牛病毒性腹瀉病毒離子孔道蛋白p7多肽多克隆抗體的制備和鑒定
2021-11-19 03:07:30付強郭妍婷陳俊貞王金泉史慧君
生物技術通報 2021年10期
關鍵詞:檢測
付強 郭妍婷 陳俊貞 王金泉 史慧君
(新疆農業大學動物醫學學院,烏魯木齊 830052)
牛病毒性腹瀉病毒(bovine viral diarrhea virus,BVDV)是導致牛病毒性腹瀉(bovine viral diarrhea,BVD)的病原[1]。牛、羊、鹿等易感動物感染BVDV后,會造成腹瀉、急和慢性黏膜病、病毒血癥、免疫耐受和持續感染、免疫抑制、繁殖障礙等癥狀[2],是嚴重危害我國養殖業的重要傳染病之一。國際獸疫局(Office International Des Epizooties,OIE)將BVD定為B類傳染病,在我國進出口檢驗檢疫中也將其列為二類傳染病。
BVDV的非結構蛋白p7,位于E2蛋白與NS2蛋白之間,分子量約7 kD,是主要由疏水氨基酸構成的一個小分子肽[3]。由Harada和Carrere-Kremer等人最初提出BVDV和HCV的p7蛋白寡聚并形成離子通道[4]。因為p7的離子通道活性使其被納入在病毒孔蛋白(Viroporin)家族中,Viroporin是一種小的疏水蛋白,當這些蛋白質在宿主細胞膜中寡聚時,它們會形成親水孔,增加膜的通透性,進行離子和小分子運動,破壞細胞的許多生理特性[5-6]。大量研究發現,許多的病毒能表達膜蛋白結構,表現出細胞的離子通道樣的某些功能,以介導病毒進入細胞、幫助病毒復制和調節病毒釋放[7]。目前,針對BVDV p7的研究較少,p7寡聚形成離子孔道的機理尚不明確,抗p7抗體是開展研究工作的必要工具。本試驗通過合成p7蛋白的抗原表位多肽,免疫動物后獲取多克隆抗體,為進一步探索BVDV致病機制提供了有利工具。
1 材料與方法
1.1 材料
1.1.1 細胞、病毒和動物 MDBK細胞購自上海細胞庫;BVDV毒株NADL為國際標準毒株,購自中國藥品監察所;新西蘭大白兔,體重約2.5-3.0 kg,購自陜西西咸新區灃東新城實驗動物養殖場。本實驗通過新疆農業大學實驗動物福利倫理委員會批準(批準號2020040)。
1.1.2 主要試劑和耗材 弗氏完全/不完全佐劑(F5881)和明膠購自Sigma公司;蛋白標準Marker和透析袋10 kD購自Thermo公司;細胞爬片購自NEST公司;Proclin 300和Casein購自北京索萊寶生物公司;HRP標記山羊抗兔IgG(H+L)(SA00001)和 CoraLite594-conjugated Goat Anti-Rabbit IgG(H+L)(SA00013)購自武漢三鷹生物技術有限公司;TMB顯色液、BCA蛋白定量試劑盒、考馬斯亮藍染色、Triton X-100、小牛血清白蛋白BSA、山羊血清購自上海碧云天生物技術有限公司。
1.1.3 主要儀器 酶標儀(111-7)、蛋白電泳槽和電源購自Bio-Rad公司;NanodropTMOne(DS-11)購自DeNovi+公司;恒溫振蕩器(B0101123)購自太倉實驗設備廠;激光共聚焦顯微鏡(Zeiss LSM500)購自德國蔡司公司。
1.2 方法
