應用釀酒酵母孢子壁衍生材料向非巨噬細胞HEK293T運送微米顆粒
2021-11-29 10:45:12宋超群楊巖李鳳劉國玉高曉冬中西秀樹
食品與發酵工業 2021年22期
宋超群,楊巖,李鳳,劉國玉,高曉冬,中西秀樹
(江南大學 生物工程學院,江蘇 無錫,214122)
釀酒酵母(Saccharomycescerevisiae)在營養缺陷或極端情況下,雙倍體的酵母會進入減數分裂過程,最終在二倍體細胞中產生4個單倍體孢子,從而對抗外部環境[1]。單個釀酒酵母孢子的直徑在2 μm左右,可用作大分子生物材料[2-4]。然而現有報道中,尚未成功解析釀酒酵母孢子壁的結構[5-8]。本實驗室主要對釀酒酵母孢子進行研究,并且通過高濃度鹽洗脫釀酒酵母孢子表面,在洗脫液中發現了RNA[9]。
HEK293T細胞來源于人胚胎腎細胞HEK293細胞的優化,是生物分子研究中常用到的細胞之一,由于HEK293T細胞的轉染效率高,已成為廣大研究者研究基因功能的一個首選細胞株[10]。HEK293T細胞不同于巨噬細胞,沒有明顯的吞噬作用[11-13]。實驗室發現HEK293T可以吞噬釀酒酵母孢子,且內化效率較高[9]。本研究使用購自Sigma公司的聚苯乙烯乳膠顆粒(latex beads amine-modified polystyrene, beads),其直徑為2 μm。HEK293T細胞不能內化beads,但是beads通過靜電結合釀酒酵母孢子表面分子后,可以被HEK293T細胞內化。
因此,本文著力研究釀酒酵母孢子表面分子與beads交聯后,能否出現吞噬作用,以及影響吞噬效率的因素[14-15]。本研究為誘導非巨噬細胞內化大顆粒機制研究、釀酒酵母孢子表面的特性研究奠定了基礎[16-17],也為未來釀酒酵母孢子的醫用提供了理論基礎。
1 材料與方法
1.1 實驗材料
1.1.1 菌株
釀酒酵母菌株是以SK-1背景下的二倍體菌株AN120作為野生型(wild type,WT)菌株;大腸桿菌DH5α。本研究所用菌株均為本實驗室保存。
1.1.2 酶及試劑
Lyso-Tracker Red,碧云天生物技術公司;RNase、Protease、Loading buffer,寶生物工程(大連)有限公司(TaKaRa);PBS緩沖液、Agrose M、酵母提取物、胰蛋白胨、腺嘌呤、乙酸鉀、葡萄糖、氯化鈉、山梨醇,生工生物工程有限公司;Liticase溶菌酶、山梨醇、RNAiso Plus、beads,Sigma;DNA marker 1kb plus,北京全式金生物技術有限公司;氯仿、異丙醇、戊二醛,中國醫藥集團有限公司。
1.1.3 培養基及其他溶液配制
LB培養基(g/L):酵母提取物5,胰蛋白胨10, NaCl 5,瓊脂粉20(固體培養基)。
YPAD液體培養基(g/L):酵母提取物10,胰蛋白胨20,腺嘌呤3,葡萄糖20。
YPACe培養基(g/L):酵母提取物10,胰蛋白胨20,腺嘌呤3,乙酸鉀20。
KACe培養基(g/L):乙酸鉀20。
1.4 mol/L 山梨醇:255 g山梨醇溶解于1 L 0.5% PBS緩沖液。
1.1.4 儀器與設備
