吉祥草甾體皂苷RCE-4誘導人宮頸癌Ca Ski細胞凋亡與自噬相互作用的分子機制
2021-12-16 06:15:30游方芳陳劍鋒
中國藥理學通報 2021年12期
游方芳,程 凡,鄒 坤,張 晶,陳劍鋒
(三峽大學生物與制藥學院天然產物研究與利用湖北省重點實驗室,湖北 宜昌 443002)
宮頸癌是一種常見的婦科惡性腫瘤,化療是主要的治療手段之一,但耐藥性和嚴重的毒副作用往往導致治療預期不佳。天然來源的藥物因其多靶點、毒性低的優勢一直備受關注。RCE-4(結構見Fig 1)是本實驗室從百合科鈴蘭族植物吉祥草中分離得到的一個螺甾烷型甾體皂苷,實驗室前期研究發現,RCE-4對宮頸癌Ca Ski細胞具有較好的選擇性細胞毒性,能誘導其發生凋亡和自噬[1-3],對荷瘤裸小鼠的最高抑瘤率可達到69.1%,且對正常組織的毒性極低[4],顯示了其治療宮頸癌的巨大潛力。但同時也發現了RCE-4的一些不足之處,比如用藥劑量較大、抑瘤率還有待提高以及其抗宮頸癌作用機制研究還不夠深入等。
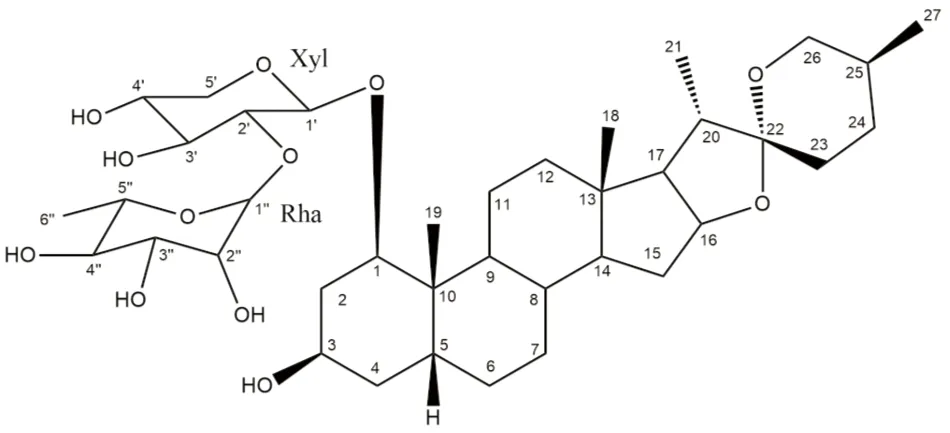
Fig 1 The chemical structure of RCE-4
化療藥物的最終目標是誘導腫瘤細胞死亡,凋亡和自噬作為細胞程序性死亡的主要方式,盡管其特征和機制不同,但大量研究已證實,二者并不是孤立存在的,一些關鍵的調控因子在雙方的調控網絡中頻繁地出現,說明在兩者之間存在著復雜的相互作用。現有的研究表明兩者之間可能存在著如下幾種可能:(1)促進關系。當凋亡和自噬其中一種被激活或者抑制時,另外一種也相應被激活或抑制。宋峰等[5]發現,自噬激動劑雷帕霉素預處理細胞上調自噬可進一步增加IL-1β誘導的細胞凋亡,而使用自噬抑制劑3-MA預處理抑制細胞自噬后則可逆轉IL-1β誘導的INS-1胰島細胞凋亡。(2)拮抗關系。兩者之間是對抗的關系,自噬對腫瘤細胞起保護作用,對腫瘤的發展起促進作用。Zhuang等[6]發現,人參皂甙Rh2誘導的自噬可以保護人白血病U937和K562細胞免于凋亡,3-MA處理能增強其誘導的細胞凋亡。(3)合作關系。自噬和凋亡協作引起細胞死亡,兩者可能同時發生;或者當其中一種受到阻礙或抑制時,另一程序則被激活并繼續完成細胞死亡進程[7]。雖然凋亡和自噬相互作用的分子機制尚未完全闡明,但探討化療藥物誘導的細胞凋亡和自噬之間的關系并進行精準調控對腫瘤治療和抗腫瘤藥物研發具有非常重要的意義,已成為當前引人注目的研究領域之一。
鑒于此,本實驗擬在前期研究的基礎上,通過不同抑制劑和siRNA質粒的應用,初步探討RCE-4誘導的Ca Ski細胞凋亡和自噬的相互作用,以期找到降低用藥劑量、提高其療效的途徑,為RCE-4將來的臨床應用提供理論支撐。
(1β,3β,5β,25S)-spirostan-1,3-diol1-[α-L-rhamnopyranosyl-(1→2)-β-D-xylopyranoside]
1 材料與方法
1.1 材料
1.1.1細胞株 人宮頸癌 Ca Ski細胞株由中國科學院典型培養物保藏委員會細胞庫提供。
