超聲波強化木質纖維素酶解的研究進展
2021-11-19 03:07:38胡芳董旭史長偉吳學棟
生物技術通報 2021年10期
胡芳 董旭 史長偉 吳學棟
(1. 齊齊哈爾大學輕工與紡織學院,齊齊哈爾 161006;2. 亞麻加工技術教育部工程研究中心,齊齊哈爾 161006)
能源需求不斷增加,而化石燃料儲量日益減少,使得可再生能源越來越受到關注。生物乙醇是世界上使用最多的生物燃料[1],可與汽油、柴油等原油衍生液體運輸燃料混合使用。作為能源安全和氣候變化問題的共同解決方案,是近年來非常活躍的研究領域。木質纖維素生物質,如農林業廢棄物和部分城市固體廢物,是可再生的、豐富的、廉價的第二代生物乙醇生產原料。第二代生物乙醇的生產涉及4個步驟,包括木質纖維素的預處理、纖維素酶解成可發酵糖、可發酵糖經發酵轉化為乙醇和蒸餾。雖然原料的收購價格比化石燃料具有競爭力,但利用現有技術將木質纖維素轉化為生物乙醇的成本仍然很高。酶法糖化路線約占過程總成本的20%,這一步驟具有創新和降低成本的潛力[2]。近年來,超聲波在酶催化生物轉化中得到了廣泛的應用,具有短時間內強化反應過程、提高產率、降低能耗、更好地利用原料和催化劑等顯著優點[3]。很多研究人員提出利用超聲波強化的策略,有大量文獻報道,超聲波強化的木質纖維素酶解提高了可發酵糖及生物乙醇的產率,并對強化機理進行了不同側面的探索。本文綜述了超聲波強化木質纖維素酶解的應用進展,并著重對超聲波強化酶解的機理進行了分析,旨在為該領域的深入研究提供參考。
1 木質纖維素酶解過程及限制酶解的因素
木質纖維素主要由纖維素、半纖維素和木質素組成,纖維素是葡萄糖單元通過 β-1,4糖苷鍵連接的線型高分子量聚合物。纖維素中存在的羥基形成分子間和分子內氫鍵,其超分子結構由結晶區和無定形區組成,結晶區分子排列規則且緊密,多糖鏈穩定性強,難于降解;而無定形區分子排列無規則且疏松,反應性能較高。半纖維素是高度分支的、由多種糖基構成的不均一聚合物,是無定形的。木質素是高度支化的芳香族聚合物,具有疏水性,由苯丙烷結構單元組成,與半纖維素共價交聯結合,緊密地嵌入纖維素纖維。
1.1 酶解過程
酶催化纖維素水解過程涉及3類酶:內切葡聚糖酶(EC 3.2.1.4)、外切葡聚糖酶(CBH I,EC 3.2.1.176 ;CBH II,EC 3.2.1.91)和 β-D-葡萄糖苷酶(或稱纖維二糖酶)(EC 3.2.1.21),它們以相互作用的方式對纖維素進行水解[4-5]。纖維素酶解為多步非均相反應,不溶性纖維素最初通過內切葡聚糖酶和外切葡聚糖酶的協同作用在固液界面分解。內切葡聚糖酶啟動纖維素水解過程,破壞纖維素鏈無定形區域的內部糖苷鍵,增加纖維素鏈末端數量;然后兩種類型的外切葡聚糖酶從纖維素鏈的還原性末端或非還原性末端分離出纖維二糖。以上過程伴隨著可溶性中間產物的進一步液相水解。最后,纖維二糖酶將纖維二糖轉化為可由酵母菌或細菌發酵成乙醇的D-葡萄糖。
