CysLT1R對APP-HEK293細(xì)胞Aβ生成的調(diào)控作用及其機(jī)制
2021-12-16 06:15:26龍燕,柯璇,洪浩
中國藥理學(xué)通報 2021年12期
龍 燕,柯 璇,洪 浩
(中國藥科大學(xué)藥學(xué)院藥理系,江蘇 南京 211198)
淀粉樣蛋白-β(Aβ)是阿爾茨海默病(Alzheimer′s disease,AD)的主要病理標(biāo)志,Aβ誘導(dǎo)的神經(jīng)毒性和神經(jīng)纖維纏結(jié)被認(rèn)為是引起和延續(xù)AD神經(jīng)退行性過程的關(guān)鍵事件。Aβ級聯(lián)假說認(rèn)為Aβ在腦內(nèi)異常升高、聚集、交聯(lián)、沉積形成老年斑(senile plaque,SP),造成神經(jīng)元變性、壞死、突觸可塑性下降和記憶障礙。Aβ主要有Aβ1-42和Aβ1-40兩種,其中Aβ1-42的疏水性更強(qiáng),更易形成寡聚體,形成SP沉淀的核心。Aβ通過β分泌酶和γ分泌酶先后剪切β淀粉樣蛋白前體蛋白(amyloid β-protein precursor,APP)而產(chǎn)生[1]。正常情況下,Aβ的產(chǎn)生和清除保持平衡,但是當(dāng)APP基因突變、APP裂解酶或Aβ降解酶中任一者功能異常時,Aβ產(chǎn)生和清除間的平衡就被打破,Aβ異常升高導(dǎo)致級聯(lián)病理損傷,形成AD[2]。
半胱氨酰白三烯受體1(cysteinyl leukotriene receptor 1,CysLT1R)是一種G蛋白偶聯(lián)受體,由6個親水環(huán)連接著7個跨膜疏水區(qū)組成[3]。CysLT1R在外周的氣道平滑肌細(xì)胞和上皮細(xì)胞、血管內(nèi)皮細(xì)胞以及嗜酸性粒細(xì)胞等均有表達(dá),在中樞的神經(jīng)元和膠質(zhì)細(xì)胞也有表達(dá)[4]。有研究資料顯示,生理狀態(tài)下大腦CysLT1R表達(dá)水平明顯低于病理狀態(tài)下的水平[5],這提示CysLT1R表達(dá)上調(diào)很可能是組織器官病理反應(yīng)的信號。近年來,人們研究發(fā)現(xiàn)基因敲除CysLT1R和藥物阻斷(普魯司特和孟魯司特)CysLT1R可通過抑制小膠質(zhì)細(xì)胞激活、神經(jīng)炎癥和神經(jīng)毒性,減輕LPS和鏈脲佐菌素誘導(dǎo)的AD樣損傷[6-8];向小鼠腦海馬注射CysLT1R 激動劑半胱氨酰白三烯D4(leukotriene D4,LTD4)可以降低小鼠學(xué)習(xí)記憶能力[9];而CysLT1R拮抗劑普侖司特可以顯著改善Aβ誘導(dǎo)的AD模型小鼠認(rèn)知功能障礙[10]。本課題組近期研究結(jié)果顯示,AD患者海馬和前額葉皮層中CysLT1R蛋白和mRNA水平明顯增加[11]。以上研究均表明CysLT1R在AD的發(fā)病過程中起重要作用,但目前其作用機(jī)制仍不清楚。本研究的目的是探究CysLT1R對APP-HEK293細(xì)胞中Aβ生成的影響并探尋其可能的作用機(jī)制,為進(jìn)一步闡明CysLT1R在AD發(fā)病過程中的重要作用提供理論依據(jù)。
1 材料與方法
1.1 材料
1.1.1細(xì)胞系 穩(wěn)定表達(dá)APP的人胚胎腎293細(xì)胞(APP-HEK293)由中國藥科大學(xué)徐曉軍副教授課題組惠贈。