1.1.2藥物及試劑 RCE-4由本實驗室從吉祥草中分離得到[8];自噬抑制劑3-甲基腺嘌呤(3-Methyladenine,3-MA)(M9281)、巴弗洛霉素A1(bafilomycin A1,Baf A1)(HY-100558)和氯喹(chloroquine,CQ)(C6628)及凋亡抑制劑Z-VAD(OH)-FMK(Z-VAD-FMK)(HY-16658)均購于中國Med Chem Express公司;基因沉默質粒Bcl-2 siRNA(6441S)、Beclin 1 siRNA(6222S)一抗Beclin 1(3495S)、phospho (p)-Beclin 1(84966S)、LC3 Ⅰ/Ⅱ(4108S)、ATG 4B(13507S)、ATG 14(96752S)、Bax(5023T)、Bcl-2(4223S)、cleaved caspase-3(9664T)、cleaved caspase-7(8438T)、cleaved caspase-9(9505T)及β-actin(8457S)購于美國CST公司;二抗辣根酶標記山羊抗小鼠IgG(GB23301,武漢賽維爾生物科技有限公司);RPMI 1640細胞培養基(31800-022,美國Gibco公司);四季青胎牛血清(11011-8611,浙江天杭生物科技股份有限公司);青霉素-鏈霉素混合液(SV30010,北京索萊寶科技有限公司);四甲基偶氮唑鹽(MTT)(BS0328,美國Amersco公司);ECL超敏發光液(BL520A,北京普利萊基因技術有限公司);GFP-CERTIED?細胞凋亡/壞死檢測試劑盒(ENZ-51002-100,美國ENZO公司);AO/EB雙熒光染色試劑盒(DA0039,雷根生物公司)。
1.1.3儀器 滅菌器(型號:HVE-50)購于日本Hirayama公司;CO2培養箱(型號:9040-0112)購于德國Binder公司;熒光顯微鏡(型號:XD30A-RFL)購于寧波舜宇儀器有限公司;高速離心機(型號:FC5515R)購于德國Eppendorf公司;酶標儀(型號:INFINITE 200 PRO)購于瑞士Tecan公司;流式細胞儀(型號:FACS Verse)購于美國 BD 公司;低速離心機(型號:SC-2542)購于安徽中科中佳科學儀器有限公司;超純水儀(型號:Smart-S15UVF)購于上海和泰儀器有限公司Smart-S15UVF;電泳儀(型號:EPA300)購于北京基昂生物科技有限公司。
1.2 方法
1.2.1細胞培養及傳代 Ca Ski 細胞用RPMI 1640培養基(含10%胎牛血清、1% 青霉素-鏈霉素混合液)置于37 ℃、5% CO2飽和濕度培養箱中孵育,待Ca Ski細胞貼壁生長,取對數生長期細胞用于實驗,0.25%胰酶消化傳代。
1.2.2MTT法檢測細胞增殖 以0.5、1、2、4、6、8、16、32 μmol·L-1RCE-4單用及分別與作用濃度為8 mmol·L-1的3-MA、0.4 μmol·L-1的Baf A1、30 μmol·L-1的CQ或30 μmol·L-1的Z-VAD-FMK聯合使用,并設置對照組,每組設定5個復孔,檢測對Ca Ski細胞的增殖抑制效果。收集對數生長期Ca Ski細胞,調整細胞懸液濃度并以1×107個·L-1的細胞濃度種96孔板,每孔加入100 μL,四周用PBS液封,于37 ℃、5% CO2飽和濕度培養箱孵育,待細胞貼壁后:分別設置空白對照組;3-MA組;Baf A1組;CQ組;Z-VAD-FMK組;RCE-4組;RCE-4+3-MA組;RCE-4+Baf A1 組;RCE-4+CQ組;RCE-4+ Z-VAD-FMK組;RCE-4+Beclin 1 siRNA組;RCE-4+Bcl-2 siRNA組。反應48 h后每孔加入5 g·L-1MTT工作液20 μL,避光孵育4 h后,每孔加入150 μL DMSO,室溫震蕩3 min。在酶標儀上492 nm波長處測定各孔的吸光度值,計算對細胞的抑制率。細胞抑制率(%)=1-(加藥孔吸光度值-空白孔吸光度值)/(對照孔吸光度值-空白孔吸光度值)×100%,每組重復3次,取平均值。
