支鏈脂肪酸的食源性分析及活性機(jī)理研究進(jìn)展
2022-01-06 02:31:58黃夢瑤王存芳
食品科學(xué) 2021年23期
關(guān)鍵詞:研究
黃夢瑤,程 明,王存芳,*,姜 華,*
(1.齊魯工業(yè)大學(xué)(山東省科學(xué)院)食品科學(xué)與工程學(xué)院,山東 濟(jì)南 250353;2.青島市畜牧獸醫(yī)研究所,山東 青島 266000)
支鏈脂肪酸(branched-chain fatty acids,BCFA)是在直鏈碳鏈上具有一個或多個支鏈的飽和脂肪酸,其支鏈多為甲基,根據(jù)支鏈甲基的數(shù)量可分為單支鏈脂肪酸和多支鏈脂肪酸。當(dāng)支鏈甲基在碳骨架的倒數(shù)第二個碳原子上時稱為異構(gòu)支鏈脂肪酸(iso-BCFA),在倒數(shù)第三個碳原子上時稱為反異構(gòu)支鏈脂肪酸(anteiso-BCFA)[1]。特殊的支鏈結(jié)構(gòu)使得BCFA具有低熔點(diǎn)、防凍、氧化穩(wěn)定性和良好的熱穩(wěn)定性。目前國內(nèi)外對BCFA的研究還相對較少,且多集中在反芻動物制品。本文對BCFA的食物來源及活性機(jī)理進(jìn)行了總結(jié)歸納,以期為BCFA的進(jìn)一步研究提供理論依據(jù)。
1 BCFA的食源性分析
1.1 BCFA的母乳源性分析
母乳是嬰兒的最佳營養(yǎng)來源,1958年首次從母乳中檢測出BCFA[2],BCFA在母乳中的質(zhì)量分?jǐn)?shù)高達(dá)1.5%。以嬰兒對乳汁的需求量計(jì)算,3 月齡嬰兒對BCFA的攝入量約為23 mg/kg,遠(yuǎn)高于其對二十二碳六烯酸(docosahexenoic acid,DHA)的攝入量(12 mg/kg)[3]。母乳中主要的BCFA組成及其質(zhì)量分?jǐn)?shù)如表1所示。有研究表明,隨著泌乳期的延長,母乳中BCFA的含量均有所下降,其中anteiso-C17:0、anteiso-C15:0和iso-C17:0下降最為明顯[4]。BCFA在母乳中的含量與多種因素有關(guān),其中飲食是最大的影響因素。Dingess等[5]對美國辛辛那提州、上海和墨西哥3個地區(qū)母親(產(chǎn)后4 周)所分泌的乳汁進(jìn)行脂肪酸組成分析。結(jié)果顯示不同地區(qū)母乳中BCFA的含量存在顯著性差異,辛辛那提州母乳中BCFA的含量約為上海地區(qū)的2 倍。分析差異原因與飲食有關(guān),歐美地區(qū)的人們多食用乳制品和牛羊肉,而在中國人群飲食中,乳制品和反芻動物肉制品的占比比較低。這說明母乳中的BCFA含量受哺乳期母親飲食的影響很大,尤其是反芻類動物制品的攝入量。
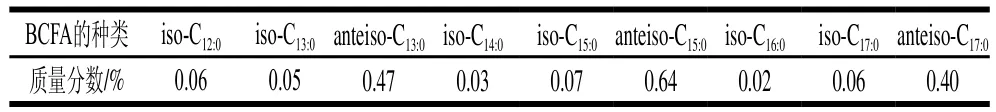
表1 母乳中BCFA組成及其含量[4]Table 1 Composition and contents of BCFAs in breast milk[4]
