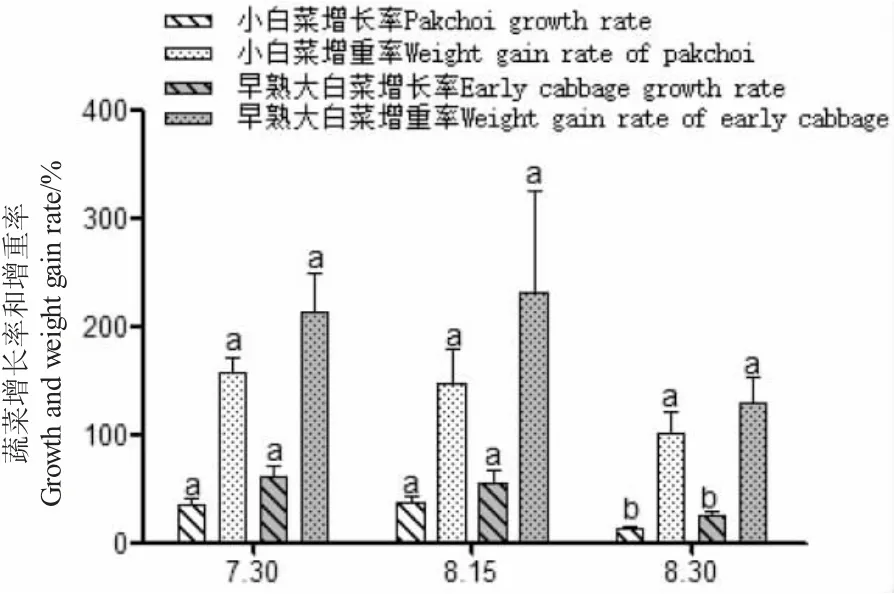
圖8 蝦菜共生系統中增長率和增重率隨時間變化Fig.8 Change in vegetable growth and weight gain rate with time in shrimp-vegetable symbiosis systems
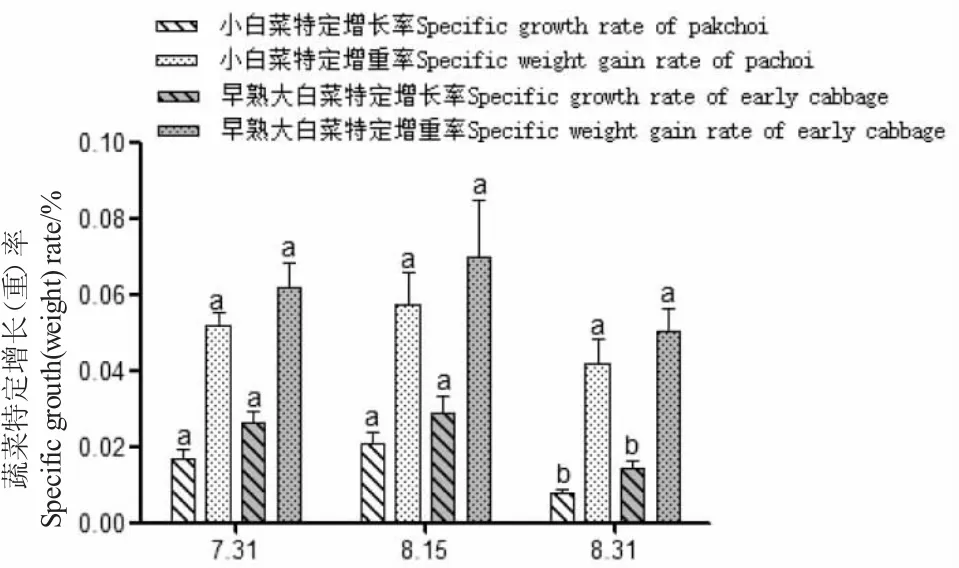
圖9 蝦菜共生系統中蔬菜特定增長率隨時間變化Fig.9 Change in specific growth rate of vegetable with time in shrimp-vegetable symbiosis systems
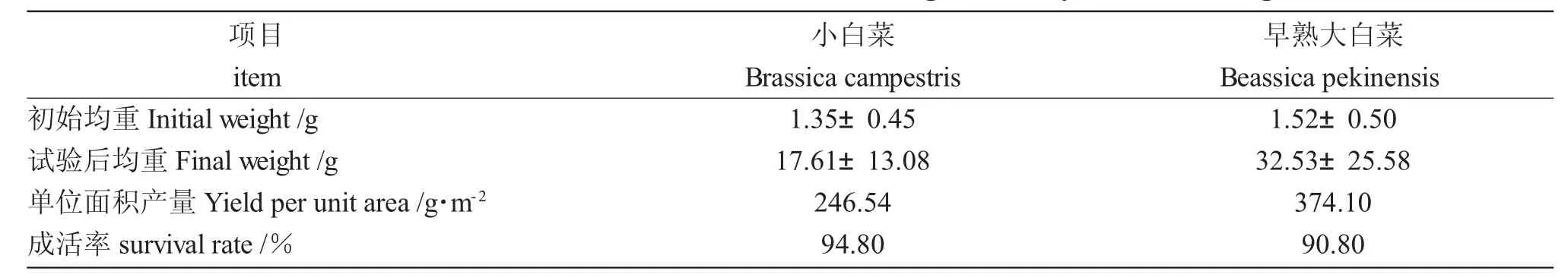
表3 第一批小白菜和早熟大白菜生長情況Tab.3 Growth of the first batch of Chinese cabbage and early Chinese cabbage
2.3.2 基質對蔬菜生長的影響
種植管道中有基質栽培的早熟大白菜質量增長略快于無基質(圖10),但差異不顯著(P>0.05),無基質栽培可以節約成本。結合水質指標,表明在規模化生產中可以采取無基質水培方式。蝦菜共生系統其他產出。
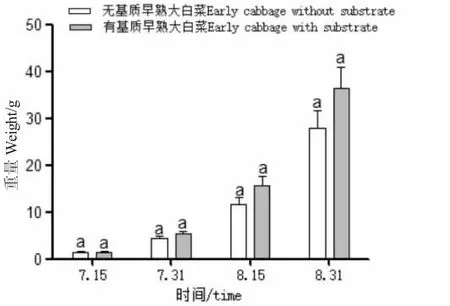
圖10 早熟大白菜在有無基質條件下重量隨時間變化Fig.10 Change in quality of Chinese cabbage with time in the presence or absence of substrate
除羅氏沼蝦和蔬菜,在蝦菜共生系統中投放浮萍Lemna minor Linn.和伊樂藻Elodea nuttallii 以凈化水質、遮陽、為蝦生長棲息和隱蔽場所,囊螺Physlla acuta 可以有效控制藻類密度和附著物,水芹Oenanthe javanica 定植管道中,定期剪收(表4)。

表4 蝦菜共生系統中副產物Tab.4 By-products in the prawn and vegetable symbiosis system
3 討論
3.1 三種系統中羅氏沼蝦的生長
比較發現:不同系統養殖羅氏沼蝦各有利弊,根據養殖的預期效果,可從不同角度觀察并選擇,以謀求最大的經濟效益和生態效益。但總體是:蝦菜共生系統較優于另外兩種系統。