1.2.3AO/EB染色觀察凋亡細胞形態學變化 取對數生長期的Ca Ski細胞以2×108個·L-1的單細胞懸液1 mL接種于24孔板中,待細胞貼壁后,分別設置空白對照組;RCE-4組;Z-VAD-FMK組;RCE-4+Z-VAD-FMK組;RCE-4+Beclin 1 siRNA組。于37°C、5% CO2飽和濕度培養箱孵育12 h后,收集細胞,用PBS沖洗2次,AO Stain Buffer 1×清洗細胞1次,加入適量的AO Stain Buffer 1×重懸細胞,按照細胞懸液 ∶AO Stain=19 ∶1的比例混合,并加入適量AO、EB染色液,輕輕混勻,室溫避光染色15 min,在熒光顯微鏡(激發濾光片波長488 nm,阻斷濾光片波長515 nm)下觀察凋亡細胞形態學變化,計數并拍照。
1.2.4流式細胞儀檢測細胞凋亡率 細胞瓶傳代培養細胞,細胞貼壁生長至80%左右時,分別設置空白對照組;3-MA組;Baf A1組;CQ組;Z-VAD-FMK組;RCE-4組;RCE-4+3-MA組;RCE-4+Baf A1 組;RCE-4+CQ組;RCE-4+ Z-VAD-FMK組;RCE-4+Beclin 1 siRNA組。反應24 h后將培養基吸入離心管中,向培養瓶中加3-5 mL PBS清洗兩次,加3-5 mL不含EDTA的胰酶消化收集細胞。收集細胞,離心,去上清,PBS清洗兩次(3 000 r·min-1,3 min),晾干,按照Buffer ∶水=1 ∶9配制細胞混懸液,按照懸液 ∶染色液=100 ∶1的比例加入適量凋亡、壞死染色試劑,輕輕混勻,室溫避光染色15 min,流式細胞儀進行檢測。
1.2.5Western blot檢測自噬及凋亡相關蛋白表達 取對數生長期的Ca Ski細胞用RCE-4單用或與3-MA、Baf A1、CQ、Z-VAD-FMK、Beclin 1 siRNA、Bcl-2 siRNA聯合應用處理后,于37 ℃、5% CO2飽和濕度培養箱中孵育24 h后,收集細胞,提取Ca Ski細胞蛋白,蛋白定量后,加樣品緩沖液煮沸5 min,各取20 μL加樣。樣品經SDS-PAGE分離,轉移至PVDF膜上。5%脫脂奶封閉2 h后,用TBST洗滌3次×10 min,一抗(1 ∶1 000)4 ℃孵育過夜,洗滌3次×10 min,加二抗(1 ∶5 000)室溫孵育1 h。再用ECL試劑可視化凝膠條帶,并使用發光成像系統(Tanon 5200)記錄結果,使用ImageJ軟件計算灰度值。
2 結果
2.1 RCE-4誘導的細胞自噬對凋亡的影響采用不同的自噬抑制劑(3-MA、CQ和Baf A1)和RCE-4共處理Ca Ski細胞,通過對細胞增殖和凋亡的評估,探索RCE-4誘導的自噬的作用以及和凋亡的關系。
2.1.1不同自噬抑制劑對RCE-4所誘導的自噬流的影響 細胞自噬包括吞噬泡的形成、自噬小體的形成、自噬小體與溶酶體融合形成自噬溶酶體、自噬溶酶體的降解4個步驟[9],自噬流是這些步驟在細胞內連續出現的動態過程,自噬流中的任一環節出現障礙自噬將無法順利進行。3-MA可通過抑制class III PI3K而抑制自噬小體的形成,CQ可通過改變溶酶體的pH從而阻止自噬小體與溶酶體的融合,Baf A1可阻止自噬溶酶體的降解[10]。
2.1.2RCE-4與自噬抑制劑聯合應用對Ca Ski細胞增殖的影響 結果如Fig 2所示,當用不同的自噬抑制劑阻斷自噬流時,Ca Ski細胞對RCE-4的敏感性明顯增強,其中,Baf A1與RCE-4的協同效果最佳,使RCE-4對Ca Ski細胞的IC50由4.669 μmol·L-1減少到1.368 μmol·L-1(P<0.01)。