除了以上3種主要的纖維素酶以外,還有許多輔助酶攻擊半纖維素[6],如木聚糖酶切割木聚糖的主鏈;α-阿拉伯呋喃糖苷酶和α-葡萄糖醛酸酶分別去除阿拉伯糖和4-O-甲基葡萄糖醛酸取代基;阿魏酰基酯酶攻擊阿拉伯糖取代基和阿魏酸之間的酯鍵。提高酶用量,可以獲得較高的糖產量,但同時導致成本增加,為了避免這一點,可以利用纖維素酶與其他酶之間的協同作用[7-8]。
1.2 限制酶解的因素
木質纖維素不易被纖維素酶水解的主要原因是:(1)與纖維素共存的木質素和半纖維素也是木質纖維素生物質酶解中的屏障,限制酶對纖維素的可及性[6,9];(2)微晶纖維素的可及性低,阻礙了纖維素酶的有效作用[6,9];(3)非均相反應中的傳質阻力[10-11];(4)產物抑制作用[6,12]。
木質纖維素轉化為生物乙醇的第一個步驟:預處理,采用機械粉碎、稀酸預處理、堿預處理、水熱法、氧化法、氨爆破、蒸汽爆破、有機溶劑預處理、離子液體預處理和生物法等[13],可提高底物比表面積、降低纖維素結晶度、破壞木質素和半纖維素的屏障,從而促進纖維素的水解反應[14]。雖然預處理去除了部分木質素和半纖維素,但為了提高酶解的總糖收率,總會有部分木質素和半纖維素保留在原料中,并影響酶解過程[15-16]。木質纖維素生物質的酶轉化涉及從液體介質向固體底物表面的傳質,由于這種傳質是由擴散控制的,所以酶解的總反應速率由酶大分子的擴散速率控制。一般來說,大型三維酶分子的擴散速率很低,這大大阻礙了水解反應的整體速率。使用纖維素酶的一個重要限制是,可溶性產物(纖維二糖和葡萄糖)的抑制作用降低了反應速率。減少產物抑制作用的措施,包括水解過程中補充β-D-葡萄糖苷酶、添加纖維二糖脫氫酶、或采用同步糖化發酵(simultaneous saccharification and fermentation,SSF)工藝[6]。
2 超聲波強化木質纖維素酶解的應用
超聲波強化木質纖維素酶解,主要由以下幾個途徑實現:超聲波預處理酶-底物、超聲波強化糖化、超聲波強化SSF、超聲波強化產酶過程,其中文獻報道最多的是超聲波強化糖化過程,如表1所示。

表1 超聲波強化木質纖維素酶解的應用Table 1 Applications of ultrasound intensification for enzymatic hydrolysis of lignocellulose
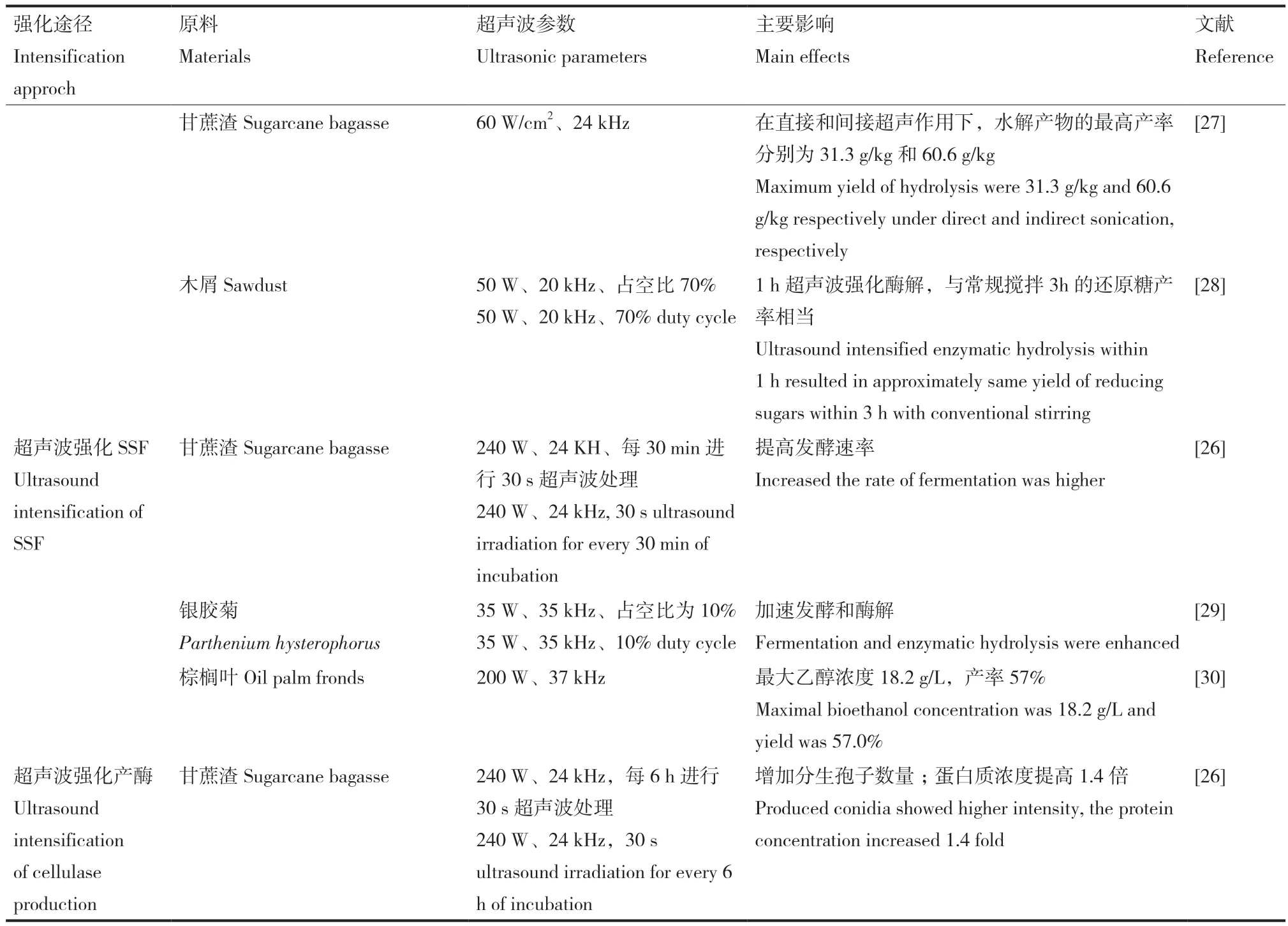
表1續表 Continued
Silvello等[1]利用超聲波對酶-底物進行預處理,然后在50℃、300 r/min條件下水解24 h。超聲波預處理的最佳條件為150.7 W/cm2、20 kHz、330 s和25℃,纖維素酶活性提高了42.35%,還原糖濃度達到對照樣的1.9倍。Subhedar 等[17]利用廢報紙酶解生產可發酵糖,并與常規酶解進行了比較。在相同的底物濃度下,常規酶解的條件為:酶用量0.14%(W/V)、水解72 h,還原糖得率11.569 g/L;以超聲波輔助酶解,酶用量0.08%(W/V)、水解6.5 h、還原糖產量27.6 g/L。