蝦菜共生系統控制水質的優勢更突出,室內循環水養殖系統和土工膜覆蓋池略有不足。蝦菜共生池過濾器和基質以及植物根系具有物理過濾、微生物分解和吸收凈化。蔬菜生長良好,小白菜、早熟大白菜、萵苣等生長周期在30~40 d 即成品可食。副產物浮萍、水芹、螺生長旺盛,具有很強的凈水效果,并能為蝦提供天然餌料和棲息、避敵場所。在整個試驗過程中水體較清瘦,藻類稀少,水體透明度高,微藻、細菌等作為養殖池生態系統的重要組成部分,其群落結構與多樣性在水質調節、養殖生物的生長、疾病控制及維持生態系統的穩定性等方面發揮著重要作用[21]。蔬菜發達的根系以及基質火山巖和陶粒可附著微生物,是微生物良好的生長載體,可顯著降低水體中氮磷含量[21-23],減少了水體中一些不利于養殖生物生長的細菌數量,如變形桿菌Proteus sp.、擬桿菌Bacteroides sp.和假單胞菌Pseudomonas sp.等[24]。由表1 可知,蝦菜共生系統中氨氮最低水平,pH 最穩定,尤其在降低硝酸鹽和磷酸鹽含量,控制水質的能力優于室內循環系統。
室內循環水養殖系統有過濾和加溫裝置,可在一定程度上維持水體穩定;室內受氣候影響更小,養殖溫度更易控制。由圖4 可知,當溫度低于養殖適宜溫度時,可采用恒溫加熱器等調溫設施保持溫度適宜。然而,小水體和自然光照不足等因素限制藻類的生長,室內循環水養殖系統的磷酸鹽和硝酸鹽含量顯著高于其他系統(P<0.05),間接說明植物和藻類具有積極消除磷酸鹽、硝酸鹽積累的作用[25,26]。
土工膜覆蓋池養殖期間水質良好,水色呈茶褐色,含有豐富的微藻和菌群,為羅氏沼蝦生長提供了部分餌料和營養物質。土工膜覆蓋池水溫晝夜溫差小,保溫效果好,但土工膜覆蓋池水質最不穩定,特別是pH 易過高,養殖期間共換水4 次,每次換水1/4。
土工膜覆蓋池中羅氏沼蝦的生長最好,其次是蝦菜共生系統,最后是室內循環水養殖系統。
蝦菜共生系統中羅氏沼蝦的生長速度較另外兩種系統處于中等水平,飼料系數和成活率卻分別處于最高和最低。養殖試驗前期蝦的死亡率較高,推測原因:(1)水體浮游生物稀少,水體透明度大,蝦的應激較大,雖然有水草和浮萍等,但效果依然不好;(2)地上池加有保溫層但不如地下土工膜覆蓋池和室內養殖系統保溫好,前期天氣不穩定,對蝦的成活率的影響較大;(3)可能運行早期系統不夠成熟,放養密度偏高而系統水質凈化能力不足。確切原因可通過研究水體、蔬菜根系以及蝦腸道的微生物組成差異,了解微生物環境基因組[27],進一步探討影響實驗結果的微觀原因。