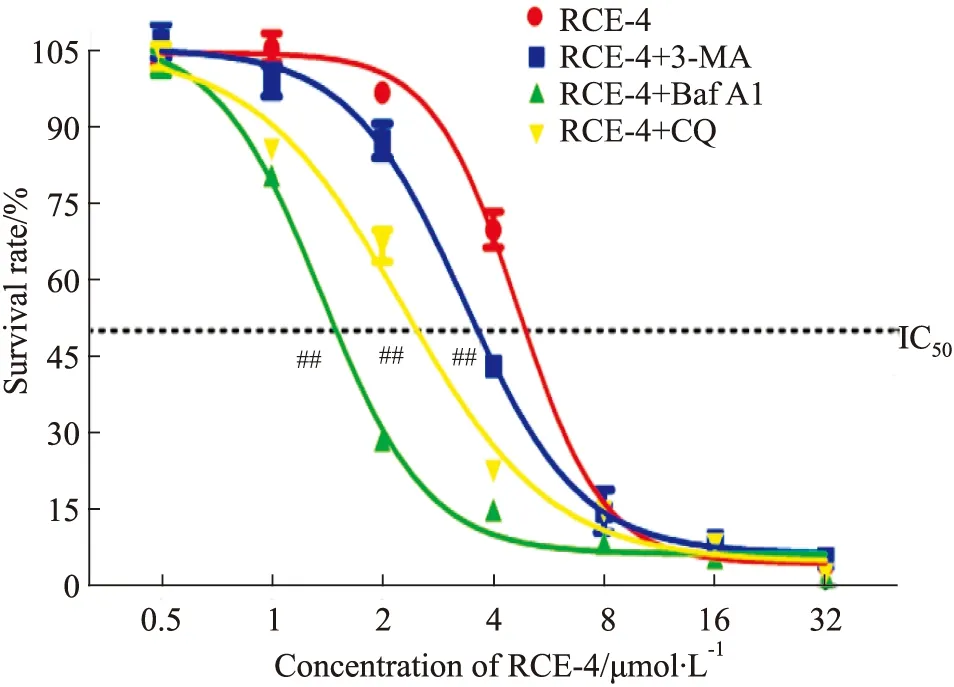
Fig 2 Proliferation of Ca Ski cells inhibited by RCE-4 combined with or without autophagy inhibitors n=3)##P<0.01 vs RCE-4
2.1.3RCE-4與自噬抑制劑聯合應用對Ca Ski細胞凋亡的影響 結果如Fig 3所示,與單用RCE-4組相比,當用不同的自噬抑制劑與RCE-4聯用時,細胞凋亡率明顯升高,其中,Baf A1與RCE-4的協同效果最佳,使RCE-4所誘導的Ca Ski細胞凋亡率由49.96% 增加到91.08%(P<0.01)。同時,如Fig 4A,B和C所示,與單用RCE-4組相比,聯用自噬抑制劑使凋亡相關蛋白cleaved caspase-3、cleaved caspase-9的表達進一步增加,Bcl-2的表達進一步降低,表明當RCE-4誘導的自噬被抑制后,其誘導的凋亡增強(P<0.01)。
2.2 RCE-4誘導的細胞凋亡對自噬的影響與上一個實驗的設計相似,我們擬采用凋亡抑制劑和RCE-4共處理Ca Ski細胞,通過對細胞增殖和自噬的評估,探索RCE-4誘導的凋亡的作用以及和自噬的關系。以前的研究已證實RCE-4能誘導Ca Ski細胞發生線粒體途徑的凋亡,因此,本實驗選用Z-VAD-FMK來抑制凋亡。Z-VAD-FMK是一種細胞滲透性,不可逆的泛caspase抑制劑,能夠抑制caspase家族蛋白酶活性從而同時阻斷線粒體途徑和死亡受體途徑的凋亡。
2.2.1RCE-4與凋亡抑制劑聯合應用對Ca Ski細胞增殖的影響 結果如Fig 5所示,當RCE-4與Z-VAD-FMK聯用時,與單用RCE-4組相比,細胞增殖差異無顯著性,凋亡抑制劑Z-VAD-FMK與RCE-4無協同效果。
2.2.2RCE-4與凋亡抑制劑聯合應用對Ca Ski細胞自噬的影響 結果如Fig 6所示,當RCE-4與Z-VAD-FMK聯用時,與單用RCE-4組相比,LC3II、Beclin 1、p-Beclin 1、ATG 4B、ATG 14的表達量降低,表明當RCE-4誘導的凋亡被部分抑制時,其誘導的自噬也相應被削弱(P<0.01)。
2.3 Beclin 1在RCE-4誘導的細胞凋亡自噬互反饋調控中的作用Beclin 1是自噬進程的關鍵調控因子之一,它可以與HMGB-1、ATG 14、Vps34、Vps15等自噬相關蛋白結合,形成不同的復合體,在不同階段參與自噬的進程[11]。考慮到Beclin 1的重要性,我們用siRNA 質粒將其沉默,以觀察Beclin 1對RCE-4所誘導的凋亡和自噬的影響。
2.3.1RCE-4與Beclin 1 siRNA聯合應用對Ca Ski細胞增殖的影響 結果如Fig 7所示,當應用Beclin 1 siRNA處理后,Ca Ski細胞對RCE-4的敏感性明顯增強(P<0.01),使RCE-4對Ca Ski細胞的IC50由4.669 μmol·L-1減少到1.538 μmol·L-1。