1.2.1 p7蛋白多肽抗原的設計與合成 使用蛋白二級結構預測平臺JPred(http://www.compbio.dundee.ac.uk/jpred4 /index.html)進行BVDV p7蛋白(GenBank登錄號NC_001461.1)的二級結構分析,預測抗原表位,設計多肽并偶聯KLH載體蛋白形成MSQYGAGEIVMMGN-Cys-KLH,交由吉爾生化(上海)有限公司合成,并由該公司完成RP-HPLC檢測多肽純度和質譜分析多肽分子量。
1.2.2 抗血清的制備 首次免疫前,對2只新西蘭大白兔進行耳緣靜脈采血并分離血清,作為陰性對照。取多肽與等體積的弗氏完全佐劑或弗氏不完全佐劑進行充分乳化;對2只新西蘭大白兔進行背部皮下多點免疫注射,首免使用弗氏完全佐劑,劑量為400 μg/只,濃度為1 mg/mL;此后使用弗氏不完全佐劑乳化多肽,每兩周進行一次免疫;第5次免疫PBS溶解的多肽,劑量為400 μg/只,濃度為2 mg/mL。免疫完成后對新西蘭大白兔進行心臟采血,收集血清。
1.2.3 抗原親和柱制備 將多肽與溴代預活化的交聯瓊脂糖基質Bromohydrin Purose?4 Fast Flow偶聯。先用10倍體積純水將基質中乙醇洗凈,用篩選通過注射器抽干基質;使用偶聯緩沖液(0.2 mol/L Na2CO3和 0.5 mol/L NaCl,pH 10) 溶 解 多 肽 成 7 mg/mL多肽溶液;將活化的基質與多肽溶液按1∶1混合,25℃反應約20 h;使用Nanodrop測定流出液的濃度并計算偶聯效率,大于95%表明多肽偶聯成功;偶聯結束后,用10倍柱體積的去離子水沖液洗去多肽溶液;添加1 mol/L乙醇胺反應8 h,以封閉未反應的活性基團;反應結束后用去離子水沖洗干凈。
1.2.4 抗原特異性抗體純化 將上述抗原親和基質裝填重力柱,上樣前先用5倍柱體積0.1 mol/L,pH 9.5的Na2HPO4·12H2O沖洗親和柱,進行柱平衡;加入抗血清,每次10 mL,棄去流出液;加入5倍柱體積的0.1 mol/L,pH 9.5 Na2HPO4·12H2O淋洗,棄去淋洗液;加入3倍柱體積的0.2 mol/L,pH 4.0 檸檬酸溶液洗脫抗體,收集洗脫液后立即加入1 mol/L,pH 8.0 Tris緩沖液中和;將洗脫抗體溶液使用PBS溶液透析2 h;0.22 μm濾器過濾后,4℃保存備用。
1.2.5 抗體效價檢測 使用pH 9.6碳酸鹽緩沖液將600 ng/孔多肽包被至96孔酶標板,4℃孵育過夜;每孔加入200 μL封閉液(2%明膠、1‰ Proclin 300、5‰ Casein,PBS配制),37℃孵育2 h;對純化后的抗體進行10倍比梯度稀釋,稀釋比依次從1∶10-1∶1×1012;按照每孔100 μL將稀釋后的抗體加入各孔中,37℃孵育1 h;每孔100 μL加入HRP標記羊抗兔 IgG(H+L)(稀釋度 1∶5 000),37℃孵育 30 min;TMB 顯色液顯色10 min后,使用2 mol/L稀硫酸終止顯色反應,酶標儀測定OD450nm值。檢測孔OD450nm值平均值/陰性對照孔OD450nm平均值 > 2.1則判定為陽性;可檢測到陽性信號的抗體最低稀釋倍數為抗體效價。
1.2.6 抗體純度檢測 使用BCA蛋白定量試劑盒對抗體蛋白定量后,加入5×SDS-PAGE loading buffer混勻,100℃加熱10 min使蛋白變性。使用12%SDS-PAGE電泳分離5和10 μg變性蛋白,100 V電泳約2 h;使用考馬斯亮藍染色30 min,脫色液(30%甲醇、10%冰醋酸,PBS配制)脫色2 h。使用GeL Doc2000成像系統獲得圖像,并使用ImageJ分析p7抗體所占灰度值。