試驗后期室內循環水養殖系統中蝦的生長速度明顯低于另外兩種養殖方式。試驗結束時室內循環水養殖系統中羅氏沼蝦的平均體質量、成活率、飼料系數等指標不盡如人意。因此,室內立體循環系統還需進一步改進和完善,才能有穩定的生態效益和合理的經濟效益。
土工膜覆蓋池羅氏沼蝦的生長速度、成活率均高于其他系統,飼料系數明顯低于其他系統。由圖5、圖6 可知:試驗中后期,土工膜覆蓋池中蝦的生長明顯快于另兩種養殖方式,可能與餌料充足、水質良好及應激小有關。
本次試驗所用蝦苗受長途運輸、應激反應、放養密度大[28]等因素的影響,三種養殖模式的成活率或高于某些池塘養殖羅氏沼蝦模式[29],但不及池塘精養模式[30],生長性能高于實驗室內魚缸養殖羅氏沼蝦[31],低于池塘精養模式。
觀察比較發現:三種養殖模式中羅氏沼蝦的額劍和第一步足顏色不一。蝦菜共生系統中小部分羅氏沼蝦的額角尖端呈紅褐色,第一步足的掌節、腕節、長節部位呈紅色(圖11-b),另外一部分額角和第一步足顏色幾乎是無色透明;土工膜覆蓋池中絕大部分羅氏沼蝦的額劍和第一步足的掌節、腕節、長節部位呈淡青藍色(圖11-c);室內循環水養殖系統中兩種顏色類型的羅氏沼蝦均存在一小部分,另外一部分羅氏沼蝦額劍和第一步足顏色幾乎是無色透明(圖11-a)。其原因可能是水體透明度、環境顏色不一或環境脅迫[32]。當羅氏沼蝦長期處于氧化應激狀態時,體內游離的抗氧化物質蝦青素、VC、VE 含量下降[33],這種狀態的持續會導致身體某一部位顏色的改變。

圖11 各系統羅氏沼蝦的體色Fig.11 Body color comparison of giant river prawn M.rosenbergii in three systems
3.2 蝦菜共生系統中蔬菜生長情況
本試驗所用小白菜、早熟大白菜等苗種根部至少長4~6 cm,根上部分長6~8 cm 時即可移植至定植管道中,定植過程中避免損壞幼苗。試驗中蔬菜主要蟲害為蚜蟲,通過噴水和粘蟲板物理除蟲害。試驗所用蔬菜平均成活率為92.8%,即使是同一種蔬菜的生長狀況也有所不同,單個植株大小不均勻。水培蔬菜的生長周期與陸生基本一致。由圖10可知,早熟大白菜在有基質條件下生長速度略快于無基質條件,但差異并不顯著。有基質栽培的早熟大白菜根系更發達,扎根蓬松,而無基栽培蔬菜的根系在水流作用下聚集在一起,吸收養分的面積相對較小,所以有基質條件下的根系處理水的效果也優于無基質的根系。在滿足對水質凈化的前提下,可選擇無基質水培方式,減少火山巖、陶粒的使用,降低成本,減輕支持框架負荷,方便蔬菜的采收。