不同脂肪酸在母乳乳脂甘油三酯中的位置不同,導(dǎo)致嬰幼兒對脂肪酸的消化吸收速度有所不同。油酸、棕櫚酸和亞油酸是母乳乳脂中3 種主要的脂肪酸,油酸主要分布在Sn-1位,棕櫚酸主要分布在Sn-2位,而亞油酸在Sn-3位上[6]。Sn-1和Sn-3位上不飽和脂肪酸優(yōu)先釋放被腸道吸收,Sn-2位的棕櫚酸可阻止不溶性皂鈣的形成,從而提高嬰幼兒對鈣、礦物質(zhì)的吸收[7]。有研究表明[8],與甘油三酯的Sn-1和Sn-3位相比,BCFA在母乳乳脂中的Sn-2位上富集(約占BCFA的68%),但是在牛羊乳脂中BCFA在Sn-1、Sn-2和Sn-3位呈隨機(jī)分布。在人體腸道中脂肪酸會被分解成2-單酰甘油(2-monoacylglycerols,2-MAGs)和游離脂肪酸的形式再被吸收,Sn-2位的BCFA會以2-MAGs的形式優(yōu)先被人類腸道細(xì)胞攝取,且Sn-2位的BCFA在腸細(xì)胞中的攝取量約為游離脂肪酸的2 倍;母乳中BCFA在Sn-2位的富集特性有利于嬰兒腸道對BCFA的消化吸收,尤其對于嬰幼兒來說,母乳是其出生之后獲取BCFA的最直接途徑。因此,母乳喂養(yǎng)的嬰兒對BCFA的攝取能力更佳,可為嬰幼兒奶粉中BCFA的強(qiáng)化提供參考。
1.2 BCFA的畜產(chǎn)品源性分析
乳脂是BCFA的主要來源,不同反芻動物乳脂BCFA的碳原子數(shù)大多為14~18,但其含量有所不同。王蕾等[9]對乳脂中主要BCFA含量的分析結(jié)果表明,anteiso-C17:0在乳脂中的含量最高,可達(dá)0.501 g/100 g。馬露[10]分別對奶牛、牦牛、水牛、娟珊牛、山羊、駱駝以及馬等不同物種的乳汁中BCFA含量進(jìn)行了研究,由圖1可見,牦牛乳中BCFA含量最高,而馬乳中BCFA含量最低;對于BCFA的類型而言,在所有動物乳中anteiso-C13:0的含量最低,在4 種牛乳中iso-C15:0的含量最高,山羊乳和駱駝乳中anteiso-C17:0的含量最高,馬乳的iso-C15:0含量最高。孫萬成等[11]對不同泌乳期的牦牛乳進(jìn)行了BCFA的檢測,結(jié)果表明,全乳期牛乳中BCFA的含量為2.00 g/100 g,牦牛乳中BCFA的含量為:全乳期4.07 g/100 g、半乳期為7.49 g/100 g。其中,牦牛乳中含量最高的BCFA是anteiso-C15:0和anteiso-C17:0[12]。有研究進(jìn)一步對牦牛酥油及其酥油磷脂中的BCFA進(jìn)行檢測分析[13],結(jié)果顯示牦牛酥油中的BCFA含量顯著高于酥油磷脂中的BCFA含量,而且冬季與夏季得到的牦牛酥油及其酥油磷脂中BCFA含量均不同,其中冬季牦牛酥油中BCFA質(zhì)量分?jǐn)?shù)可高達(dá)9.10%,夏季酥油磷脂中BCFA質(zhì)量分?jǐn)?shù)僅有3.24%。Ran-Ressler等[14]對美國零售牛奶中BCFA含量進(jìn)行了測定,發(fā)現(xiàn)樣品中anteiso-BCFA占BCFA總量的一半以上,且BCFA平均占乳脂脂肪酸的(2.05±0.14)%,與Pegolo等[15]的研究結(jié)果一致。
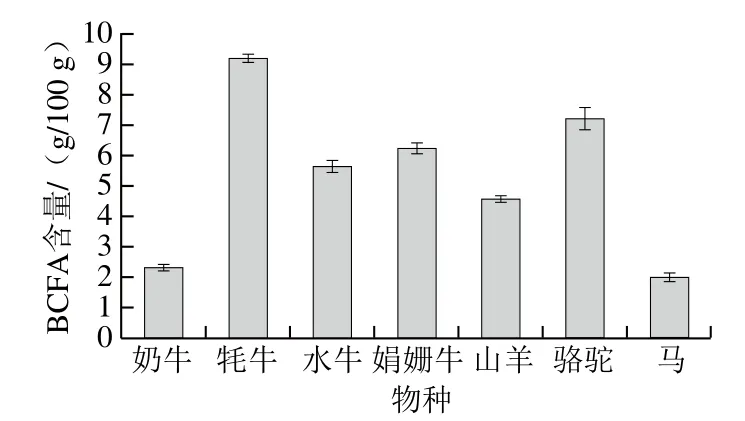
圖1 不同物種乳中BCFA含量[10]Fig. 1 Contents of BCFAs in milk from different species[10]