1.2.7 間接免疫熒光檢測多克隆抗體特異性 將MDBK細胞接種至鋪有細胞爬片的24孔細胞培養板中,置于細胞培養箱中繼續培養,待細胞匯合度達60%時,接種1 000 TCID50BVDV感染24 h后,棄掉細胞培養液,使用4%多聚甲醛溶液固定15 min,加入0.5% TritonX-100孵育10 min,加入封閉液(0.5%Triton X-100、3% BSA、1%山羊血清,PBS配制)封閉1 h;加入p7多克隆抗體(稀釋度1∶100),4℃孵育過夜;使用CoraLite594-conjugated Goat Anti-Rabbit IgG(H+L)(稀釋度1∶250)室溫孵育1 h,使用抗熒光淬滅封片液封片,使用激光共聚焦顯微鏡觀察熒光陽性情況及定位。
2 結果
2.1 p7蛋白多肽抗原的設計
為合成p7蛋白的多肽抗原,使用JPred預測p7蛋白抗原表位,結果如圖1所示,BVDV p7蛋白共由71個氨基酸所組成,其中綠色為β片層結構,紅色為α螺旋結構,其中B表示氨基酸疏水情況;在25% cut-off溶劑可及性時,p7蛋白的親水區域主要位于該蛋白的氨基端,設計MSQYGAGEIVMMGN多肽,并偶聯KLH載體蛋白以刺激輔助T細胞,進而誘導B細胞免疫反應,合成MSQYGA GEIVMMGNCys-KLH多肽。

圖1 P7蛋白二級結構預測Fig. 1 Prediction of secondary structure of P7 protein
2.2 多肽純度和分子量檢測
使用RP-HPLC檢測多肽的純度,同時使用質譜分析多肽的分子量。結果如圖2和3所示,合成多肽的保留時間為9.810 min,峰面積1 197 903(mAU*s),多肽的純度為 80.19%(1 197 903/1 493 847),合成的多肽分子量為794.2 Da。

圖2 RP-HPLC檢測多肽純度Fig. 2 Determination of polypeptide purity by RP-HPLC
2.3 p7多肽抗體效價檢測

圖3 質譜鑒定多肽分子量Fig. 3 Identification of molecular weight of polypeptide by mass spectrometry
第5次免疫后10 d,心臟采血并收集血清,使用多肽偶聯的抗原純化柱進行親和純化。間接ELISA檢測純化后、PBS對照組和免疫前的抗體反應性。結果如表1所示,經過親和純化后的抗體效價達1∶100 000(PBS對照組的陰性參考值為OD450nm=0.110),與抗原有顯著的反應性,滿足后續實驗需求。
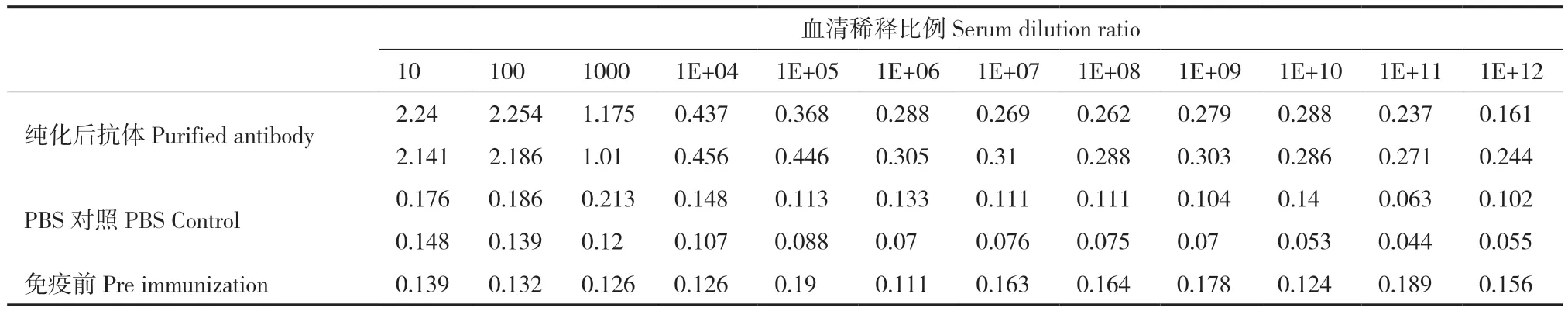
表1 間接ELISA檢測抗體效價Table 1 Detection of antibody titer using indirect ELISA
2.4 SDS-PAGE檢測抗體純化結果
多抗血清通過多肽偶聯的抗原純化柱親和純化后,使用10 kD半透膜對純化血清進行透析,并使用0.22 μm濾器過濾除雜質。SDS-PAGE電泳后使用考馬斯亮藍染色鑒定抗體純度。結果如圖4所示,在55 kD處可見抗體重鏈分子,在25 kD處可見輕鏈分子,使用ImageJ分析p7抗體所占灰度值比為90.2%。表明已純化出兔抗p7多肽IgG抗體。

