單核細胞增生李斯特菌進化家系的研究進展
2021-12-02 09:20:42周繼福趙曉美
食品科學 2021年21期
周繼福,王 娉,郭 佳,3,趙曉美,陳 穎,*
(1.南京財經大學食品科學與工程學院,江蘇 南京 210023;2.中國檢驗檢疫科學研究院,北京 100176;3.天津科技大學食品科學與工程學院,天津 300000)
單核細胞增生李斯特菌(Listeria monocytogenes)(簡稱單增李斯特菌)是一種胞內兼性厭氧的可對人類健康造成嚴重危害的食源性致病菌[1]。其分布廣泛,食品(乳及乳制品、生熟肉類、蛋類、即食食品、水果蔬菜等)[2-3],食品加工、儲存設備(設備表面、管道、生產環境和冰箱)[4-5],自然環境(土壤、污水)[6]和動物宿主(哺乳動物、非哺乳動物)細胞[7]中均有分離獲得該菌的報道。單增李斯特菌可在低pH值(3.3~4.2)、低溫、高鹽濃度(0.5%~1.2%)和高壓下存活[8],易感人群為幼童(young)、老年人(old)、孕婦(pregnant)和免疫功能缺陷人群(immune-compromised people),也被簡稱為“YOPI”[8]。感染后,住院率(92%)和死亡率(20%~30%)較高[9]。
根據菌體抗原(O)和鞭毛抗原(H)的血清凝集實驗可以將單增李斯特菌分成13 種血清型(1/2a、1/2b、1/2c、4b、3a、3b、3c、4a、4c、4e、4ab、4d和7)[10-11]。不同血清型菌株的致病性存在差異,從臨床病例和受污染的食品中分離得到的菌株至少95%以上屬于血清型1/2a、1/2b、1/2c、4b[12-13],其中1/2a和1/2b血清型菌株通常為食品分離株[11,14-15]。血清型4b菌株在臨床患者樣本中常見[16],李斯特菌病中約有50%的病例是由4b血清型菌株感染所致。研究認為血清型1/2和4b菌株致病性的差異與菌株細胞的壁磷壁酸(wall teichoic-acid,WTA)組成及其聚合物結構不同有關[17]。依據表型特征和基因分型方法,可以將單增李斯特菌分成4 種進化家系,家系I和家系II菌株的來源種類和數量最多,而家系III較少,家系IV則較為罕見[18]。家系I菌株包含6 種血清型(1/2b、4b、3b、4e、4d和7),家系II菌株包含4 種血清型(1/2a、1/2c、3a和3c),家系III和IV包含的血清型為4b、4a和4c[14],4b血清型菌株在家系I、III和IV中均有發現[19]。菌株進化家系研究通常會結合菌株的血清型(serotypes)、血清組(serogroups)、流行克隆(epidemic clones,ECs)、毒力類型(virulence types,VTs)、克隆復合體(clonal complexes,CCs)和序列類型(sequence types,STs)[20],以比較菌株在系統發育、遺傳多樣性、生態位和致病性方面的異同點。不同家系的菌株致病性和宿主特異性不同,對人類健康的威脅程度也不同,在一些流行病學研究中,需要對單增李斯特菌進化家系進行鑒別,為食品安全的評估、可能的感染暴發提供科技支撐。本文對單增李斯特菌進化家系的劃分依據、不同家系的特點、流行特征、檢測方法等進行了歸納總結,希望能為研究與菌株家系有關的單增李斯特菌病提供一定參考。
1 單增李斯特菌進化家系劃分