超聲波強化使酶解時間大大縮短,酶用量顯著減少,還原糖的釋放濃度達到對照樣的2.4倍。Subhedar等[18]以花生殼、椰子殼和開心果殼3種木質纖維素生物質為原料,超聲波輔助堿預處理后進行常規酶解和超聲波輔助酶解,得到了相似的結果。Borah等[23]利用超聲波促進雜草酶解過程,超聲波代替機械攪拌可以使水解動力學提高近10倍,機械攪拌120 h與超聲處理10 h獲得了相同的糖產量。Gasparotto等[27]研究了固態發酵產纖維素酶在直接和間接超聲波作用下對未經處理蔗渣的酶解,認為間接超聲波是更有前途的輔助酶反應技術。
研究者們還進行了超聲波強化SSF工藝[26,29-30]和超聲波強化微生物生產纖維素酶[26]的研究。SSF工藝具有操作條件溫和、發酵罐單一、水解和發酵兩步結合的優點。在SSF過程中,發酵微生物消耗生物質中纖維素酶解釋放的己糖,降低糖化產物對酶的抑制作用,也降低了侵入性微生物污染的可能性。與傳統的兩步法相比,SSF工藝更加節能。然而,SSF工藝的一個主要問題是酶解的最佳溫度(50-55℃)與發酵的最佳溫度(30-35℃)之間的不同,因此SSF過程的主要限制是兩個獨立過程的最佳條件的折衷。Ofori-Boateng等[30]以棕櫚葉為原料,在超聲波-有機溶劑預處理后,在200 W、37 kHz、培養時間5 h、40℃、pH值5、酵母濃度15 g/L、固體負荷10%(W/V)條件下進行SSF工藝,得到最大乙醇濃度18.2 g/L,產率57%。在相同的條件下,進行無超聲波處理SSF工藝,生物乙醇濃度(3.1 g/L)和產率(14.0%)較低,常規SSF工藝需要超過24 h才能獲得高生物乙醇產量,而超聲波輔助SSF工藝在短時間和高固體負載下可顯著提高生物乙醇的產量。Singh等[29]在超聲波輔助下,利用銀膠菊,采用SSF生產生物乙醇,研究發現,在30℃下,發酵時間18.3 h后,乙醇濃度提高到15.62 g/L,乙醇產率為0.4 g/g預處理生物質;無超聲波SSF實驗在發酵54 h后,乙醇濃度最高為10.57 g/L,乙醇產量為0.27 g/g預處理生物質。超聲波處理對SSF過程最顯著的影響是縮短了發酵時間。Velmurugan等[26]研究了超聲波對甘蔗渣培養里氏木霉產纖維素酶、酶解及SSF生產乙醇的影響。甘蔗渣培養里氏木霉分生孢子的萌發強度明顯提高,超聲波處理通過增強細胞分裂來增加分生孢子數量。超聲波強化酶解的葡萄糖濃度為23.5 g/L,而常規酶解為19.7 g/L。240 W超聲下,水解液中乙酸和糠醛的最大濃度分別為399 mg/L和109 mg/L,遠低于抑制水平。在SSF實驗中,測定了超聲波輔助水解條件與釀酒酵母生長的相容性,在400 W下,超聲波刺穿了細胞;而240 W的超聲波不會使細胞破裂,從而提供了超聲波對發酵的積極作用。超聲波-SSF發酵速率比常規SSF高,且大部分糖在發酵30 h后轉化為乙醇,最大濃度為11 g/L。另一方面,沒有超聲波的SSF實驗顯示,即使在發酵36 h后,乙醇濃度仍逐漸增加,獲得的最大濃度為9 g/L。