DS-Ri2共聚焦熒光顯微鏡,Nikon公司;TY7622振蕩混合器,易擴中國有限公司;QB-128旋轉培養器,其林貝爾儀器制造有限公司;HYL-A電熱恒溫搖床,上海三發科學儀器有限公司;MX 150高速冷凍離心機,日本日立 HITACHI公司;DYY-6C凝膠電泳儀,北京六一儀器廠;Universal Hood II凝膠成像儀,Bio-Rad(美國)公司;JY92-IIN超聲波細胞粉碎機,寧波新芝生物科技有限公司;NanoDrop2000,賽默飛世爾科技有限公司公司。
1.2 實驗方法
1.2.1 酵母培養與產孢
本文用到的酵母為AN120(野生型,二倍體)。酵母營養細胞使用YPAD培養基,酵母產孢使用YPACe培養基和KACe培養基。
先將釀酒酵母接入5 mL的YPAD培養基于試管中,放入30 ℃,220 r/min的搖床上培養24 h;將5 mL培養物轉入100 mL的YPACe培養基于搖瓶中,放入30 ℃,220 r/min的搖床上培養24 h;離心收集菌體,再將菌體轉入100 mL的KACe培養基于搖瓶中,放入30 ℃,220 r/min的搖床上培養24 h;取5 μL在顯微鏡下觀察產孢率,統計釀酒酵母和孢子的數量,再用孢子數量除以總數即為產孢率,實驗中的產孢率要達到90%;最后離心收集孢子。
1.2.2 孢子的純化
將收集的細胞重懸于5 mL 1.4 mol/L山梨醇中,向其中加入20 μL的1 mg/mL溶菌酶充分混勻,37 ℃孵育2 h后棄上清液。向孢子中加入5 mL水,超聲破碎20 min,將子囊膜破碎,在顯微鏡下觀察,90%以上的孢子處于游離狀態。以3 000×g,2 min離心收集菌體。用0.5% TritonX-100洗滌3~6次,每次均以3 000×g,2 min離心收集菌體,隨后用水洗滌3次,每次均以3 000×g,2 min,最后離心收集孢子。
1.2.3 釀酒酵母孢子表面RNA的提取
將純化好的孢子進行稱重,加入適量水,使孢子終質量濃度為0.1 g/mL,取500 μL,9 000×g離心1 min, 去除水;加入500 μL 0.6 mol/L NaCl溶液,放入振蕩器振蕩3 min;15 000 r/min 離心,取上清液500 μL, 加入RNAiso Plus(TaKaRa)500 μL[18]。充分混勻,冰上放置5 min;加入200 μL氯仿(國藥),迅速混勻,冰上靜置15 min;12 000×g4 ℃ 離心15 min; 取600 μL上清液轉入新的1.5 mL 離心管,加入 600 μL 異丙醇,冰上靜置30 min,隨后12 000×g4 ℃ 離心10 min,棄上清液;加入500 μL 75%乙醇清洗沉淀,7 500×g4 ℃ 離心5 min;去除上清液,冰上靜置10 min, 加入50 μL DEPC水。最后用Nano Drop檢測 RNA的濃度。
1.2.4 釀酒酵母營養細胞與大腸桿菌RNA提取
酵母細胞于5 mL YPAD過夜培養,取1 mL于1.5 mL 離心管中,9 000×g離心1 min,加入適量玻璃珠,加入RNAiso Plus(TaKaRa)1 mL 于振蕩器上振蕩1 min,破碎細胞,放置冰上5 min;加入200 μL氯仿,迅速混勻,冰上靜置15 min;12 000×g4 ℃ 離心15 min;取600 μL上清液轉入新的1.5 mL 離心管,加入 600 μL異丙醇,冰上靜置30 min,12 000×g4 ℃ 離心10 min,倒去上清液;加入500 μL 75%乙醇清洗沉淀,7 500×g4 ℃ 離心5 min;去除上清液,冰上靜置10 min,加入50 μL DEPC水。最后在Nano Drop檢測 RNA的濃度。