嘗試在蝦菜共生池內水面的泡沫浮板上定植的小白菜和早熟大白菜,但效果不佳,甚至有的植株慢慢枯死,而栽培管道中的蔬菜卻能正常生長,說明水流對蔬菜的生長有一定的促進作用,有利于根系對水中營養物質的吸收利用[34]。在硝酸鹽濃度為25 mg/L 的靜水水體中,定植在泡沫浮板上的小白菜、早熟大白菜生長明顯優于定植于水體硝酸鹽濃度為4 mg/L 的蝦菜共生池中。
3.3 結論
蝦菜共生系統將養殖池、過濾裝置、水耕栽培和循環凈化設施有機整合在一起,維持了水質的穩定性,達到了生態養殖、增加養殖容量、提高產量的目的[35]。本試驗系統設置于地面之上,便于操作管理、觀察蝦的生長和收獲,可大規模、高密度養殖。設置水質環境智能檢測系統,實時掌握養殖環境狀況[36,37],極大地節約了人力。蝦菜共生系統收獲了羅氏沼蝦、蔬菜、水芹等產物,但產量還有待提高,特別是提高蝦的成活率、減少應激等。鑒于養殖前期,在新建蝦菜共生系統內羅氏沼蝦成活率低于較室內循環水養殖系統,蝦苗可在室內循環水養殖系統內培育,待蝦苗體長達1.5 cm 左右時移入蝦菜共生系統。室內循環水養殖系統的中間培育方便收集轉移,但生長慢;在土工膜覆蓋池中間培育生長速度快,但收集轉移不便,環境溫度不易控制。結合不同養殖系統中各時段羅氏沼蝦的生長性狀,可以選擇不同的養殖系統進行養殖生產。可利用蝦菜共生系統優良的養殖條件,作為成蝦售前的暫養池,改善蝦的養殖環境,提高肥滿度,產量和質量。室內循環水養殖系統有待進一步改進,如適當補光、調節水交換率和改進養殖管理方法等,以更適合蝦類的循環水養殖。土工膜覆蓋池與傳統高密度池塘養殖一樣需要解決養殖污水處理問題,水質處理會加大養殖成本,可采取池中種草和池塘漂浮水培蔬菜進行綜合種養以達到水質調控減少換水的效果。
猜你喜歡
環境(2023年5期)2023-06-30 01:20:01
工業設計(2022年8期)2022-09-09 07:43:20
小讀者(2021年2期)2021-03-29 05:03:48
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
華人時刊(2019年13期)2019-11-17 14:59:54
當代水產(2019年1期)2019-05-16 02:42:04
文苑(2018年22期)2018-11-19 02:54:14
家庭影院技術(2017年9期)2017-09-26 03:41:45
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