陳韜等[16]對不同品種的山羊肉脂中BCFA進(jìn)行檢測發(fā)現(xiàn),其BCFA含量均在4.35~5.15 g/100 g范圍內(nèi),且另有研究表明羊肉中的BCFA大多都是iso-BCFA[17]。Aldai等[18]在對加拿大零售牛肉的調(diào)查中得到其BCFA含量在1.25~1.82 g/100 g之間。Ran-Ressler等的研究也表明牛肉中的BCFA含量較低,在1.60~1.89 g/100 g,且iso-BCFA含量高于anteiso-BCFA[19]。Wang Donghao等[20]對意大利臘腸和牛肉中的BCFA進(jìn)行檢測,結(jié)果表明,牛肉中C17:0和C18:0的BCFA占比更高,且牛肉中的BCFA含量顯著高于豬肉。各類乳制品中BCFA的含量如表2所示,其中總BCFA質(zhì)量分?jǐn)?shù)最高的是綿羊奶酪,可達(dá)2.73%,最低是淡奶油(1.37%)。除冰淇淋外,其他乳制品中均是anteiso-BCFA質(zhì)量分?jǐn)?shù)高于iso-BCFA,anteiso-C15:0在大多乳制品如奶酪、酸奶和黃油等中的含量最高,但是在冰淇淋中則是anteiso-C17:0含量最高[19]。由此可見,在畜產(chǎn)品源中,牦牛乳中的BCFA含量最高。在乳及乳制品中anteiso-BCFA含量相對較高,而肉制品中則是iso-BCFA的含量相對較高。在畜產(chǎn)品中BCFA的分布較為廣泛且含量相對其來源產(chǎn)品較高。
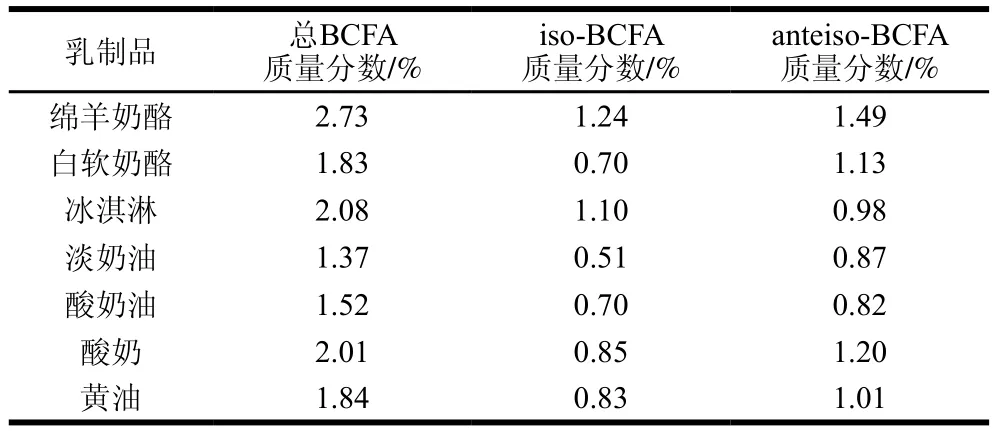
表2 不同乳制品中的BCFA含量[19]Table 2 BCFA contents in different dairy products[19]
1.3 BCFA的水產(chǎn)品源性分析
水產(chǎn)品中含有豐富的不飽和脂肪酸,BCFA在不同種類的魚及不同組織如魚肉制品、魚油、魚皮、內(nèi)臟中的分布及含量亦存在差異。Yan Yuanyuan等[8]分析了金槍魚、鳀魚、巴沙魚、古比魚和鮭魚5 種海魚油中BCFA的含量及位置分布,其中巴沙魚油的BCFA質(zhì)量分?jǐn)?shù)最低,為0.19%,古比魚油的BCFA質(zhì)量分?jǐn)?shù)可高達(dá)2.94%。魚油中的BCFA絕大部分都是iso-BCFA,但甘油三酯Sn-2位主要分布的是anteiso-BCFA。Wang Donghao等[21]對在美國東北部捕獲的27 種淡水魚進(jìn)行了脂肪酸含量的檢測。在這27 種淡水魚中,BCFA的平均含量為其可食用魚肉總脂肪酸的1%左右。在這27 種魚類中,魚皮中的BCFA含量明顯高于魚肉組織,且BCFA的主要種類為iso-C15:0、anteiso-C15:0、iso-C16:0、iso-C17:0和anteiso-C17:0。馮大偉等[22]通過氣象色譜-質(zhì)譜聯(lián)用技術(shù)分析比較了鯉魚、魷魚和鱈魚皮中的脂肪酸,其中BCFA在3 種魚皮的魚油中均有少量存在,尤其3,7,11,15-四甲基十六烷酸在鱈魚皮中質(zhì)量分?jǐn)?shù)高達(dá)5.29%。樓喬明等[23]對海洋生物中的3 種多支鏈脂肪酸進(jìn)行了分析,結(jié)果表明多BCFA在海洋生物中的質(zhì)量分?jǐn)?shù)均較低,一般都在0.1%以下,其中內(nèi)臟中的含量明顯高于肌肉。孫麗霞等[24]采用超聲提取法從羅非魚內(nèi)臟中提取油脂,其中BCFA質(zhì)量分?jǐn)?shù)為0.94%,iso-C17:0的含量占總BCFA的一半。通過BCFA的水產(chǎn)品源分析發(fā)現(xiàn),海魚中的BCFA含量高于淡水魚;不同組織相比較,魚皮中的BCFA含量相對最高,內(nèi)臟次之,肌肉組織相對最少。