Silvana等[31]采用超聲波強化球孢白僵菌產β-1,3-葡聚糖酶,在24 h的發酵過程中,以195 W、24 kHz、50%占空比處理5 min,使β-1,3-葡聚糖酶的產量增加了46%。在掃描電子顯微鏡(SEM)照片中觀察到菌絲體的破碎,證實了酶更好地釋放到細胞外。Shaheen等[32]以80 mW/cm2、1.5 MHz、占空比20% 的超聲波強化里氏木霉產纖維素酶過程,超聲波輻照每天12次、每次5 min、發酵6 d,纖維素酶活性提高22%。未接受超聲波處理的樣品,菌絲壁圓而光滑;而超聲波處理后的里氏木霉,菌絲收縮、菌絲壁上有許多裂紋和開口,這些裂縫和開口可能促進纖維素酶的釋放。
3 超聲波強化木質纖維素酶解的機理
基于不同的實驗方法和側重點,關于超聲波強化木質纖維素酶解機理,研究者們提出了多種解釋,現歸納如下。
3.1 促進非均相系統的擴散和傳質
在非均相系統中,空化泡發生非對稱坍塌,當氣泡在固體表面附近破裂時,會產生高壓力/高速液體射流,這些射流可以清潔固體表面,并通過中間體的脫附而活化固體催化劑,還可以通過擾動界面邊界層來增加傳質。固體表面的不對稱塌陷也會導致底物碎片和表面粗糙[33]。因此,超聲波可以增加底物比表面積、微觀/分子水平的混合,從而增加傳質。
Yachmenev等[11]和 Easson 等[34]認為木質纖維素酶促反應可以總結為以下4個階段(圖1),其中2個階段(1和4)涉及酶大分子和酶反應產物到達和離開固體底物表面的運輸,這兩個階段都受擴散控制,因此酶水解的總反應速率由酶大分子的擴散速率控制。但酶蛋白大分子的擴散速率很低,大大阻礙了水解反應的整體速率。非均相系統(如酶溶液-木質纖維素生物質)中空化泡的潰滅與均相系統不同,由于表面對液體流動提供阻力,表面上或表面附近的空化泡是非對稱的。其結果是大量液體從氣泡的另一側(遠離底物表面)涌出,形成強大的液體射流(約500 m/s)并作用于表面。同樣重要的是,空化泡的快速潰滅在氣泡周圍的液體中產生顯著的剪切力,并因此產生強烈的機械攪拌效應。在非均相系統中,空化效應比均相系統中的大幾百倍。這種理想的攪拌機制,發生在固液界面的緊密邊界層,大大提高了酶大分子對底物表面的供應。由于酶促反應的特殊性,超聲波在促進擴散和傳質的同時,還發揮與之相關的以下積極影響:
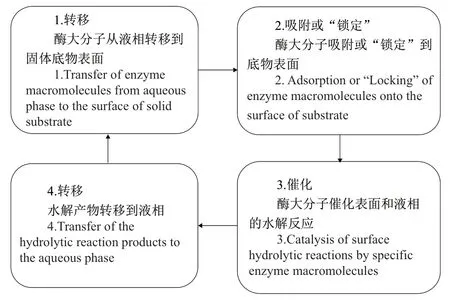
圖1 固體底物酶促反應階段Fig. 1 Schematic diagram of the general stages of an enzymatic reaction on a solid substrate
