牡蠣胰島素相關多肽3 克隆表達及純化
2021-11-27 00:17:48楊丙曄陳仲巍
中國醫藥導報 2021年30期
楊丙曄 李 瑩 陳仲巍
廈門醫學院廈門市海洋藥用天然產物資源重點實驗室,福建廈門 361000
牡蠣屬于軟體動物門、雙殼綱、珍珠貝目、牡蠣科,是一類非常重要的海洋經濟物種,其蛋白含量豐富,肉質鮮美,是人們非常喜愛的海洋食物之一,素有“海中牛奶”的美稱[1-2]。牡蠣地理分布廣泛,沿海的國家、地區幾乎都有牡蠣的分布,且生長迅速,產量高和經濟效益好,是世界各國重要的海水養殖種類[3]。中國是世界第一海水養殖大國,牡蠣的養殖產量也位居世界首位[4]。到2007 年,牡蠣養殖產量達到海洋養殖總產量的1/3,是中國海水養殖產量最大的種類。但近幾年牡蠣的養殖也出現了種質退化的問題[5],其中主要的問題就是生長慢、成熟個體小型化、品質下降[6-7],解決這一問題除了培育新品種,近年來有研究學者發現體外添加胰島素樣生長因子可以促進雙殼類軟件生物的生長[8]。
在哺乳動物中,胰島素超家族在生長發育、代謝和壽命等方面具有重要的調節控制作用[9-11]。胰島素超家族主要包括四類:胰島素、胰島素樣生長因子(insulin growth factor,IGF)、松弛素和胰島素相關多肽(insulin-related polypeptides,IRP)。胰島素超家族相對比較保守,研究發現胰島素超家族對生物體的多種生理過程都有重要調節作用[12]。IGF 主要在脊椎動物中起調節作用[13],IRP 主要在無脊椎動物中被發現起作用。利用大腸埃希菌表達脊椎動物的IGF 已經有很多報道[14-15],使用大腸埃希菌表達海洋無脊椎動物的胰島素相關多肽還未見報道。本研究利用原核表達系統,表達純化牡蠣IRP3 蛋白,為IRP3 蛋白的抗體制備和體外作為生長因子添加劑的應用打下基礎。
1 材料與方法
1.1 實驗材料
質粒提取試劑盒(Plasmid DNA extraction kit,DP1103)和DNA 膠回收試劑盒(DNA gel Extraction Kit,DP209)購自天根生化科技有限公司;T4 DNA 連接酶(T4 DNA Ligase,M0202L)和Taq DNA 聚合酶(Taq DNA Ploymerase,C11304-011)來自寶生物工程有限公司;ECL 顯色試劑盒(Enhanced chemiluminescent detection reagent kit,SW2020)購自泰德生物科技有限公司;質粒pET30a 和菌株E.coli BL21 實驗室保存。
1.2 IRP3 基因克隆構建和篩選
利用全基因合成技術合成IRP3 基因,并在IRP3基因兩端加入限制性酶切位點Nde Ⅰ和Hind Ⅲ,利用雙酶切和連接技術將IRP3 基因克隆構建到載體pET30a 中,然后轉化Top10 感受態細胞,在含有卡那霉素(Kanarnycin,Kan)的LB 平板培養基上篩選長出的陽性菌落,分別利用雙酶切和測序的方法確認構建載體的準確性,提取陽性質粒轉化BL21(DE3)表達菌株。
1.3 IRP3 蛋白表達及鑒定