1.1.2試劑 YM17690(108806-41-3,上海康曼生物科技有限公司),普侖司特(150821-03-7,美國Sigma-Aldrich公司),LV-CysLT1R-EGFP(00015399,上海吉凱基因化學(xué)技術(shù)有限公司),DMEM培養(yǎng)基(12100046,美國Hyclone公司),人Aβ1-40酶聯(lián)免疫試劑盒、人Aβ1-42酶聯(lián)免疫試劑盒(CSB-E06928Eq、CSB-E10684h,武漢華美生物工程有限公司),兔抗APP 抗體、兔抗BACE1抗體、兔抗NF-κB p65抗體(2452S、5606S、4764T,美國CST公司),兔抗PS1抗體、兔抗PS2抗體、鼠抗CysLT1R抗體(sc-7860、sc-1456-R、sc-514181,美國Santa cruz公司),兔抗β-actin抗體、山羊抗兔二抗IgG(BM3873、BA1039,中國博士德生物工程有限公司),Alexa Fluor 488 標(biāo)記驢抗兔 IgG、Alexa Fluor 594標(biāo)記山羊抗小鼠IgG(34206ES60、33212ES60,上海翊圣生物科技有限公司),BCA 蛋白濃度測定試劑盒、DAPI(P0012、C1002,碧云天生物技術(shù)有限公司),膜蛋白、核蛋白和胞質(zhì)蛋白抽提試劑盒(C510002,上海生工生物工程股份有限公司),Clathrin siRNA、β-arrestin-2 siRNA、Rab-5 siRNA(蘇州吉瑪基因股份有限公司),Lipofactamine 2000(11668-019,美國Invitrogen公司),Protein A agarose瓊脂糖珠(16-156,美國Sigma-Aldrich公司)。
1.1.3儀器 RT-6000型酶標(biāo)儀(深圳雷杜生命科學(xué)股份有限公司),轉(zhuǎn)移電泳槽和轉(zhuǎn)移電泳儀(美國Bio-Rad公司),Tanon4200 型化學(xué)發(fā)光成像分析系統(tǒng)、Tanon3500 型數(shù)碼凝膠成像分析系統(tǒng)(上海天能科技有限公司),Eppendorf Master Cycler PCR儀(德國Eppendorf),DMI3000 B熒光顯微鏡(德國 LEICA)。
1.2 方法
1.2.1試劑的配置 含普侖司特的培養(yǎng)基配制:稱取普侖司特0.490 5 g,溶解到1 mL PBS(0.1 mol·L-1,pH 7.4)中,得到終濃度為1.0 mol·L-1的母液。臨用取1.0 μL母液,用培養(yǎng)基稀釋至1 000 mL。含YM17690的培養(yǎng)基配制:稱取YM17690 0.491 5 g,溶解到1 mL PBS(0.1 mol·L-1,pH 7.4)中,得到終濃度為1.0 mol·L-1的母液。臨用取1.0 μL母液,用培養(yǎng)基稀釋至1 000 mL。
1.2.2APP-HEK293細(xì)胞的病毒轉(zhuǎn)染 APP-HEK293細(xì)胞接種于細(xì)胞瓶中。用促轉(zhuǎn)染溶液(enhanced infection solution,Eni.S)將慢病毒稀釋成病毒滴度為1×1010TU·L-1的病毒稀釋液。細(xì)胞瓶中加入病毒稀釋液和Polybrene(50 mg·L-1)稀釋液,轉(zhuǎn)染5 d(120 h)后,檢查病毒轉(zhuǎn)染效率。
1.2.3ELISA法檢測細(xì)胞培養(yǎng)基中Aβ的含量 給予CysLT1R激動劑YM17690(1 μmol·L-1)及拮抗劑普侖司特(1 μmol·L-1)12 h后,收集各組細(xì)胞的培養(yǎng)基上清液,按照試劑盒說明書規(guī)定的方法測定細(xì)胞培養(yǎng)基中Aβ1-40和Aβ1-42的含量。
1.2.4Western blot法檢測APP、BACE1、PS1、PS2蛋白表達(dá)水平和CysLT1R和NF-κB p65的細(xì)胞分布 藥物干預(yù)后用細(xì)胞刮收集細(xì)胞,按照試劑盒說明書步驟分步提取細(xì)胞的膜蛋白、核蛋白和胞質(zhì)蛋白。用BCA法檢測蛋白含量,上樣、電泳、轉(zhuǎn)膜,加入一抗APP(1 ∶1 000)、BACE1(1 ∶1 000)、PS1(1 ∶1 000)、PS2(1 ∶1 000)、β-actin(1 ∶3 000)、CysLT1R(1 ∶1 000)、NF-κB p65(1 ∶1 000)、β-actin(1 ∶3 000)、Histone 3(1 ∶3 000)和Na+-K+-ATPase(1 ∶3 000),4 ℃過夜,洗去一抗,加入適當(dāng)?shù)亩?1 ∶5 000),室溫孵育2 h,洗去二抗,隨后采用化學(xué)發(fā)光成像系統(tǒng)成像,蛋白條帶積分光密度值用Gel-Pro掃描分析。其中以β-actin為胞質(zhì)蛋白內(nèi)參、Histone 3為核蛋白內(nèi)參、Na+-K+-ATPase為膜蛋白內(nèi)參。