1.4 BCFA的其他食品源性分析
由于亞洲國家對乳制品消費(fèi)相對較低,所以Wang Donghao等[25]推測發(fā)酵食品可能是亞洲食品中BCFA的來源,并對納豆、蝦醬、魚露、味噌、泡菜和豆豉進(jìn)行了BCFA檢測。其中,納豆中的anteiso-C15:0含量最高,可達(dá)總BCFA的1/3。蝦醬中的iso-C17:0質(zhì)量分?jǐn)?shù)最高,為0.5%。以上幾種食品中的iso-BCFA含量遠(yuǎn)遠(yuǎn)高于anteiso-BCFA,且BCFA碳原子個數(shù)均在15~18之間,其中蝦醬中還含有iso-C20:0和iso-C24:0。作為發(fā)酵食品的一種,納豆中的BCFA含量與乳脂相當(dāng),平均值為0.6 mg/g。與食品本身的成分相比,發(fā)酵可以顯著增加BCFA的含量。因此,發(fā)酵食品可為素食主義者,即攝入牛奶、牛肉和其他反芻動物產(chǎn)品較少的人群提供一定量的BCFA。
2 BCFA的功能活性及作用機(jī)理
2.1 抗炎作用及其機(jī)理
BCFA具有很好的抗炎功能,其抗炎機(jī)制如圖2所示。當(dāng)脂多糖激活細(xì)胞表面的Toll樣受體(toll-like receptor 4,TLR-4),進(jìn)而激活核因子(nuclear factor,NF)-κB時,BCFA可直接與NF-κB相互作用,抑制其表達(dá),從而降低白細(xì)胞介素-8(interlukin-8,IL-8)的合成[26]。Yan等[27]研究了BCFA對人腸道上皮細(xì)胞株(Caco-2)產(chǎn)生IL-8和NF-κB的影響。通過用特定的BCFA預(yù)處理細(xì)胞,然后用脂多糖刺激使細(xì)胞產(chǎn)生炎癥。結(jié)果表明,anteiso-BCFA和iso-BCFA都能降低白細(xì)胞介素的抗炎基因表達(dá)。而且,在經(jīng)脂多糖刺激的Caco-2細(xì)胞中,anteiso-BCFA的炎癥抑制效果比iso-BCFA更好。該研究首次證明了BCFA對人腸道上皮細(xì)胞有明顯的抗炎作用。
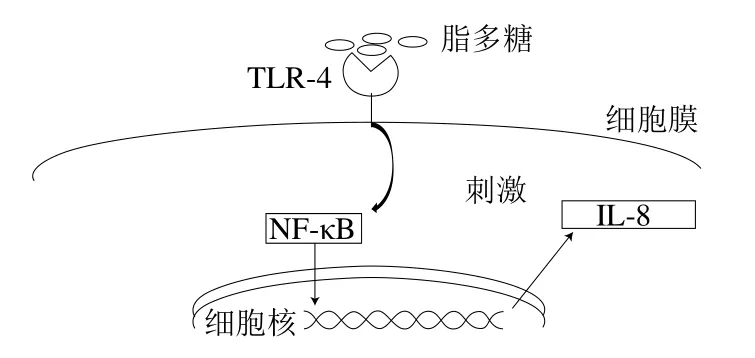
圖2 BCFA抗炎機(jī)理[27]Fig. 2 Anti-inflammatory mechanism of BCFAs[27]
新生兒壞死性小腸結(jié)腸炎(neonatal necrotizing enterocolitis,NEC)是一種常見的死亡率極高的新生兒胃腸道急癥。早產(chǎn)、腸道發(fā)育、喂養(yǎng)方式以及腸道菌群定植為影響NEC的主要因素[28]。與母乳喂養(yǎng)的早產(chǎn)兒相比,配方奶粉喂養(yǎng)的嬰兒發(fā)生NEC的概率更高[29],這可能是因?yàn)榇蠖鄶?shù)配方奶粉不含有BCFA。在妊娠后期,隨著胎兒吞食含有BCFA的皮脂量增加,其NEC的發(fā)病率下降。因此推測NEC的發(fā)生與BCFA的缺乏有關(guān)。Ran-Ressler等[30]實(shí)驗(yàn)發(fā)現(xiàn)BCFA喂養(yǎng)的幼鼠回腸中的抗炎細(xì)胞因子IL-10含量增加了3 倍,BCFA可使NEC發(fā)病率降低50%以上,驗(yàn)證了攝入BCFA能降低NEC發(fā)病率的假設(shè)。此外,BCFA會被回腸磷脂、血清和肝臟組織選擇性吸收,攝取BCFA的幼鼠腸道微生物群組成發(fā)生了改變。其枯草芽孢桿菌和銅綠假單胞菌的數(shù)量明顯增加,系統(tǒng)地改變幼鼠盲腸微生物群。由此可見,BCFA對預(yù)防NEC具有相當(dāng)明顯的作用。