單增李斯特菌起初的“家系”相關劃分源于多位點酶電泳(multilocus enzyme electrophoresis,MEE)顯示不同電泳簇(electrophoretic types,ETs)結果和菌株血清型分布不一致[21],為進一步尋找分離株之間的潛在關系而引入“進化家系”的概念。“家系(lineages)”名稱的正式使用是在1997年,研究人員利用核糖體分型和3 種毒力基因hly、actA和inlA將133 株單增李斯特菌劃分為家系I和家系II,且對核糖體片段和16S rRNA基因序列的分析中首次確定菌株另外一種獨特家系——家系III的存在[22]。基于家系III菌株所呈現的獨特性質和非典型性狀,Roberts等[23]采用EcoRI核糖分型和hly基因的聚合酶鏈式反應-限制性酶切長度多態性(polymerase chain reaction-restriction length polymorphism,PCR-RFLP)法對46 株家系III菌株進行分析,系統發育研究證明了家系III存在3 個亞系(III A、III B和III C)[24]。家系IV的正式劃分是在2008年,Ward等[25]利用多位點基因分型(multilocus genotyping,MLGT)方法建立系統發育樹,說明家系IV菌株獨立于家系III。至此,明確了單增李斯特菌可劃分為4 種不同的進化家系[26](表1)。菌株家系的劃分既可以明確分離株之間的相關關系,為菌株的系統發育和致病性研究提供思路,也可以作為菌株溯源和表達菌株生態位關系的依據。

表1 單增李斯特菌的家系劃分Table 1 Classification of the lineages of Listeria monocytogenes
2 單增李斯特菌進化家系的特點
2.1 單增李斯特菌進化家系表型差異
單增李斯特菌分離株遺傳特性的不同導致了菌株表型差異。不同家系菌株在生長速率、毒力相關表型、生物膜形成能力、對NaCl的耐受能力[28]、產細菌素的能力[29]以及對食品的污染等方面均存在差異。這些差異可為食品加工條件的設定、菌株的生態位關系表征及初步判斷菌株毒力強弱提供部分依據和基礎。
不同家系菌株表型差異首先表現在菌株生長速率和對食品的污染方面,在菌株生長速率研究中,不同家系菌株在不同的培養溫度和培養基中的生長速率有所不同,其中37 ℃、含有質量分數6% NaCI的BHI培養基中家系I和III菌株的生長速率明顯高于家系II[20,30]。特定培養環境造成菌株生長速率改變的同時一定程度上影響著菌株對食品的污染和對動物、人類的侵襲,不同處理方法下菌株對食品的污染能力不同,在高壓和溫熱下,家系I菌株對食品的污染能力高于家系II[31]。此外,在多種脅迫條件下家系II菌株的生長和存活能力顯著高于家系I[32-33]。
菌株對細胞的污染和侵襲程度與細菌在細胞表面的生物膜形成相關[34],研究顯示家系II菌株比家系I更易生成生物膜[35],同時家系II菌株生物膜胞外基質中含有更多的胞外DNA[36-37]。胞外基質對細菌的黏附和定植至關重要,發生定植而溶解目標細胞的李斯特菌溶血素(listeriolysin S,LLS)多產生于家系I而非家系II菌株[29]。借助原子力顯微鏡研究細菌黏附發現,家系II菌株與家系I相比,其總體黏附力較高但特異性力(氫鍵)和非特異性力(范德華力和靜電作用力)較低,并且細胞表現出更高的楊氏模量,同時表面形成更長、更硬的生物聚合物[38-39]。