(1)超聲波不僅增大酶/底物的親和力,而且提高了酶/底物復合物轉化為產物的速度。酶/底物的親和力增大是運輸更快和酶與底物的相互作用增強的結果,這歸因于反應混合物中由超聲波空化產生的微湍流和強烈的混合作用。Nadar等[3]、Sulaiman等[10]和 Subheder等[35]利 用 Michaelis-Menten方程變換,繪制了Lineweaver-Burk曲線,得到Michaelis常數(Km)和當酶被底物飽和時的最大反應速率(vmax)。結果表明,與未處理酶相比,超聲波處理后纖維素酶Km降低,而vmax增加。Silvell等[36]得到了相似的結果,在超聲波處理下,Km比對照組降低23%,vmax提高了21%。Km表示酶與底物之間的親和力,超聲波處理下Km的降低,是由于更快的傳質和酶與底物相互作用增加。vmax的增加表明,在超聲波場下,反應物向酶活性部位大量轉移,酶 - 底物復合物快速裂解[23,34]。(2)促進可溶性產物,即纖維二糖和葡萄糖,從纖維素表面被帶走并在介質中被稀釋,降低與酶活性位點結合的可能性,從而降低產物抑制程度。Borah 等[23]和Singh等[29]采用Holtzapled等[37]提出的一級產物抑制模型(又稱HCH-1酶解纖維素模型),研究了超聲波對銀膠菊和4種雜草的酶解強化過程,由實驗結果得到HCH-1模型參數值,其中產物結合常數降低,證實了超聲波強化減少了酶解過程中的產物抑制作用。(3)酶大分子通常不是完全剛性的,而是在溶液中具有某種構象柔性,因此,由超聲波作用引起的固液界面邊界層的劇烈攪拌,將有助于酶大分子更容易“合適”地定位在底物上[11]。(4)微湍流破壞酶分子聚集體,使得酶活性位點更易于進行反應,從而提高活力[3,36]。
3.2 增加纖維素底物的可及性
微氣泡的快速崩解產生剪切力,有助于將木質纖維素材料分解成更細小的顆粒,從而顯著增加酶攻擊的表面積。由空化氣泡的崩潰所產生的機械沖擊,為酶促作用打開了固體底物的表面[9,25,38]。
Adewuyi 等[9]利用BET比表面積測定法和XRD(X-ray diffraction,X射線衍射)技術對天然和處理后的纖維素進行表征,結果表明,天然纖維素的比表面積為2.18 m2/g,但在僅酶存在的水解反應后增加到2.68 m2/g,在超聲波/酶存在下增加到4.32 m2/g,分別增加了23%和近100%。經過酶和(或)超聲波處理后,纖維素結晶度有顯著變化,從天然纖維素的75%降低到酶處理纖維素的70.37%、超聲波-酶處理纖維素的66.15%。因此,在超聲波存在下纖維素比表面積增加,同時纖維素粉末結晶度略有降低,酶和超聲波的協同作用使纖維素的微觀結構發生了變化,促進了纖維素水解。
3.3 影響酶蛋白空間結構,提高酶活性
酶蛋白的空間結構對其催化效率有至關重要的作用,研究者們利用內源熒光光譜法和圓二色(circular dichroism,CD)光譜法分析了超聲波對酶分子空間結構的影響。
內源性熒光主要與芳香族氨基酸殘基有關,特別是色氨酸殘基。Borah等[23]發現,機械攪拌和超聲處理均能使熒光強度降低,但超聲波的作用更明顯,最佳熒光發射波長(348 nm)沒有發生任何紅移或藍移。這種變化是由于空化產生的強烈微對流使得蛋白質分子間疏水相互作用被破壞,導致蛋白質分子去折疊,纖維素酶分子疏水性氨基酸基團和酶分子內部結構暴露,導致活性增強。Subhedar等[35]得出了相同的結論。Wang等[2]發現,纖維素酶分子的內源熒光強度隨著超聲波功率、超聲波頻率和超聲波處理時間的增加而降低。