2.2 抗癌作用及其機(jī)理
有研究表明BCFA具有明顯的抗癌活性,可通過抑制細(xì)胞癌的生長、抑制脂肪合成酶以及誘導(dǎo)癌細(xì)胞凋亡等途徑表達(dá)。Yang Zhenhua等[31]利用大豆發(fā)酵產(chǎn)物13-甲基十四烷酸(13-methyltetradecanoic acid,13-MTD)治療腫瘤細(xì)胞,2 h后即觀察到腫瘤細(xì)胞的死亡,且在低質(zhì)量濃度(10.03 μg/mL)下就可有效致死腫瘤細(xì)胞。尹心寶等[32]研究表明13-MTD有很強(qiáng)的誘導(dǎo)膀胱癌細(xì)胞凋亡的作用,其能明顯抑制膀胱癌T24細(xì)胞的生長,且質(zhì)量濃度在35~140 mg/L范圍內(nèi)增加時,細(xì)胞抑制率逐漸增大;不同的處理時間相比,24 h處理組的細(xì)胞抑制率高于12 h處理組,因此13-MTD的抑癌特性具有時間與濃度依賴性。翁繩美[33]對13-MTD誘導(dǎo)人早幼粒細(xì)胞白血病細(xì)胞HL60凋亡進(jìn)行研究,結(jié)果表明,在藥物處理24 h時其半抑制質(zhì)量濃度為20.5 μg/mL,而且13-MTD對HL60的抑制和殺滅作用有明顯的劑量依賴性。綜上,13-MTD主要是通過誘導(dǎo)細(xì)胞凋亡而有效抑制腫瘤細(xì)胞生長,且沒有明顯的毒副作用,是一種潛在的腫瘤化療藥物。
由于腫瘤細(xì)胞可表達(dá)高水平的脂肪酸合成酶,所以脂肪酸合成酶可作為抗癌藥物開發(fā)的靶點(diǎn)。對脂肪酸合成酶的抑制作用可以觸發(fā)細(xì)胞凋亡的信號轉(zhuǎn)導(dǎo)通路,從而導(dǎo)致細(xì)胞死亡。Wongtangtintharn等[34]證明了BCFA對人乳腺癌細(xì)胞中的抗腫瘤活性是由于BCFA對脂肪酸合成酶有直接的抑制作用,并且可以通過降低前體酶活性關(guān)系來降低脂肪酸的合成。在鏈長與抗腫瘤活性關(guān)系的研究中,iso-C16:0的活性最高,并且從C16:0開始,隨著鏈長的增加或減少,其抗腫瘤活性均降低。BCFA對癌細(xì)胞的細(xì)胞毒性與共軛亞油酸相當(dāng)。袁錦瑩等[35]從牦牛酥油中提純出的BCFA對人乳腺癌細(xì)胞的抑制作用與其結(jié)果一致。
對于BCFA的抑癌機(jī)理,Wright等[36]發(fā)現(xiàn)動脈注射含12-甲基十四烷酸(13-methyltetradecanoic acid,12-MTA)的靶向劑可顯著抑制兔肌肉VX2鱗狀細(xì)胞癌的生長,抑制機(jī)理為12-MTA顯著降低了5-脂氧合酶(lipoxygenase,5-LOX)(一種前致癌物)在腫瘤細(xì)胞內(nèi)的水平,同時增加了環(huán)氧合酶15-LOX-2的活性,從而導(dǎo)致了其產(chǎn)物羥基乙酸(一種抗癌化合物)水平的顯著增高。這是首次報(bào)道BCFA(如12-MTA)的靶向動脈輸送可能是治療實(shí)體腫瘤的一種潛在新療法。