細菌成斑實驗和溶脂表型實驗可為菌株毒力強弱的測定提供初步依據,不同家系菌株形成的噬菌斑、空斑大小和成斑率以及溶脂性現象等毒力相關表型均表現出不同。在噬菌斑成斑實驗中,家系II菌株的成斑率低于家系I和III,同時其形成的噬菌斑更小[22]。類似的,家系I和家系II菌株相較于家系III能形成更大的空斑[40]。相比于家系I和家系II的分離株,家系III菌株在卵黃瓊脂平板上表現出更強的溶脂活性[40]。
2.2 單增李斯特菌進化家系基因組差異
菌株在基因組水平上的差異源于基因特異性研究,不同家系菌株的基因組差異主要體現在基因含量、基因突異、基因重組和基因表達等方面。
菌株的基因組差異首先表現在基因含量上,利用多種基因組分析工具發現家系I和家系II菌株基因含量存在差異,這種差異除前噬菌體數目、編碼蛋白質的偽基因個數和菌株特異性基因外[41],還包括毒力基因、應激反應基因和代謝相關的基因。使用RPKM(reads per kilobase per million mapped)法發現家系II菌株比家系I缺少編碼膜相關蛋白的167 個基因[42]。Chen Jianshun等[43]發現參與編碼菌株精氨酸脫亞氨酶和胍丁胺脫亞氨酶系的部分基因簇在家系I和家系II及大部分家系IV菌株中存在,但在家系III和少部分家系IV菌株則缺乏。對于不同家系菌株的基因組差異,菌株毒素-抗毒素系統的14 個等位基因同樣表現出明顯的差別[44]。與毒力基因相關的差異還包括李斯特菌致病島(Listeriapathogenicity islands,LIPI)LIPI-3和LIPI-4組成基因[45],其中LIPI-3主要存在于家系I分離株[46]。除基因含量的差異外,單核苷酸多態性(single nucleotide polymorphism,SNP)、轉座因子和基因缺失[29,47]等基因突變是基因組水平上最常見的差異。Zhang Xu等[48]通過Python編程語言對菌株基因組數據挖掘后,找到多個可以區分家系I、II、III菌株的特定SNP的候選基因。在基因突變中較為罕見的李斯特菌溶素正調節蛋白(listeriolysin positive regulatory protein,prfA)的移碼及錯義突變使得家系I和家系II菌株的毒力降低[49]。此外,水平基因轉移現象也能反映不同家系菌株在基因水平上的差異,在基因轉移上,家系III菌株對于家系I和家系II是最佳的基因供體[50]。
不同家系菌株的進化形成與同源重組和正向選擇密切相關[51]。菌株系統發育中的基因重組表明,家系II菌株比家系I更容易發生重組[50],這導致了家系II菌株的高遺傳變異性,也使得家系II菌株更容易適應變化的生態位,進而可能影響食品及其相關生態位中家系II和家系I菌株的數量[52]。
2.3 單增李斯特菌進化家系轉錄組差異
轉錄組研究是對基因結構和基因功能的拓展研究,不同家系菌株的轉錄組研究顯示出菌株在噬菌體基因[53]、毒力相關基因、應激調控因子[54]和細胞壁合成[41]等方面的差異。
在證明菌株基因組多樣性的同時,不同家系菌株在毒力基因分布上存在差異,研究顯示高毒力基因和低毒力基因分別分布在家系I和家系II菌株中[7]。在氧化劑、酸和熱響應等應激反應方面的轉錄組研究顯示,家系I比家系II的ST菌株顯示出對氧化劑更高的應激抵抗力,同時家系II菌株在體積分數0.6% H2O2作用下表現更為敏感[33]。Sigma B(σB)作為菌株最主要的調節因子,其對家系I、II和III B菌株在酸和氧化應激條件下的存活起重要作用,但是對家系III A菌株效果不明顯[55]。此外,作為σB的替代調節因子,PrfA也影響著菌株在壓力和毒性下的差異轉錄,菌株對膽汁的應激抵抗研究發現,家系II菌株發生差異轉錄的基因要少于家系I[56]。