利用CD光譜法可以分析酶蛋白二級結構的變化,Wang等[2]發現,24 kHz、15 W、10 min的超聲波處理使纖維素酶的構象發生變化,部分α-螺旋發生變形,而無規卷曲含量增加,相應地,酶活力提高到59.58 μ/mL;而在29 kHz、50 W、30 min條件下,纖維素酶α-螺旋含量增加且無規卷曲含量減少,纖維素酶的酶活力下降到31.62 μ/mL。Subhedar等[35]研究發現,在最佳超聲條件下,與未處理酶相比,α-螺旋含量下降了12.4%,無規卷曲增加了29.6%。Fan等[39]研究了超聲波處理對β-D-葡萄糖苷酶二級結構的影響,得到了相同的結論。還有研究者發現,超聲波對蛋白質構象的影響同樣適用于固定化酶[2,40],Ladole 等[40]將纖維素酶固定在磁性納米粒子上,在24 kHz、6 W、6 min條件下,酶催化活性提高近3.6倍。CD光譜分析表明,超聲波處理使β-片層和無規卷曲分別增加了6.23%和0.69%,而α-螺旋和β-轉角的含量則減少了1.8%和2.11%。CD光譜研究證實,超聲波處理使得酶蛋白的有序結構減少,而無序結構增加,有利于暴露酶蛋白內部的催化位點,纖維素酶顯示出更大的靈活性,使得纖維素酶易于與底物相結合,有助于提高催化效率。
但超聲波處理對酶活性也存在負面影響,超聲波空化產生高活性中間體,如H·、OH·、H2O2、HO2和H2,可能與底物結合域或催化域的氨基酸殘基發生反應[41]。有研究者提出[6],活性中間體與蛋白質骨架作用,會導致酶的聚集,從而阻礙活性位點,同時也降低蛋白質的穩定性。此外,超聲波處理產生的剪切力在酶失活中也有相當大的作用[37,41]。但Nguyen等[41]和Sun等[42]的研究證實,超聲波并沒有引起纖維素酶分子的劇烈損傷,其一級結構沒有降解或改變。
超聲波處理使纖維素酶活性增加或降低,與超聲波參數設置密切相關。總的來說,低強度超聲波將酶分子聚集體分解成更小的碎片,暴露更多的活性位點,有助于酶活性的增加;同時,高強度超聲波結合長時間處理導致自由基反應,可使酶失活[43-45]。但不同的文獻中,引起酶活性下降的超聲波參數是不同的,這與超聲波發生器的類型、底物的性質、其他參數的設定等有關。Su等[19]的研究發現,在高強度(>120 W)下,纖維素酶活性在50℃時明顯下降;然而,在所有輸出功率(0-500 W)下,37℃時的纖維素酶活性都是穩定的。Szabó等[43]發現,在40%振幅(40 kHz,最大功率500 W)下,酶活性在65 min內損失了12%。當振幅增加到60%和80%時,酶活性下降更為顯著,65 min后酶的活性分別只有原來的80%和75%。Silva等[46]發現,在20 kHz和50 W/cm2的超聲波條件下,β-D-葡萄糖苷酶活性提高1.5倍。Chen等[47]發現,在968 W/cm2和10 min時,β-D-葡萄糖苷酶活性下降了84.71%。Sun等[42]發現低溫度(20-45℃)、低超聲波強度(<181.53 W/cm2)和短處理時間(<15 min)導致了 β-D-葡萄糖苷酶的激活,而高溫度(45-60℃)、高超聲波強度(>181.53 W/cm2)和長處理時間(>15 min)導致其抑制。