1.2.5細(xì)胞免疫熒光法檢測CysLT1R和NF-κB p65的細(xì)胞分布 細(xì)胞接種于24孔板中,培養(yǎng)過夜。給予CysLT1R激動劑YM17690(1 μmol·L-1)及拮抗劑普侖司特(1 μmol·L-1)2 h后,固定封閉處理細(xì)胞,孵育抗體CysLT1R(1 ∶200)、NF-κB p65(1 ∶200),4 ℃過夜,PBS沖洗5 min×3次。加入適當(dāng)?shù)臒晒舛?1 ∶500),室溫避光孵育2 h,PBS沖洗5 min×3次,加入少量DAPI染核5 min,PBS沖洗5 min×3次,加入防淬滅甘油,Leica DMI3000熒光顯微鏡下觀察。
1.2.6siRNA轉(zhuǎn)染 將細(xì)胞接種于6孔板中。取clathrin siRNA、β-arrestin-2 siRNA、Rab-5 siRNA按說明書制成siRNA-Lipofactamine 2000復(fù)合物。6孔板加入siRNA-Lipofactamine 2000復(fù)合物后,細(xì)胞無血清培養(yǎng)基培養(yǎng)6 h后更換為完全培養(yǎng)基,12 h后細(xì)胞分組并開展后續(xù)藥物干預(yù)實驗。
1.2.7免疫共沉淀法分析CysLT1R和NF-κB p65的蛋白相互作用 單獨(dú)給予CysLT1R激動劑YM17690(1 μmol·L-1)或同時給予拮抗劑普侖司特(1 μmol·L-1)2 h后,RIPA裂解液裂解細(xì)胞,離心后收集上清液,加入Protein A瓊脂糖珠,離心,收集上清,棄去Protein A瓊脂糖珠。BCA法測定蛋白含量,稀釋蛋白至1 g·L-1,以每1 mL總蛋白中加入1 μg抗體的比例(1 ∶1 000)加入NF-κB p65抗體,4 ℃過夜,等量Normal Rabbit IgG作為陰性對照。加入Protein A瓊脂糖珠,4 ℃孵育過夜。洗滌、離心,重復(fù)6次。上樣緩沖液重懸瓊脂糖珠-抗原抗體復(fù)合物,99 ℃變性10 min,電泳,轉(zhuǎn)膜,加入一抗CysLT1R(1 ∶1 000)4 ℃過夜。洗去一抗,加入適當(dāng)?shù)亩?1 ∶5 000),室溫孵育2 h,洗去二抗,隨后采用化學(xué)發(fā)光成像系統(tǒng)成像。
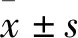
2 結(jié)果
2.1 激活CysLT1R增加APP-HEK293細(xì)胞中Aβ的生成轉(zhuǎn)染LV-CysLT1R-EGFP 5 d后,Western Blot驗證病毒轉(zhuǎn)染效率,結(jié)果顯示:與陰性對照組相比,感染LV-CysLT1R-EGFP的APP-HEK293細(xì)胞中CysLT1R表達(dá)水平明顯增加(Fig 1)。ELISA檢測結(jié)果顯示,CysLT1R激動劑YM17690升高APP-HEK293細(xì)胞中Aβ1-40和Aβ1-42的分泌水平,而預(yù)先給予CysLT1R拮抗劑普侖司特能抑制Aβ1-40和Aβ1-42的分泌(Fig 2A、B)。

Fig 1 LV-CysLT1R-EGFP transfection significantly Western blot detection of the expression of CysLT1R in APP-HEK293 cells after LV-CysLT1R-EGFP transfection.*P<0.01 vs LV-EGFP group.