2.3.2RCE-4與Beclin 1 siRNA聯合應用對Ca Ski細胞凋亡的影響 結果如Fig 9所示,不管是單用Beclin 1 siRNA還是與RCE-4共處理Ca Ski 細胞,Beclin 1的表達均被明顯抑制,表明Beclin 1 siRNA發揮了作用。同時,與單用RCE-4組相比,Beclin 1siRNA與RCE-4共處理組中cleaved caspase-3/7/9和Bax的表達均明顯增加(P<0.01),而抗凋亡蛋白Bcl-2的表達則明顯降低(P<0.01),表明抑制Beclin 1促進了RCE-4誘導的Ca Ski細胞凋亡,Beclin 1在RCE-4誘導的凋亡和自噬互反饋調控中扮演了重要角色。
2.4 RCE-4與Bcl-2 siRNA 的聯合應用Bcl-2因具有拮抗細胞凋亡的功能而被廣泛熟知,但現有研究發現Bcl-2同時也是凋亡自噬互反饋調控的關鍵聯系蛋白之一[12]。因此,我們也重點考察了Bcl-2在RCE-4所誘導的凋亡自噬互反饋調控中的作用。
2.4.1RCE-4與Bcl-2 siRNA聯合應用對Ca Ski細胞增殖的影響 結果如Fig 10所示,當應用Bcl-2 siRNA處理后,Ca Ski細胞對RCE-4的敏感性明顯增強(P<0.01),使RCE-4對Ca Ski細胞的IC50由4.669 μmol·L-1減少到1.582 μmol·L-1,表明Bcl-2在RCE-4誘導的凋亡和自噬互反饋調控中扮演了重要角色。
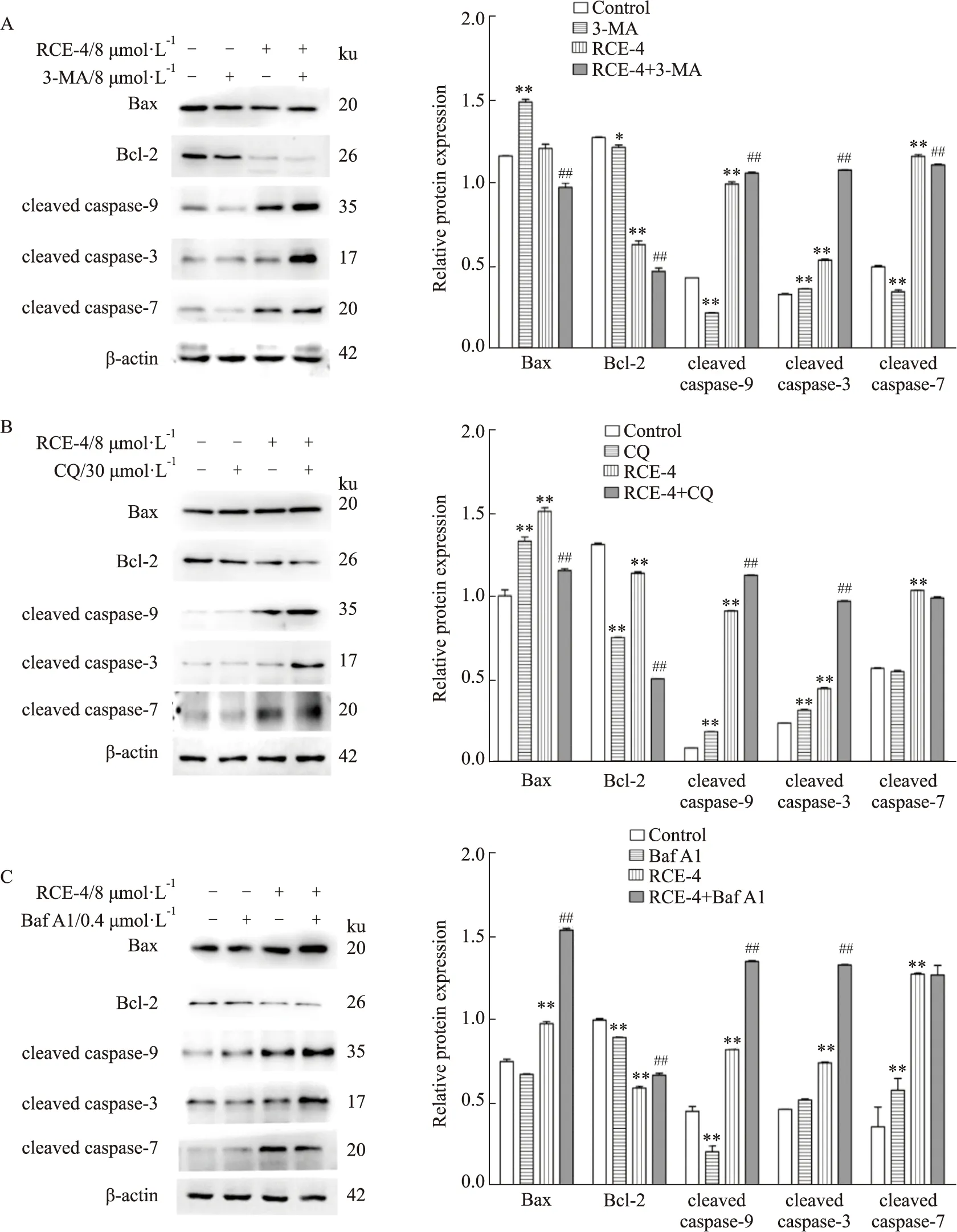
Fig 4 Expression of apoptosis related proteins in Ca Ski cells treated with RCE-4 combined with or without autophagy inhibitors n=3)*P<0.05 vs control,**P<0.01 vs control,##P<0.01 vs RCE-4
2.4.2RCE-4與Bcl-2 siRNA聯合應用對Ca Ski細胞自噬的影響 結果如Fig 11所示,不管是單用Bcl-2 siRNA還是與RCE-4共處理Ca Ski 細胞,Bcl-2的表達均被明顯抑制,表明Bcl-2 siRNA發揮了作用。同時,與單用RCE-4組相比,Bcl-2 siRNA與RCE-4共處理組中Beclin 1、p-Beclin 1、LC3II、ATG 4B和ATG 14的表達均明顯增加(P<0.01),表明抑制Bcl-2促進了RCE-4誘導的Ca Ski細胞自噬。
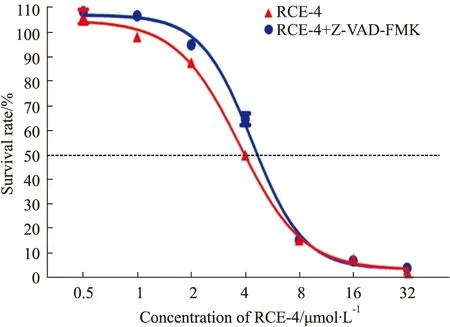
Fig 5 Proliferation of Ca Ski cells inhibited by RCE-4 combined with or without apoptosis inhibitors n=3)
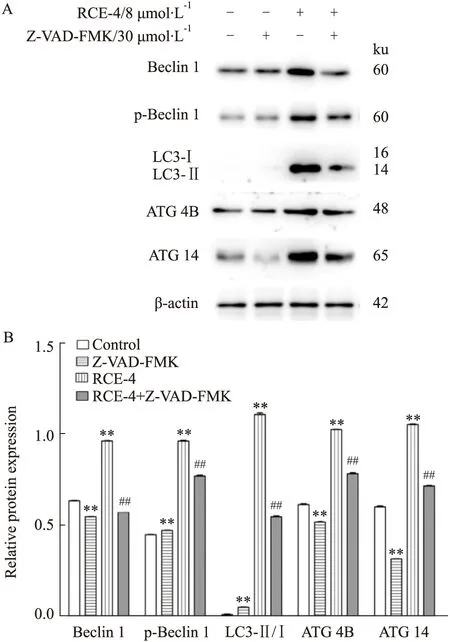
Fig 6 Expression of autophagy related proteins in Ca Ski cells treated with RCE-4 combined with or without apoptosis inhibitors n=3)**P<0.01 vs control,##P<0.01 vs RCE-4.
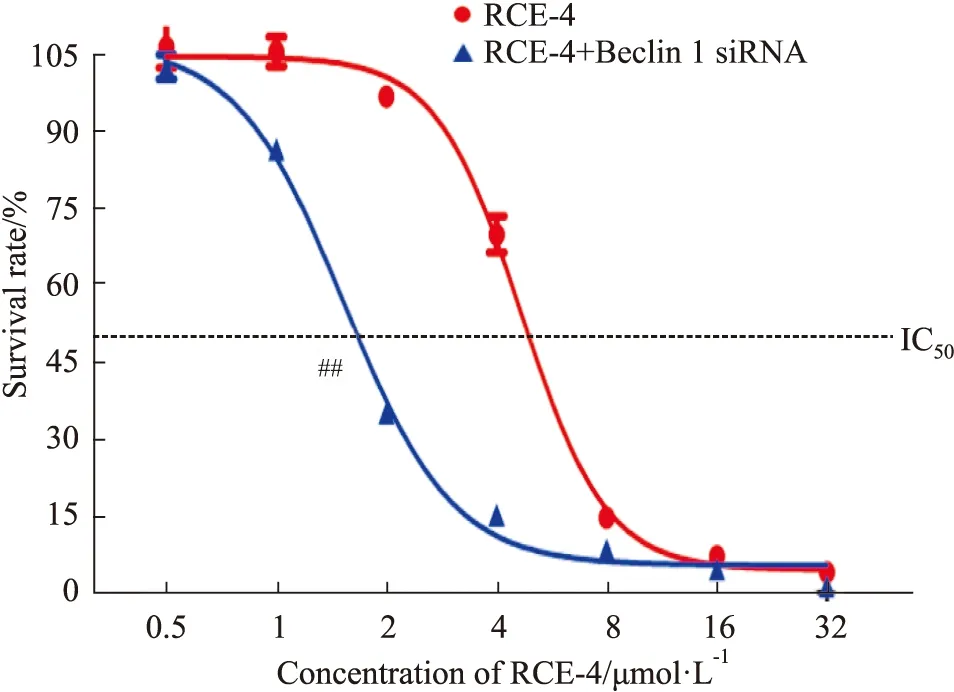
Fig 7 Proliferation of Ca Ski cells inhibited by RCE-4 combined with or without Beclin 1 siRNA n=3)##P<0.01 vs RCE-4.