大腸桿菌于5 mL LB培養基過夜培養,操作步驟同上。
1.2.5 釀酒酵母孢子表面分子的提取
將純化好的孢子進行稱重,加入適量水,控制孢子終質量濃度為0.1 g/mL,取500 μL,9 000×g離心1 min, 棄上清液;加入500 μL 0.6 mol/L NaCl溶液,放入振蕩器振蕩3 min;15 000 r/min 離心,取上清液500 μL。 考慮到NaCl的濃度對于吞噬效率以及表面分子的影響,對0.6 mol/L NaCl進行10倍稀釋,具體稀釋比例可根據后期需要的RNA的量,計算得出對應的 0.6 mol/L NaCl的量,再進行10倍稀釋。
1.2.6 beads的交聯
不加入戊二醛交聯:將beads置于2 mL離心管中,RNA組(加入定量的RNA),釀酒酵母表面分子組(按照RNA的量,計算出加入釀酒酵母表面分子的量),加入DEPC水,定容至2 mL。最后置于旋轉儀上(4 ℃,24 h),充分交聯[19]。
戊二醛交聯:將beads置于2 mL離心管中,向離心管中加入200 μL 2%戊二醛預處理beads,置于30 ℃ 培養箱1 h;離心(5 000×g, 1 min),并用去離子水洗滌,再離心,重復3次;RNA組(加入定量的RNA),釀酒酵母表面分子組(按照RNA的量,計算出加入釀酒酵母表面分子的量),加入DEPC水,定容至2 mL。最后置于旋轉儀上(4 ℃,24 h),充分交聯。
1.2.7 RNA的檢測
對于釀酒酵母孢子表面分子,分別用RNase(0.5 U) 進行處理,37 ℃,1 h,再進行1.2.6中的交聯。
1.2.8 HEK293T細胞的培養
在長滿HEK293T細胞的培養皿中,去除培養液,加入10 mL新的培養液充分懸浮,并取適量轉入新的24孔培養板中,計數,數量約為2.5×106個,37 ℃培養。
1.2.9 beads的吞噬與顯微鏡觀察計數
將1.2.6中的終產物,3 000×g2 min 離心,去除上清液,加入10 μL 水,混勻,取10 μL加入1.2.8中的HEK293T細胞培養液中,加入30 μL lysosome tracker,37 ℃ 放置1 h。取出培養板,除去培養液,加入100 μL PBS。在顯微鏡下觀察吞噬現象,并計數,計算吞噬的數量,從而得出吞噬效率。
2 結果與分析
2.1 HEK293T細胞對beads的吞噬
首先,在本研究室前期的研究工作中,發現釀酒酵母孢子能夠被HEK293T細胞吞噬,孢子直徑2 μm左右,所以HEK293T細胞對于微米級的顆粒也有著吞噬的作用。為此,研究中購買來自Sigma公司的聚苯乙烯乳膠珠可用于創建乳膠凝集系統,稱為beads,beads表面帶有氨基基團,呈正電荷,直徑2 μm。用beads來進行吞噬研究。研究中將beads和HEK293T細胞共同培養1 h,然后共聚焦顯微鏡下觀察beads內化現象。如圖1所示, beads本身具有藍色熒光,在顯微鏡DAPI下能明顯觀察到;lysosome tracker顯示的紅色即為被吞噬的beads,所以在TRAIC下能觀察到紅色的圓圈即為被吞噬;BF為拍到的beads和HEK293T細胞;MERGE為前3張圖的重疊圖。結果顯示beads能被HEK293T細胞吞噬。考慮到研究中加入的beads的數量是否會改變吞噬效率,因此研究了不同的beads量對于吞噬效率的影響,如圖2所示,100個HEK293T細胞吞噬的beads的數目即為吞噬百分比,進而來表示吞噬的效率,吞噬百分比越高即為100個HEK293T細胞吞噬的beads數目越多。在加入不同量的beads條件下,100個HEK293T細胞吞噬的beads數目在1~2個左右,且差距不大,在后續的研究中使用8×107個beads做吞噬研究。