此外,Lee等[7]基于多位點序列分型(multilocus sequence typing,MLST)發現家系I和家系II菌株在大部分轉錄水平上存在差異。
2.4 單增李斯特菌進化家系蛋白質組差異
在基因組和轉錄組水平上的研究不足以完全體現不同家系菌株的特點,基因調控和表達的變化同樣會造成不同家系菌株致病性和侵襲能力的差異[57]。不同家系菌株可在蛋白質定量表達[58]、蛋白質表達程度和蛋白質種類(核糖體蛋白種類[59]、表面蛋白)等方面體現差異。
家系I和家系II菌株在蛋白組水平上的直接差異體現在蛋白質表達量上[58]。其次,使用二維凝膠電泳顯示家系II菌株的蛋白表達程度沒有家系I明顯[60]。建立蛋白質種類和家系之間的聯系不僅可以促進家系分型,也可以為菌株致病性研究提供幫助。Ojima-Kato等[59]借助基質輔助激光解吸電離飛行時間質譜(matrix-assisted laser desorption ionization-time of flight mass spectrometry,MALDI-TOF MS)發現不同家系(家系I、II、III)的3 種核糖體蛋白存在差別。對于哺乳動物所需的表面蛋白lmo0320,家系II菌株是家系I的2 倍左右[32]。與菌株毒力相關的表面蛋白在不同家系菌株中同樣存在差異[61]。這些蛋白質水平上的差異研究對于菌株致病性研究和生態位關系的確定有著積極促進作用。
2.5 單增李斯特菌進化家系代謝組差異
通過對菌株細胞內源物質的動態研究發現,不同家系菌株在代謝組的差異主要分為對物質的吸收利用和分解[57]、代謝途徑差異[62]和物質的分泌等方面。
不同成分碳源物質的培養基會影響菌株的生長繁殖。含D-阿洛糖的富集肉湯可以改善菌株的分離并減少非目標細菌的生長,研究發現只有家系II菌株可以利用D-阿洛糖作為生長的碳源,其余家系菌株均不能[63]。此外,家系II菌株還可以代謝蔗糖[57]。研究菌株對物質的分解發現,III B和III C亞系分離株缺乏發酵鼠李糖的能力,但亞系III A則能夠利用鼠李糖和乳酸[23]。這也從側面說明不同家系菌株在碳源利用上存在一定的差異[57]。
由于代謝組位于基因組和轉錄組的下游,因此不同家系菌株在基因組和轉錄組的差異會導致代謝組的不同。轉錄組學分析結果表明家系II和家系I菌株在磷酸鹽代謝、脂質代謝和輔酶輔基代謝中存在不同的富集趨勢[7]。基于全基因組分析菌株遺傳關系時發現家系I和家系II菌株的部分基因座編碼的醛糖1-表異構酶代謝途徑存在差異[62],這表明不同家系菌株基因組和代謝之間存在高度保守性[57]。
2.6 單增李斯特菌進化家系致病性
李斯特菌病的高致死率促進了對單增李斯特菌致病性研究,而菌株的致病性取決于各種毒力因子[64]。目前發現的毒力因子有李斯特致病島家族(LIPI-1、LIPI-2、LIPI-3和LIPI-4)、蛋白質(表面蛋白(Internalin A、Internalin B)和P60蛋白[65])和轉錄調節因子[49,66]等,其中最主要的是LIPI-1。各種毒力因子的致病機理有所差異,如轉錄激活因子PrfA負責調節和控制許多毒力基因的轉錄表達[42],inlAB操縱子編碼的兩個內部蛋白對于進入非吞噬細胞至關重要[66]。不同進化家系菌株的致病性差異還與毒力因子的分泌相關。作為最重要的轉錄調節因子,σB的存在影響著家系I、II和III B菌株對人上皮Caco-2細胞的入侵[55]。