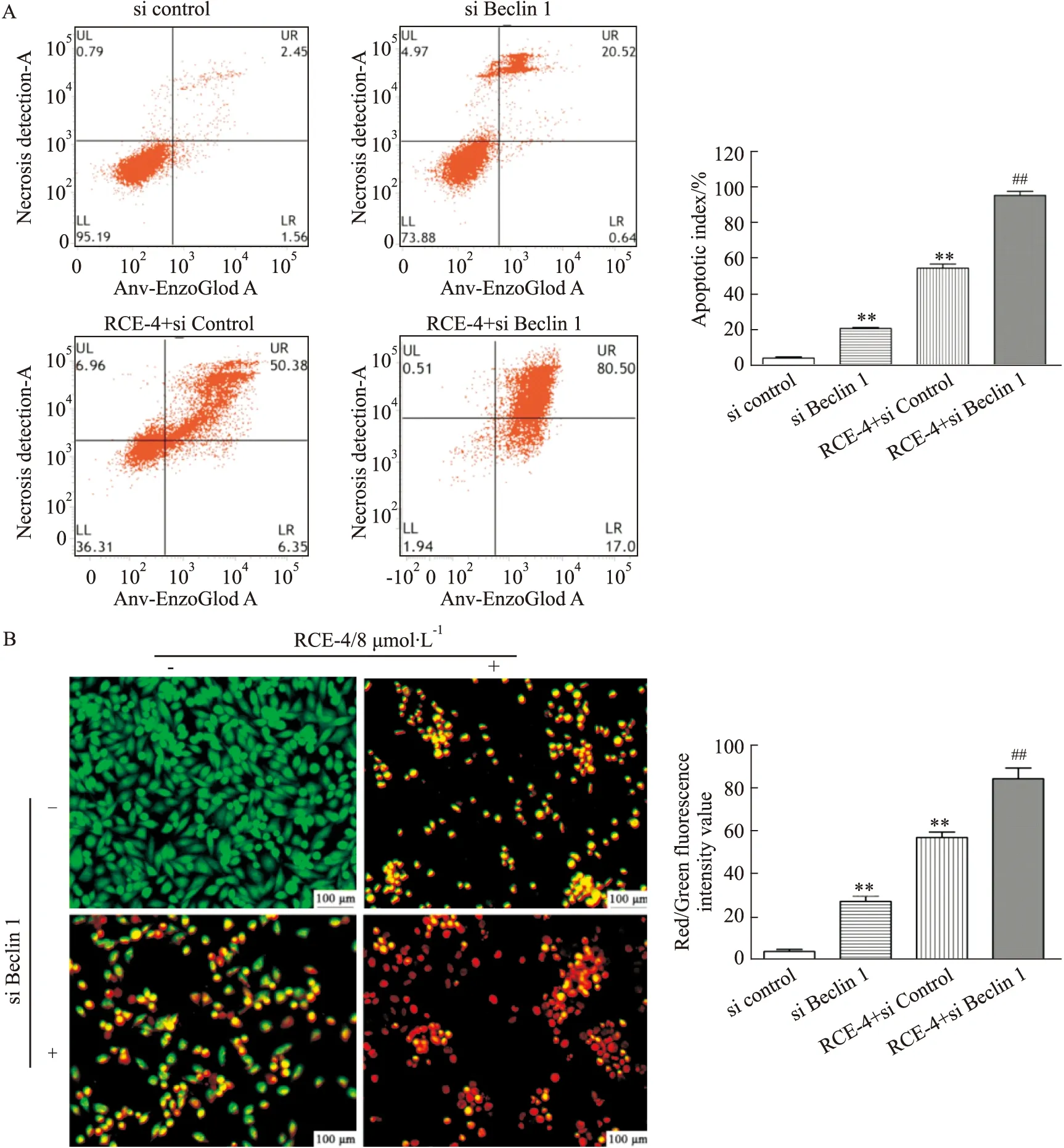
Fig 8 Effect of RCE-4 combined with Beclin 1 siRNA on apoptosis of Ca Ski cells n=3)Scale:400×.**P<0.01 vs control,##P<0.01 vs RCE-4.