挑取BL21(DE3)陽性表達菌株單克隆,接種到4 ml的含有Kan LB 液體培養基中,37℃培養4 h,菌液OD600 達到0.8 時,向菌液中加入IPTG 使之終濃度為0.1 mmol/L,之后分別于15℃和37℃誘導表達培養。收集誘導后的培養液,12 000 r/min 離心5 min,去除上清液,加入PBS 液重懸菌液沉淀,最后加入上樣緩沖液,于沸水浴中加熱樣品10 min,將樣品12 000 r/min離心10 min,將上清液加入SDS-PAGE 凝膠中,電泳分離蛋白。分離好蛋白的凝膠考馬斯亮藍染色液染色,隨后加入脫色液脫色直至凝膠沒有蛋白的地方透明清晰。
1.4 IRP3 蛋白純化
將所有菌液收集,加入含有20 mmol/L PBS(pH7.2),300 mmol/L NaCl,20 mmol/L Imidazole(含1% Triton X-100,1 μg/mL Pepstatin A,1 μg/ml Leupeptin)裂解液,利用超聲儀器(JY92-IdII,寧波新芝生物科技股份有限公司)進行超聲破碎菌液,離心機離心收集上清液,利用Ni-IDA 親和層析柱純化表達的蛋白,利用SDS-PAGE 對純化蛋白進行檢測分析。用含有20 mmol/L PBS(pH7.2),300 mmol/L NaCl,8 mol/L Urea,20 mmol/L Imidazole 的緩沖液溶解包涵體,利用Ni-IDA親和層析柱純化包涵體裂解上清蛋白,利用SDS-PAGE對純化蛋白進行檢測分析。
1.5 IRP3 蛋白的鑒定
利用SDS-PAGE 電泳和Western blot 技術,以BSA 作為對照,對純化透析后的樣品分別進行鑒定分析,SDS-PAGE 電泳后,一塊膠用于考馬斯亮藍的染色,一塊膠繼續以恒電壓100 V 轉移60 min 進行轉膜操作,將電泳凝膠中的蛋白轉移到PVDF 膜上,然后經過膜封閉和6HIS 抗體的孵育結合,最后通過ECL 試劑盒顯色,用儀器熒光觀察拍照。
2 結果
2.1 IRP3 基因質粒克隆的檢測結果
IRP3 基因序列長度為1818 bp,開放閱讀框長度為483 bp,編碼161 個氨基酸,NCBI 序列號為XM_011456859.3。瓊脂糖凝膠電泳結果發現經過雙酶切的樣品泳道只有兩個條帶(圖1),大小與pET-30a 和IRP3目的基因片段大小相符。顯示pET-30a 和IRP3目的基因片段已成功進行了克隆構建。

圖1 雙酶切瓊脂糖凝膠電泳
2.2 IRP3 蛋白誘導表達結果
SDS-PAGE 電泳結果發現誘導樣品2 和3 都有目的蛋白表達,且表達量基本一致。但15℃過夜表達樣品的蛋白條帶更少,提示菌體內表達的雜蛋白相對較少(圖2),15℃過夜培養表達更優。

圖2 IRP3 蛋白的誘導表達
2.3 IRP3 蛋白的純化
SDS-PAGE 電泳結果顯示菌液普通裂解上清樣品沒有發現目的蛋白IRP3(圖3)。包涵體溶解離心的上清及上清液純化后的蛋白含有大量的目的蛋白IRP3,提示利用鎳柱親和層析的方法從包涵體裂解液中獲得了較純的IRP3 融合(圖4)。