不同家系菌株中的毒力基因也影響著菌株的致病性,編碼Internalin蛋白的基因inlG、inlK和inlF通常出現在家系II菌株中[66]。同樣,InlA蛋白(由inlA基因編碼)的存在會影響菌株穿越人腸道屏障[67],而擁有全長inlA基因和LIPI-3/4的菌株通常會導致嚴重的李斯特菌病[68]。Chen Yi等[69]利用全基因組測序(whole-genome sequencing,WGS)技術對即食食品分離株的inlA基因的提前終止密碼子(premature stop codon,PMSC)測序,發現家系I或帶有全長inlA的分離株通常比家系II或帶有inlAPMSCs的菌株具有更大的毒力潛能。除毒力因子外,不同家系菌株的致病性可能和細胞壁的WTA相關[17]。
不同家系菌株致病性強弱相差較大,大多數人類李斯特菌病的暴發是由含4 種外毒素的家系I菌株引起[70],這可能和其高度克隆的特性相關[32]。菌株進化中毒力會發生變化,這與毒力基因突變和相關蛋白丟失有關,研究發現家系II菌株的毒力喪失可能與毒力基因(prfA、inlA和inlB等)的突變有關[71],表面蛋白的缺失同樣會促使家系III菌株毒力發生改變[72]。菌株致病性研究往往依賴于菌株分離,而菌株的分離獲得與國家地域、飲食風俗、衛生條件和采樣方法相關。Chenal-Francisque等[73]對五大洲共300 份分離株進行流行病學研究,發現很少有流行性菌株存在。在一些國家的研究中發現家系II菌株引起的人類李斯特菌病要多于家系I[74]。
3 單增李斯特菌不同進化家系的分型方法
單增李斯特菌流行病學研究中,分離株的家系分型結果和菌株來源有著密切聯系,對分離獲得的菌株進行家系鑒別后,需要進一步研究分析不同來源分離株的關系以說明菌株的遺傳特性,而這種解釋往往依賴于合適的分型方法。將分型方法和生態位相結合可以為李斯特菌病防控和衛生服務建設提供參考。
菌株精確的家系分型能為菌株種內遺傳變異研究提供依據,尤其是菌株致病性方面[85]。最先用于家系分型的方法包括MEE、脈沖場凝膠電泳(pulsed-field gel electrophoresis,PFGE)、基于限制性片段的DNA指紋分型技術和核糖體分型等,其中PFGE使用范圍廣、區分能力強,被認為是分型的“金標準”。Lopez-Valladares等[74]根據PFGE條帶將菌株分為家系I和家系II。然而PFGE結果在不同研究之間的解釋和比較上難以統一且不易準確表明疾病暴發后菌株與環境之間的關聯性[77]。為此,MLST[86-87]、基于SNP的高分辨熔解(high resolution melting,HRM)技術[88]、多位點可變數目串聯重復序列分析(multilocus variable number of tandem repeat analysis,MLVA)分型[89]、多位點毒力基因序列分型(multivirulence-locus sequence typing,MVLST)[90]及其他基于PCR的相關分型方法相繼投入應用。Miya等[77]基于串聯重復區的核苷酸序列,建立比MLST有更強區分能力的多位點串聯重復序列分析(multilocus tandem repeats sequence analysis,MLTSA)分型方法,成功將菌株分為家系I和家系II。馬彥寧等[82]利用LIPI-1的等位基因特異性寡核苷酸,將菌株分為家系I、家系II和家系III。Wang Yi等[81]建立多重PCR方法用于4 種家系和2 個亞系菌株的快速區分鑒定。