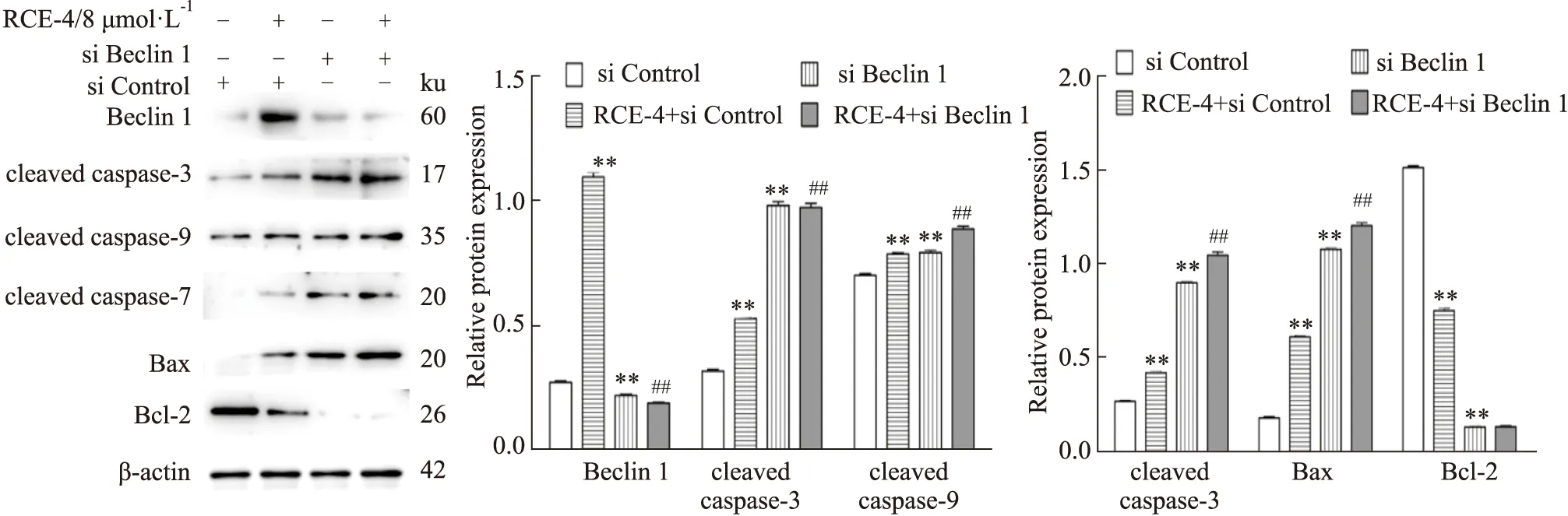
Fig 9 Expression of apoptosis-related proteins in Ca Ski cells treated with RCE-4 combined with or without Beclin 1 siRNA n=3)**P<0.01 vs control,##P<0.01 vs RCE-4.
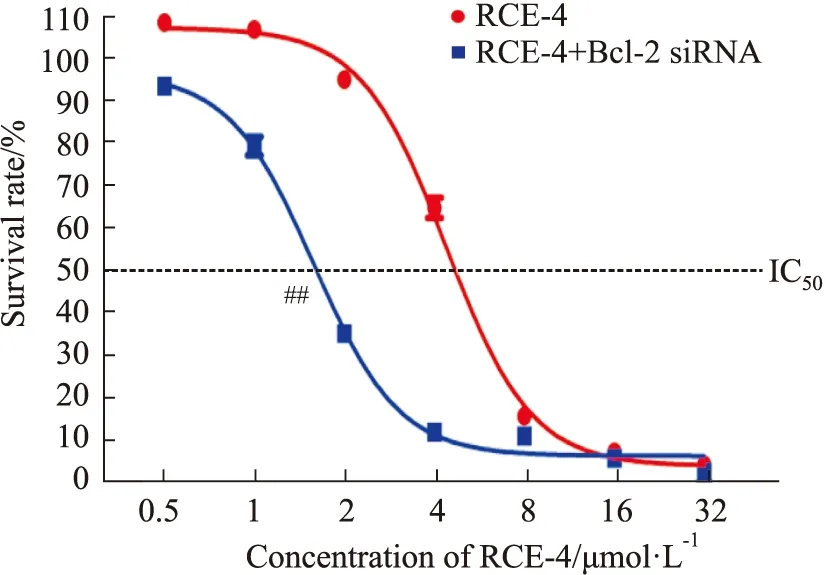
Fig 10 Proliferation of Ca Ski cells inhibited by RCE-4 combined with or without Bcl-2 siRNA n=3)##P<0.01 vs RCE-4.
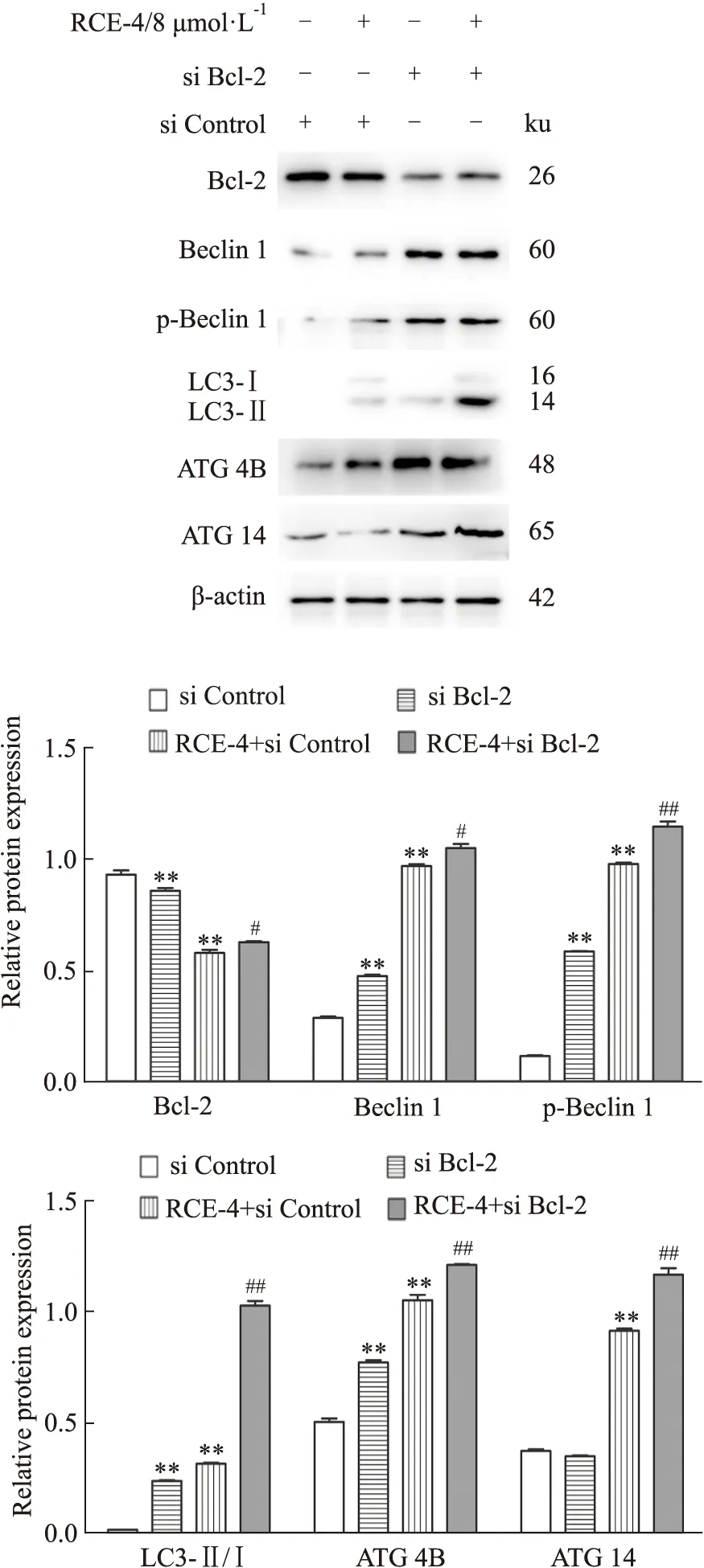
Fig 11 Expression of apoptosis-related proteins in Ca Ski cells treated with RCE-4 combined with or without Bcl-2 siRNA n=3)**P<0.01 vs control,#P<0.05 vs RCE-4,##P<0.01 vs RCE-4.