為了進(jìn)一步探究CysLT1R影響Aβ生成的具體環(huán)節(jié),給予CysLT1R激動劑YM17690(1 μmol·L-1)及拮抗劑普侖司特(1 μmol·L-1)12 h后,我們采用Western blot檢測APP-HEK293細(xì)胞中APP、β分泌酶BACE1、γ分泌酶亞基PS1、PS2的表達(dá),結(jié)果顯示,激活CysLT1R對APP和PS1、PS2的表達(dá)沒有影響,但能上調(diào)BACE1的表達(dá),并且預(yù)先給予普侖司特能抑制BACE1的上調(diào)(APP:P>0.05;BACE1:P<0.05;PS1:P>0.05;PS2:P>0.05;n=4,Fig 3A-F)。

Fig 2 Activation of CysLT1R increased Aβ 1:LV-EGFP+Veh+Veh;2:LV-CysLT1R-EGFP+Veh+Veh;3:LV-CysLT1R-EGFP+YM17690+Veh;4:LV-CysLT1R-EGFP+YM17690+Pran.A:Levels of Aβ1-40 in cell culture medium;B:Levels of Aβ1-42 in cell culture medium.*P<0.05 vs LV-CysLT1R-EGFP+Veh+Veh group.
2.2 激活CysLT1R引起受體和NF-κB p65亞基的核轉(zhuǎn)位GPCR在激活后往往發(fā)生內(nèi)化從而下調(diào)細(xì)胞表面受體的水平,因此,我們觀察CysLT1R激活后細(xì)胞內(nèi)受體的分布情況。Western blot結(jié)果顯示,激活CysLT1R使細(xì)胞膜及細(xì)胞質(zhì)中受體水平明顯下降,而細(xì)胞核中受體水平升高,并且預(yù)先給予普侖司特能抑制受體分布的這種變化(Fig 4A-C)。免疫熒光實驗也證實了這一結(jié)果(Fig 5)。已有研究表明,CysLT1R的激活與NF-κB信號通路有關(guān)[10],為此我們研究了NF-κB p65亞基的細(xì)胞分布情況。Western blot結(jié)果顯示,激活CysLT1R使細(xì)胞質(zhì)中NF-κB p65亞基水平下降,而細(xì)胞核中其水平升高,并且預(yù)先給予普侖司特能抑制NF-κB p65亞基的分布變化(Fig 6A、B)。
2.3 抑制CysLT1R核轉(zhuǎn)位減少細(xì)胞中Aβ的生成已有文獻(xiàn)報道,CysLT1R的內(nèi)化依賴于clathrin、β-arrestin-2和Rab-5等蛋白的輔助[12],為了探究CysLT1R對Aβ生成的影響是否與其內(nèi)化后的核轉(zhuǎn)位相關(guān),我們首先應(yīng)用siRNA分別敲減上述三個內(nèi)化輔助因子。Western blot結(jié)果顯示,敲減clathrin和β-arrestin-2顯著抑制了APP-HEK293細(xì)胞中CysLT1R激活后發(fā)生的受體核轉(zhuǎn)位,而敲減Rab-5對此無明顯影響(Fig 7A-F)。免疫熒光實驗也證實了這一結(jié)果(Fig 7G、H)。檢測Aβ結(jié)果顯示,敲減clathrin和β-arrestin-2顯著抑制Aβ1-40和Aβ1-42的分泌,而敲減Rab-5對此無明顯影響(Fig 7F-H)。此外,敲減clathrin和β-arrestin-2還顯著抑制BACE1的表達(dá),而對APP的表達(dá)沒有明顯影響(Fig 8A-C)。

Fig 3 Effect of CysLT1R activity on expression of APP,BACE1,PS1,1:LV-EGFP+Veh+Veh;2:LV-CysLT1R-EGFP+Veh+Veh;3:LV-CysLT1R-EGFP+YM17690+Veh;4:LV-CysLT1R-EGFP+YM17690+Pran.A:Expression of APP and BACE1 in APP-HEK293 cells after administration of YM17690 (1 μmol·L-1)and pranlukast (1 μmol·L-1)for 12 h;B:Statistical analysis of the expression of APP;C:Statistical analysis of the expression of BACE1;D:Expression of PS1 and PS2 in APP-HEK293 cells after administration of YM17690 (1 μmol·L-1)and pranlukast (1 μmol·L-1)for 12 h;E:Statistical analysis of the expression of PS1;F:Statistical analysis of the expression of PS2.*P<0.05 vs LV-CysLT1R-EGFP+Veh+Veh group.

Fig 4 Nuclear translocation of CysLT1R by Western 1:LV-EGFP+Veh+Veh;2:LV-CysLT1R-EGFP+Veh+Veh;3:LV-CysLT1R-EGFP+YM17690+Veh;4:LV-CysLT1R-EGFP+YM17690+Pran.A:The expression of CysLT1R in nucleus.B:The expression of CysLT1R in cytoplasm.C:The expression of CysLT1R on the cell membrane.*P<0.05 vs LV-CysLT1R-EGFP+Veh+Veh group.