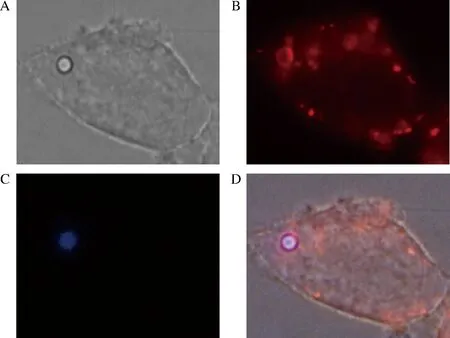
A-BF; B-TRAIC; C-DAPI; D-MERGE

圖2 不同beads量的吞噬效率
2.2 RNA對于吞噬的影響
本研究用0.6 mol/L NaCl溶液洗脫釀酒酵母孢子表面,從NaCl洗脫液中提取RNA再進行純化,然后將RNA和beads一起孵育綁定,RNA帶負電荷,氨基修飾的beads帶正電荷,通過正負電荷作用力進行綁定,形成RNA-beads后再進行吞噬實驗。研究猜測不同的RNA量對于吞噬效率有影響,所以用不同濃度的RNA綁定beads。為了探究不同來源的RNA是否對beads的吞噬有影響,分別從大腸桿菌、釀酒酵母營養細胞中提取了RNA,并與釀酒酵母孢子壁RNA以及購買的tRNA作了比較,不同RNA的凝膠電泳如圖3所示。如圖4-A所示,50 000 ng的RNA效果最優,因此后續的研究都選用50 000 ng的RNA進行反應;吞噬效率的結果如圖4-B所示,結果表明所有RNA對于吞噬都有促進作用,但是來自釀酒酵母孢子表面的RNA和tRNA有更明顯的促進作用,同時由圖3看出,釀酒酵母孢子表面RNA和tRNA 都是長度非常短的RNA,猜測短鏈RNA更能影響beads的吞噬效率。
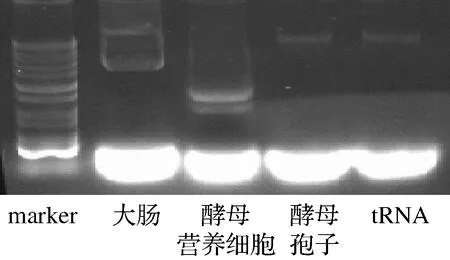
圖3 不同RNA純化后的凝膠電泳圖

A-不同RNA的量;B-不同來源RNA
2.3 RNase對beads和RNA-beads吞噬作用影響
為了進一步確認RNA對于beads的吞噬有著促進作用,因此有必要在體系中分別用RNase處理beads和RNA-beads,再和HEK293T細胞共同孵育,進行吞噬實驗。如圖5所示,結果顯示RNase不會對beads的吞噬產生影響。對于從釀酒孢子表面的RNA進行純化,并對樣品分別加入RNase(0.5 U),37 ℃, 1 h,再進行綁定然后觀察吞噬結果。研究發現RNase可以明顯的降低吞噬效率。因此研究中純化的RNA確實對吞噬有促進作用。
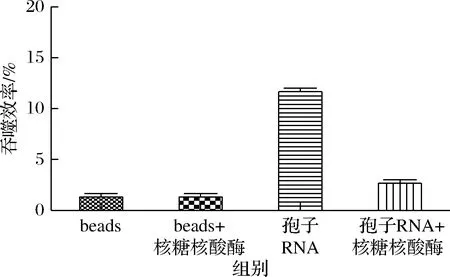
圖5 RNase對只有beads吞噬效率的影響和對加入RNA的beads吞噬效率的影響
2.4 釀酒酵母孢子表面分子和戊二醛交聯作用促進吞噬作用
本研究選擇用0.6 mol/L NaCl進行洗脫釀酒酵母孢子表面的分子,由于RNA也是從NaCl洗脫液中提取純化獲得,研究猜想是否洗脫液也會影響beads的內化,所以用beads直接綁定孢子表面洗脫液,對洗脫液進行了10倍稀釋以防高濃度的NaCl影響beads對分子的綁定。如圖6所示,NaCl的洗脫液相對于對照組、純化后的RNA都有明顯提升。考慮到釀酒酵母表面分子的洗脫液中可能還有其他物質,所以使用交聯劑戊二醛去研究是否其他分子也影響了beads的吞噬,戊二醛能夠作為交聯劑,對于氨基、羧基等有著交聯作用。結果見圖6,戊二醛的加入,使吞噬作用有了顯著提升,這說明孢子洗脫液中除RNA外還有其他分子也可以影響beads的吞噬。研究中對戊二醛的影響做了猜測,戊二醛可能交聯了釀酒孢子表面除RNA外的其他分子,對于該分子需要進一步研究。