3 討論
本研究中,我們首次解析了凋亡和自噬在RCE-4所誘導的Ca Ski細胞程序性死亡中的作用。首先,應用3種作用于自噬流不同階段的抑制劑與RCE-4共處理Ca Ski細胞后,Ca Ski細胞對RCE-4的敏感性和凋亡率均得到不同程度的增強,表明RCE-4所誘導的自噬可能是保護性的,對于Ca Ski細胞是一種存活機制。一般情況下,當腫瘤細胞受到外界刺激(如化療)后,誘發的自噬可以通過加速受損細胞器或蛋白聚集物的清除,使大分子物質得以循環利用,保護腫瘤細胞免于死亡,這也是腫瘤細胞對抗化療,產生耐藥性的機制之一[13-14];在3種自噬抑制劑中,Baf A1對RCE-4的協同效果最好,這可能是由于Baf A1阻斷了自噬流的最后一道關卡—自噬溶酶體的降解,對自噬流的阻斷最為徹底,凋亡得到了最大程度的激活所致。需要指出的是,雖然誘導的自噬是保護性的,但這并不表明促進自噬就能降低細胞死亡,自噬在腫瘤細胞中常常扮演“雙刃劍”的角色,過度激活的自噬很可能會導致細胞的自噬性死亡以及凋亡增加,細胞的死亡反而會進一步增加[15-16]。
其次,我們應用凋亡抑制劑Z-VAD-FMK與RCE-4共處理Ca Ski細胞,結果發現,當凋亡被部分抑制時,自噬水平也相應降低,表明RCE-4誘導的凋亡對觸發自噬起著重要作用。另外,相對于單用RCE-4組,凋亡抑制劑Z-VAD-FMK與RCE-4聯用時細胞死亡情況并沒有發生統計學變化,推測其原因可能是Z-VAD-FMK只是部分的抑制了凋亡。另外,聯用時其誘導的自噬也相應的被削弱,其保護性作用降低,故綜合來看,細胞的程序性死亡進程沒有受到太大的影響。
自噬起始因子Beclin l和抗凋亡蛋白Bcl-2在凋亡和自噬的信號轉導通路以及相互作用中扮演了關鍵角色[11,17]。比如Beclin l可被Bcl-2家族中的促凋亡蛋白Bax裂解成“C端片段”和“N端片段”而失去誘導自噬的能力,裂解的Beclin 1碎片可進入線粒體內,促進細胞色素C的釋放,進而啟動細胞凋亡[17]。Bcl-2可以通過BH3結構域與Beclin1結合形成Bcl-2-Beclin 1復合體,該復合體被認為是細胞程序性死亡進程的“核心”和“總開關”[18]。鑒于此,我們應用siRNA來分別沉默其表達,探索其在RCE-4所誘導的凋亡和自噬互作中的作用。結果顯示,當Beclin 1的表達被抑制之后,自噬流被阻斷,RCE-4誘導的凋亡率則顯著增強,表明Beclin 1在RCE-4誘導的凋亡中起著負調控作用,下調Beclin 1有利于凋亡的發生。而應用Bcl-2 siRNA之后則明顯不同,抑制Bcl-2既明顯增強了自噬,也增強了RCE-4誘導的凋亡以及細胞死亡。推測其原因,可能是此時自噬處于過度活化的狀態,導致細胞發生自噬性死亡,從而導致RCE-4對Ca Ski細胞的抑制作用更為明顯。因此,Bcl-2在RCE-4誘導的自噬中也起著負調控作用,下調Bcl-2有助于凋亡和自噬的深度激活,從而增強Ca Ski細胞對RCE-4的敏感性。
綜上所述,本研究首次闡明了凋亡和自噬在RCE-4所誘導的Ca Ski細胞程序性死亡中的作用。在RCE-4誘導的Ca Ski細胞程序性死亡中,凋亡占據著主導地位,對觸發自噬起著重要作用,而自噬則起著保護細胞、拮抗凋亡的作用。自噬抑制劑Baf A1和RCE-4有最佳的協同作用,Beclin 1和Bcl-2在RCE-4誘導的Ca Ski細胞程序性死亡中起著關鍵的負調控作用,可通過對其的抑制來增強Ca Ski細胞對RCE-4的敏感性,以降低RCE-4用藥劑量,提高其療效。
(致謝:本研究在三峽大學生物與制藥學院天然產物研究與利用湖北省重點實驗室完成,衷心感謝本研究的指導老師和各位參與者!)