2.4 抑制CysLT1R核轉(zhuǎn)位能抑制NF-κB p65亞基的核轉(zhuǎn)位CysLT1R核轉(zhuǎn)位與NF-κB信號通路的相關(guān)性研究結(jié)果顯示,敲減clathrin和β-arrestin-2能抑制NF-κB p65亞基的核轉(zhuǎn)位(Fig 9A、B)。
2.5 CysLT1R與NF-κB p65亞基在細(xì)胞核內(nèi)存在相互作用為了研究CysLT1R入核的意義,我們采用免疫共沉淀法研究了CysLT1R與NF-κB p65亞基是否存在相互作用。結(jié)果顯示,單獨(dú)給予YM17690激活CysLT1R時,核蛋白提取物經(jīng)anti-NF-κB p65抗體沉淀后仍能檢測到CysLT1R蛋白,表明CysLT1R與NF-κB p65亞基間存在相互作用,而預(yù)先給予普侖司特能抑制兩者之間的這種相互作用(Fig 10A)。而在其胞質(zhì)蛋白提取物中暫未發(fā)現(xiàn)CysLT1R與NF-κB p65亞基的相互作用(Fig 10B)。

Fig 5 Nuclear translocation of CysLT1R by cell immunofluorescence 1:LV-EGFP+Veh+Veh;2:LV-CysLT1R-EGFP+Veh+Veh;3:LV-CysLT1R-EGFP+YM17690+Veh;4:LV-CysLT1R-EGFP+YM17690+Pran.A:The location of CysLT1R in the nucleus;B:Statistical results of the nuclear localization of CysLT1R.*P<0.05 vs LV-CysLT1R-EGFP+Veh+Veh group.

Fig 6 Nuclear translocation of NF-κB p65 1:LV-EGFP+Veh+Veh;2:LV-CysLT1R-EGFP+Veh+Veh;3:LV-CysLT1R-EGFP+YM17690+Veh;4:LV-CysLT1R-EGFP+YM17690+Pran.A:The distribution of NF-κB p65 subunit in nucleus;B:The distribution of NF-κB p65 subunit in cytoplasm.*P<0.05 vs LV-CysLT1R-EGFP+Veh+Veh group.

Fig 7 Inhibition of nuclear translocation of CysLT1R reduces Aβ production in 1:Veh+Scramble siRNA;2:YM17690+Scramble siRNA;3:YM17690+Clathrin siRNA;4:YM17690+β-arrestin-2 siRNA;5:YM17690+Rab-5 siRNA.A-C:Expression of CysLT1R in nucleus,cytoplasm and cell membrane by Western blot and the statistical results;D-E:Expression of CysLT1R in nucleus,cytoplasm and cell membrane by cell immunofluorescence staining and the statistical results;F-H:Levels of Aβ1-40 and Aβ1-42 by Western blot and the statistical results.*P<0.05 vs YM17690+Scramble siRNA group.

Fig 8 Inhibition of nuclear translocation of CysLT1R reduced BACE1 1:Veh+Scramble siRNA;2:YM17690+Scramble siRNA;3:YM17690+Clathrin siRNA;4:YM17690+β-arrestin-2 siRNA;5:YM17690+Rab-5 siRNA.A-C:Expression of BACE1 and APP by Western blot and the statistical results.*P<0.05 vs YM17690+Scramble siRNA group.

Fig 9 Knockdown of clathrin and β-arrestin-2 inhibited nuclear translocation of NF-κB p65 1:Veh+Scramble siRNA;2:YM17690+Scramble siRNA;3:YM17690+Clathrin siRNA;4:YM17690+β-arrestin-2 siRNA;5:YM17690+Rab-5 siRNA.A:The distribution of NF-κB p65 subunit in nucleus by Western blot and the statistical results.B:The distribution of NF-κB p65 subunit in cytoplasm by Western blot and the statistical results.*P<0.05 vs YM17690+Scramble siRNA group.

Fig 10 CysLT1R interacted with NF-κB p65 subunit in nucleusA:The nucleoprotein extracts were prepared and co-immunoprecipitated by NF-κB antibody with IgG as negative control.B:The cytoplasmic extracts were prepared and co-immunoprecipitated by NF-κB antibody with IgG as negative control.