圖4 SDS-PAGE 檢測抗體純化結果Fig. 4 SDS-PAGE results of antibody purification
2.5 免疫熒光染色鑒定p7多肽多克隆抗體
為進一步鑒定多克隆抗體的反應性,使用免疫熒光染色鑒定BVDV感染MDBK細胞p7蛋白表達及分布情況。結果如圖5所示,BVDV感染MDBK細胞后表達的p7蛋白能與p7多克隆抗體發生特異性反應,可見紅色熒光廣泛分布于BVDV感染MDBK細胞的細胞膜周圍,與p7寡聚在細胞膜上形成離子孔道的蛋白分布一致,而對照組細胞則無法檢測到紅色熒光,表明p7多肽多克隆抗體具有較好的反應性和特異性。黑色箭頭指示細胞核。

圖5 免疫熒光染色測定p7多肽多克隆抗體的反應性Fig. 5 Determination of the reactivity of p7 polypeptide polyclonal antibody by immunofluorescence staining
3 討論
病毒進入宿主后,病毒孔蛋白與宿主細胞中的不同細胞器膜相互作用,借助宿主細胞某些因子組裝成離子通道,介導一些離子(Na+、Ca2+、K+、H+等)運輸而對病毒進入細胞及子代病毒在細胞中成熟產生影響[8-9]。新型丙型肝炎病毒p7離子通道抑制劑BIT225可抑制脂質膜中的p7離子通道活性,并在BVDV感染試驗中具有抗病毒活性,BIT225與重組干擾素α-2b的組合顯示出針對BVDV的協同抗病毒作用,并且通過添加利巴韋林進一步增強了協同作用[10]。用長烷基鏈亞氨基糖衍生物處理人工脂質膜中的p7,對p7的離子通道活性的抑制作用隨著濃度的升高而增強。而這類化合物對BVDV具有強大的抗病毒活性[11-12]。這表明病毒傳播過程中離子通道活性的重要性,以及p7有可能是潛在的藥物靶點。而目前關于p7如何形成離子孔道幫助BVDV釋放?p7蛋白翻譯后如何轉運?需要哪些輔助蛋白的修飾和協作?這些問題的解決都需要構建p7蛋白抗體,否則難以檢測其定位和表達,并探明其形成離子孔道的機制。在本研究中,使用p7蛋白的N端氨基酸,合成多肽,經多次免疫家兔后使用ELISA和免疫熒光染色檢測多肽多克隆抗體具有較好的反應性和特異性。
使用生物信息學平臺分析p7蛋白,發現N端1-14位氨基酸具有較好的親水性。使用化學方法合成多肽并偶聯KLH,免疫新西蘭大白兔制備抗p7多抗血清。ELISA測定經抗原純化柱親和純化后的抗體效價,結果顯示抗血清效價在1∶100 000,說明制備的p7多肽抗體具有較好的反應性。Western blot檢測抗p7多肽抗體結果顯示在55 kD和25 kD處有較為明顯的條帶。免疫熒光染色BVDV感染MDBK細胞可見細胞膜上出現與p7多克隆抗體發生特異性反應,而p7蛋白主要形成離子孔道存在于細胞膜上,綜上結果表明本研究獲得的多肽多克隆抗體可作為抗BVDV p7檢測方法中的具有特異性結合作用的抗體,也為后期深入研究BVDV p7的功能及作用機制奠定了基礎。
4 結論
成功制備兔抗BVDV p7多肽多克隆抗體,使用ELISA和免疫熒光染色發現該抗體具有較好的反應性和特異性,為進一步研究p7形成離子孔道的機理提供了有利工具。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48