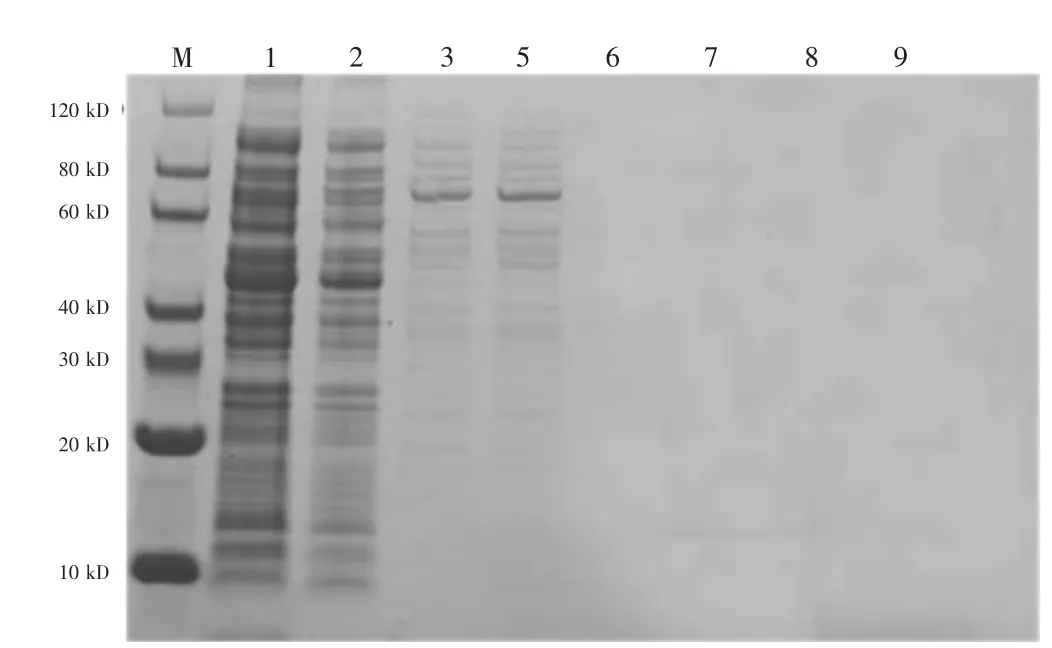
圖3 上清純化樣品電泳

圖4 包涵體純化樣品電泳分析
2.4 IRP3 純化蛋白的鑒定分析
SDS-PAGE 電泳和Western blot 結果顯示純化的IRP3 蛋白樣品條帶單一且分子量與電泳指示一致(圖5),說明純化獲得蛋白樣品確實是IRP3 蛋白,與BSA 比較發現純化的IRP3 蛋白樣品純度更高。

圖5 IRP3 純化樣品的SDS-PAGE 和Western blot 分析
3 討論
經過研究發現IRP 在貝類中也廣泛存在,貝類中的第一個類胰島素家族基因是在腹足綱的靜水椎實螺(Lymnaea stagnalis)中發現的,并有7 個貝類IRP 基因被發現且在神經節中表達[16]。至今已在多種無脊椎動物中證實轉錄水平的調控與環境應激適應力有關[17]。目前普遍認為IRP 基因參與貝類的生長、發育和繁殖調控過程[18]。同時在長牡蠣中發現IRP 基因表達量增長期與長牡蠣的快速生長期和繁殖期時間是一致的[19-20],這提示了IRP 基因可能對長牡蠣的生長和繁殖調控有重要作用[21]。
無脊椎動物中IRP 通過IRP 受體/胰島素受體,激活MAPK 和PI3K/PKB 信號通路,調節生物的生長發育、新陳代謝、免疫和繁殖等各種生理過程。IRP 與胰島素的結構有很大相似性,包括了信號肽,和A、B、C 三鏈。在蛋白表達修飾工程中,C 鏈會被剪切,這樣由A 和B 鏈形成成熟結構,在生理過程調節中,一個存在于A 鏈內部的二硫鍵和A 鏈和B 鏈之間的二硫鍵起了至關重要的作用[22],并且A 鏈和B 鏈內形成二硫鍵的半胱氨酸在生物進化過程中高度保守[23]。胰島素相關多肽在無脊椎動物中廣泛存在,其調控網絡繁多且比較復雜,一般認為是由PI3K/PKB 通路和MAPK通路這兩條信號通路組成,在這些通路組成上的各種因子會進行各種轉錄和翻譯后修飾水平的調控[24-25],由此調節無脊椎動物各種生長發育及新陳代謝生理過程[26]。本研究成功克隆獲得含有IRP3-pET30a 質粒的BL21 表達菌株,從包涵體裂解上清液中獲得了純化的IRP3/HIS 融合蛋白,為IRP3 蛋白的抗體制備和作為體外生長因子添加劑的應用打下基礎。
猜你喜歡
人人健康(2023年26期)2023-12-07 03:55:46
中等數學(2022年2期)2022-06-05 07:10:50
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:48
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
中國生殖健康(2019年2期)2019-08-23 08:12:10
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
數學小靈通·3-4年級(2017年6期)2017-06-22 11:28:50
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