除此之外,利用不同家系菌株的特定SNP候選基因同樣實現了家系的分型[48]。這些分型方法的建立和應用一定程度上加快了進化家系的遺傳發育和致病性研究。但是MLST和MLVA方法缺乏對于大量疾病暴發株的區分能力[80,91],且MLVA的區分能力略弱于PFGE[80],等位基因特異性寡核苷酸(allele-specific oligonucleotide,ASO)-PCR對單管反應確定菌株家系要求較高。為更好地進行家系菌株分型,WGS越來越多地作為開發基因分型方法的基礎[92],其數據還可以用于毒力基因的檢測和地理歸屬的確定。WGS被廣泛使用在跨區域、多來源的單增李斯特菌株家系分型、遺傳多樣性及致病性研究[93-94]。Chen Yi等[69]通過WGS序列的核基因組MLST(core genome MLST,cgMLST)構建系統發育樹對即食食品分株進行家系分型研究,結合WGS分析確認了家系I、家系II和家系III的相關分離株。比較基因組學方法不僅在致病株遺傳研究和家系分型中應用普遍[95],而且在探究對入侵宿主相關基因[96]和揭示不同家系菌株侵襲力、毒力[57]變化中同樣廣泛使用。除上述家系分型方法外,質譜和菌株表面蛋白也被用于家系分型。Koudelka等[97]利用MALDI-TOF MS對李斯特菌病和食品分離株進行家系分型,基于自建庫的分型準確率為95.7%。Zhang等[98]將菌株的表面蛋白開發為單克隆抗體的抗原靶標,特定單克隆抗體對家系II和I菌株的特異性識別率為91.3%(表2)。
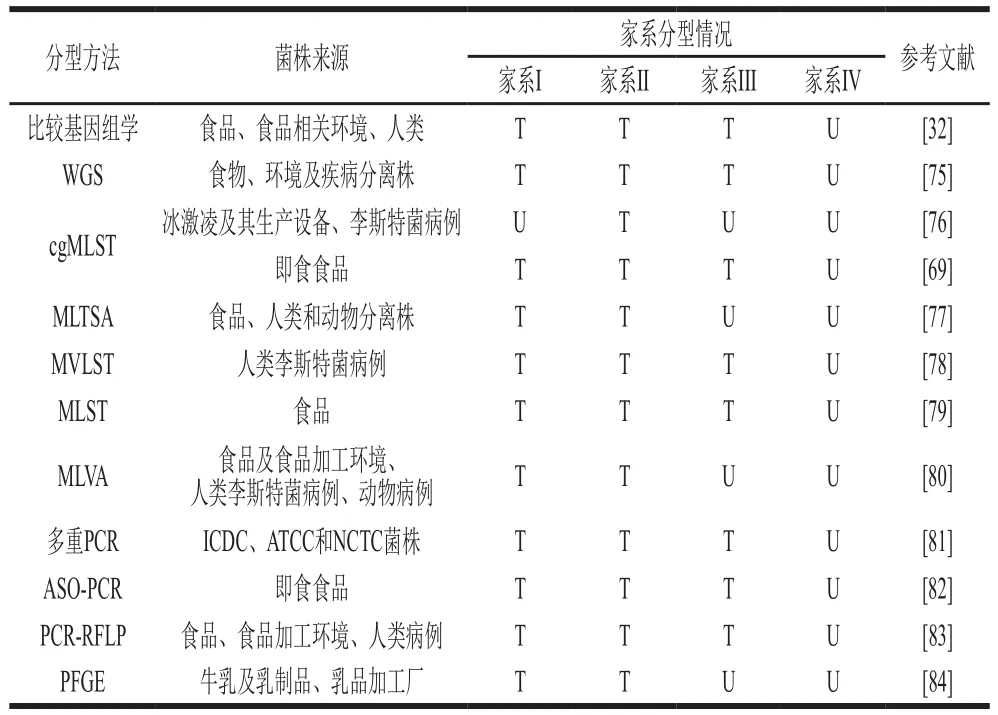
表2 單增李斯特菌典型家系分型方法Table 2 Typing methods for typical lineages of Listeria monocytogenes
基于多種家系分型結果,單增李斯特菌4 種家系之間存在重疊的生態位關系。相較而言,家系I菌株在人類和動物李斯特菌病中分離獲得較多[99],家系II菌株主要分離自食品及食品加工環境[100]。家系III和家系IV菌株則較少分離獲得[101],這可能和菌株的生物膜形成能力有關,加工過程產生的環境壓力造成菌株數量減少,從而難以分離獲得。總之,家系II和家系I菌株較常分離獲得,因而常見于菌株流行病學和遺傳發育研究。為更好地體現家系分型的研究意義,有必要綜合考慮菌株分離來源(國家或地區、分離位點、分離基質和分離季節等)。
4 結 語
綜上所述,單增李斯特菌的進化家系在表型、基因組及致病性等方面均存在差異,在食品安全領域對分離菌株進行家系分析,不僅可以對食品生產加工過程中菌株的耐受能力和膜形成能力做出預先判斷,為食品加工過程中菌株的消滅提供參考;同時,也可以了解不同家系菌株在食品中的分布規律,為食品中單增李斯特菌的風險識別提供基礎數據。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
樂府新聲(2019年2期)2019-11-29 07:34:30
幼兒園(2019年7期)2019-09-05 17:49:18
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
山東女子學院學報(2014年6期)2014-03-01 02:24:55
小小說月刊(2013年6期)2013-05-14 14:55:19
食品科學(2013年15期)2013-03-11 18:25:25