4 關于超聲波強化木質纖維素酶解的不同觀點
4.1 超聲波處理是否會導致纖維素酶活性下降
如前所述,很多文獻報道了適當條件下的超聲波處理改變纖維素酶的空間結構,使纖維素酶活性提高,但高強度超聲波會導致纖維素酶失活,對此,有研究者提出了不同的觀點。Adewuyi等[9]在611 khz、104 w、50℃和pH 5.2條件下得到最佳的纖維素水解,葡萄糖產率為41.4%,延長時間(40-50 h)可獲得高達70%的葡萄糖產率。在大約30 h后觀察到水解速率降低,可能是由于除了產物、底物和吸附抑制之外,易水解的非晶纖維素的減少。Adewuyi等[9]提出,高活性中間產物的形成,特別是在高超聲波頻率下的形成,對溶解酶大分子的長期催化/活性沒有任何顯著影響。一個可能解釋是,在通常的低酶濃度下,酶相對于大量溶劑分子(如水)的比率極低,酶大分子被滯留在空化氣泡中并遇到由潰滅的氣泡所產生的高活性中間產物的可能性很低,因此大多數酶對超聲波的抵抗力非常高,以至于產生廣泛酶活性惡化所需的超聲波強度可能太高而無法達到。Yachmenev等[11]也支持了這一觀點。但在這些報道中,并沒有測定纖維素酶的活性,研究者認為,利用超聲波和酶對纖維素水解的改善歸因于超聲波作用下纖維素的表面積增加和結晶度降低。另有研究者發現[19,43],雖然高強度/長時間超聲波處理仍然使酶催化水解效率提高,但經測定發現纖維素酶活性確實下降了,其原因是超聲波對非均相纖維素-纖維素酶反應的積極影響能夠克服其對酶活性的負面影響。因此超聲波強化木質纖維素酶解各種機理的相對貢獻,以及在酶解過程不同階段的差異,有待于進一步研究。
4.2 超聲波反應器的類型和參數設置對研究結果有重要影響
超聲波強化木質纖維素酶解的實驗室研究中,常常采用槽式或變幅桿式超聲波反應器,個別研究中采用了新型多頻超聲反應器[9]和可提供均勻可控超聲波的六邊形超聲波反應器[11,34],對于變幅桿式超聲波反應器,變幅桿的位置和直徑是影響超聲波作用的重要參數,不同類型超聲波反應器和同種類型不同參數設置的超聲波反應器,其研究結果難以比較。Gasparotto等[27]提出間接超聲波是最有前途的輔助酶反應技術,在實驗中,直接超聲波和間接超聲波均由變幅桿式超聲波反應器提供,但另有文獻[3]中提出的直接超聲波和間接超聲波被分別定義為由槽式和變幅桿式超聲波反應器提供。利用理論和實驗的方法,對超聲波反應器空化行為進行定量描述和評價,是對超聲波反應器類型和參數設置進行對比的有效途徑。超聲波反應器的設計不僅對實驗室研究結果有重要影響,更是決定超聲波強化木質纖維素酶解研究放大和實現工業化的關鍵因素。
5 超聲波強化木質纖維素酶解的經濟效益評價