圖6 戊二醛的加入對不同分子對于吞噬效率的影響
2.5 釀酒酵母孢子表面分子洗脫液中NaCl濃度對吞噬作用影響
本研究選擇用0.6 mol/L NaCl進行洗脫釀酒酵母孢子表面的分子,考慮到NaCl濃度影響,先前實驗在洗脫后進行了稀釋。在此實驗中,加入了不稀釋組,并在稀釋組與不稀釋組2組的最后體系中測量RNA和蛋白質的量,如表1所示,沒有明顯差異。在吞噬作用實驗中,如圖7所示,2組也不存在差異,因此NaCl濃度對于beads綁定分子吞噬沒有影響。
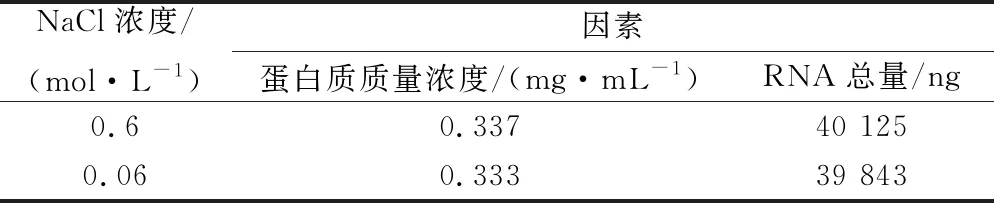
表1 不同濃度NaCl洗脫液中蛋白濃度和RNA總量
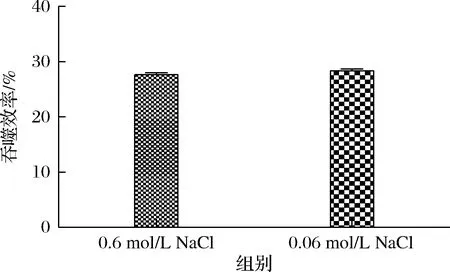
圖7 不同濃度NaCl對于吞噬效率的影響
2.6 RNase對釀酒酵母孢子表面分子促吞噬作用的影響
研究發現高鹽洗脫液也可以誘導beads內化,是否高鹽洗脫液中誘導內化的關鍵分子是RNA,為此用RNase處理了高鹽洗脫液。研究中加入了RNase(0.5 U),在37 ℃放置1 h,再進行綁定交聯然后吞噬觀察結果,如圖8所示,研究發現RNase的處理可以使吞噬數量大幅降低。因此釀酒酵母孢子表面洗脫液中RNA是調節內化的關鍵分子。

圖8 RNase對釀酒酵母表面分子和beads吞噬效率的影響
3 討論
首先,本研究發現釀酒酵母孢子表面的分子可以誘導2 μm顆粒被HEK293T細胞內吞,并對HEK293T細胞內化beads數量做了評估。進一步分析顯示釀酒酵母孢子表面的RNA可顯著影響beads的吞噬,并發現其他物種的RNA也具有誘導beads內化的功能,這說明RNA誘導beads內化具有普遍性。同時研究發現高鹽洗脫液誘導beads內化的效率要比純化后的RNA更為顯著,這說明孢子表面的其他分子也具有調節beads內化的能力,對于該分子有待進一步研究和探討。
釀酒酵母孢子表面結構復雜,由內到外依次是甘露糖層,葡聚糖層,殼聚糖層和二酪氨酸層,最新研究發現孢子表面具有脂滴等結構。0.6 mol/L NaCl 洗脫液中包含了除RNA以外的其他分子促進beads的吞噬,到底是什么分子促進了這一過程需要進一步的探索。在研究中,使用洗脫液孵育戊二醛處理過的beads的吞噬效率最高。對釀酒酵母孢子表面分子和非巨噬細胞的吞噬研究,將為未來藥物運輸的材料和傳遞提供新思路。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
甘肅教育(2020年14期)2020-09-11 07:57:42
中學生數理化(高中版.高考數學)(2020年5期)2020-06-02 09:19:08
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
商周刊(2017年9期)2017-08-22 02:57:49
汽車工程學報(2017年2期)2017-07-05 08:13:02
時代英語·高二(2015年1期)2015-03-16 00:08:11