3 討論
有研究表明,CysLT1R參與并調(diào)節(jié)AD的發(fā)病過程[8-11],并且CysLT1R在腦內(nèi)表達(dá)隨年齡的增長而增加,這與Aβ沉積增加和行為缺陷密切相關(guān)[13]。但目前對CysLT1R調(diào)節(jié)Aβ積累的具體作用機(jī)制的研究仍較匱乏,同時CysLT1R與NF-κB途徑之間的關(guān)系也需進(jìn)一步闡明。YM17690 是一種選擇性CysLT1R激動劑,其遠(yuǎn)遠(yuǎn)優(yōu)于LTD4的化學(xué)穩(wěn)定性使它逐漸成為研究CysLT1R特性的一個重要工具藥物[9-10]。因此,本實驗采用 YM17690 作為CysLT1R特異性激動劑。
CysLT1R是一種G蛋白偶聯(lián)受體,由6個親水環(huán)連接著7個跨膜疏水區(qū)組成[4]。長久以來,GPCR都被視為是一類細(xì)胞表面受體,近年來,已發(fā)現(xiàn)越來越多的GPCR在細(xì)胞內(nèi)其他的一些區(qū)室發(fā)揮作用,如細(xì)胞核、線粒體、內(nèi)質(zhì)網(wǎng)等。目前已發(fā)現(xiàn)超過40種細(xì)胞核GPCR,細(xì)胞核GPCR大部分來源于質(zhì)膜轉(zhuǎn)位[14]。已經(jīng)有大量的研究表明,GPCR內(nèi)化的最可能機(jī)制是通過clathrin包被的小泡進(jìn)行內(nèi)吞[15]。2005年首次有研究報道在細(xì)胞核中觀察到了CysLT1R[16]。此后的研究表明,如同多數(shù)GPCR一樣,CysLT1R在質(zhì)膜被激活后隨即經(jīng)內(nèi)吞作用發(fā)生內(nèi)化。CysLT1R的內(nèi)化由clathrin、β-arrestin-2和Rab-5共同介導(dǎo)[12]。基于此,我們使用siRNA在APP-HEK293細(xì)胞中分別敲減上述三種蛋白。結(jié)果顯示,敲減clathrin或β-arrestin-2可以有效抑制CysLT1R的核轉(zhuǎn)位,而敲減Rab-5對此無影響。Rab-5介導(dǎo)的內(nèi)體融合通常標(biāo)志著受體進(jìn)入降解程序[12],這提示我們本實驗中高表達(dá)CysLT1R受體的APP-HEK293細(xì)胞受體內(nèi)化后的去路是核轉(zhuǎn)位,并且CysLT1R的核轉(zhuǎn)位是其參與Aβ生成調(diào)控的必要環(huán)節(jié)。
有研究表明,Aβ激活NF-κB有2種可能機(jī)制:一種是通過RAGE間接激活,一種是直接激活方式[17]。并且,Aβ1-42誘導(dǎo)NF-κB報告基因的轉(zhuǎn)錄,誘導(dǎo)NF-κB p65亞基向細(xì)胞核移位[18]。本實驗中,我們還發(fā)現(xiàn)激活CysLT1R可以引起NF-κB p65亞基的核轉(zhuǎn)位,CysLT1R與NF-κB p65亞基間在細(xì)胞核內(nèi)相互結(jié)合。因此,我們有理由推測,激活CysLT1R引起受體內(nèi)化與核轉(zhuǎn)位,從而上調(diào)BACE1表達(dá)進(jìn)而誘導(dǎo)細(xì)胞生成Aβ1-42,后者誘導(dǎo)NF-κB p65亞基向細(xì)胞核轉(zhuǎn)位。而NF-κB p65亞基向細(xì)胞核移位是激活NF-κB信號通路的重要步驟,從而進(jìn)一步促進(jìn)Aβ生成,最終導(dǎo)致AD的淀粉樣變性和神經(jīng)退行性變。當(dāng)然,CysLT1R與NF-κB p65亞基間在細(xì)胞核內(nèi)相互結(jié)合說明這兩種蛋白之間存在相互作用,這種相互作用對NF-κB信號通路產(chǎn)生何種影響仍需進(jìn)一步探究。
綜上所述,CysLT1R的核轉(zhuǎn)位及核分布參與了NF-κB信號通路的激活,這與該受體誘導(dǎo)Aβ的生成有關(guān)。