木質纖維素生物質的酶解成本高、效率低,是阻礙第二代生物乙醇成本競爭性生產的主要原因。盡管近年來纖維素酶的生產成本大幅度降低,但木質纖維素糖化過程仍然是一個昂貴而緩慢的步驟。超聲波強化木質纖維素酶解可縮短反應時間、減少纖維素酶的用量,但同時必須考慮設備的投資和維護,以及超聲波處理是一個能量密集過程這一現實。對這一過程進行技術經濟分析是重要的研究課題,但僅有個別文獻進行了報道[47],以舊報紙、辦公廢紙為原料的超聲波輔助酶解,附加的超聲波能量為5.2-10.4 MJ,總產糖量增加,額外能量輸出為15-160 kJ,超聲波輔助酶解對總成本有消極的影響。利用現有的超聲波技術,對纖維素生物質進行預處理和熱化學/生物化學反應進行強化,額外的超聲波處理消耗的能量大得多,而由于糖產量提高而增加的能量輸出僅為消耗能量的一小部分,在超聲波輔助糖化的初期,酶效率的提高可能并不顯著,原因在于木質細胞結構非常頑固,同時,懸浮狀態下的微米/納米級木質纖維素顆粒的高體積密度,導致超聲波能量嚴重衰減。為此,需要進一步研究超聲波反應器的結構和設計、超聲波參數和操作對特定反應的適應性;超聲波與其他強化處理方法結合,特別是大規模處理高固體濃度的生物質原料;進行超聲輔助反應的動力學和機理研究,應分析不同反應所需的活化能水平,以找到超聲波強化與化學活化的最佳組合。
6 總結與展望
僅用酶法進行木質纖維素生物質水解是相當緩慢的,超聲波可參與酶-底物預處理、糖化、SSF工藝和產酶過程,以多種方式強化木質纖維素的生物轉化,對酶促反應提供明顯的強化效果,可提高可發酵糖以及生物乙醇產量、增大酶解速率、減少酶解反應時間和降低酶用量。本文總結了促進非均相系統的擴散和傳質,增加纖維素底物的可及性,改變酶蛋白空間結構、提高酶活性等超聲波強化酶解的機理,但超聲波強化木質纖維素酶解這一課題仍有很大的深入研究的空間,包括:
(1)超聲波強化與其他酶解改進措施的協同作用。為了提高木質纖維素酶解效率,可采用多種措施,如在酶解體系中加入適量的水溶性木質素、非離子表面活性劑,可有效減少底物木質素對纖維素酶的無效吸附[48-49];添加纖維二糖脫氫酶、擴展蛋白和膨脹素,可顯著提高纖維素糖化效率[50];Mg2+、Co2+、Ca2+和阿魏酸、對香豆酸等對纖維素酶具有激活作用[4]。超聲波強化與以上酶解改進措施的協同作用的研究將為木質纖維素生物乙醇的生產提供更多的機遇。
(2)超聲波空化的物理/化學與酶解的生物化學之間的確切聯系尚未建立。有必要研究在不同實驗條件下的超聲波效應,建立數學模型,量化可變參數對超聲波整體效應的貢獻,從而預測過程效率的變化。需要了解酶結構變化與超聲波處理的關系,了解酶反應表觀動力學變化與酶活性位點的數量和分布之間的聯系。
(3)超聲波強化木質纖維素酶解操作參數的綜合優化。關鍵參數包括包超聲波發生器的結構、功率、頻率、處理時間,以及酶解的一般參數,如溫度、酶用量、酶-底物比率等。此外,還應考慮其他因素,如靜壓、充氣操作和溶劑特性。在分析實際強化機理的基礎上,對超聲波強化木質纖維素酶解操作參數進行優化。