Wongtangtintharn等[37]進(jìn)一步研究表明13-MTD是通過非依賴性半胱天冬酶途徑來誘導(dǎo)乳腺癌細(xì)胞的凋亡,其主要破壞線粒體的完整性,從而阻止癌細(xì)胞的增殖[32]。12-MTA也是通過半胱天冬酶途徑抑制前列腺PC3癌細(xì)胞的增殖[38]。Lin Tianxin等[39]發(fā)現(xiàn)13-MTD通過引起線粒體功能障礙,使細(xì)胞色素c從線粒體中釋放出來,同時激活半胱天冬酶的活性,從而導(dǎo)致細(xì)胞凋亡抑制膀胱癌細(xì)胞的增殖與生存能力。
2.3 防治缺血及再灌注損傷
Faung等[40]研究表明13-MTD有很強(qiáng)的細(xì)胞溶解活性,且對血小板的溶解作用有濃度依賴性。高濃度(>60 μmol/L)的13-MTD可引起聚集的血小板溶解,其溶解作用是通過膜面積的擴(kuò)張引起細(xì)胞裂解實(shí)現(xiàn);低濃度(40 μmol/L)時13-MTD可以抑制血小板的聚集和黏附功能。綜上,13-MTD可以降低血液黏滯度,改善缺血組織的微循環(huán),從而減輕再灌注時的無復(fù)流現(xiàn)象,對缺血再灌注損傷起到一定的保護(hù)作用,同時可防止血流灌通后的再狹窄。另有研究表明BCFA可以抑制5-LOX的活性,從而減少致炎因子的產(chǎn)生,有利于減輕因缺血再灌注損傷導(dǎo)致的組織炎癥[38]。因此,BCFA在血管內(nèi)能抑制血小板和白細(xì)胞功能,在血管外可抑制5-LOX活性,降低細(xì)胞膜通透性,從而減輕水腫及炎癥反應(yīng)[41]。Yu Juan等[42]研究表明13-MTD可立即減輕局灶性缺血再灌注損傷。不同劑量的13-MTD均能有效減少再灌注后24 h的梗死體積,減輕腦水腫,增加堿性成纖維細(xì)胞生長因子和血管內(nèi)皮生長因子的mRNA和蛋白質(zhì)表達(dá),發(fā)揮神經(jīng)保護(hù)作用,促進(jìn)缺血半暗帶內(nèi)皮細(xì)胞增殖和血管生成。何宏星等[43]認(rèn)為各劑量的13-MTD對不同時間大鼠局灶性腦缺血所致的腦損傷均有保護(hù)作用,缺血6 h時,80 mg/kg的13-MTD療效較為明顯。
13-MTD具有保護(hù)神經(jīng)細(xì)胞形態(tài)以及減少其超微結(jié)構(gòu)損傷的作用,可減少神經(jīng)元的凋亡,從而提高細(xì)胞的存活率[44]。余涓等[45]研究了13-MTD對氧反常誘導(dǎo)大鼠胚腦皮質(zhì)神經(jīng)元凋亡和形態(tài)學(xué)損傷的保護(hù)作用,結(jié)果表明,模型組神經(jīng)元胞體腫脹,突起收縮甚至脫落,神經(jīng)細(xì)胞存活率顯著下降。與模型組相比,不同劑量的13-MTD可有效改善神經(jīng)元形態(tài),使其細(xì)胞結(jié)構(gòu)明顯恢復(fù),且20 mg/L的13-MTD改善效果最為顯著。因此,13-MTD對氧反常誘導(dǎo)的大鼠胚腦皮質(zhì)神經(jīng)元損傷具有明顯的保護(hù)作用。
2.4 降脂作用及其機(jī)理
肥胖是由于一系列的代謝紊亂引起的,主要表現(xiàn)為血脂異常,即低密度脂蛋白、總膽固醇和甘油三酯水平的升高以及高密度脂蛋白水平的下降[26]。棕櫚酸的過量消耗可誘導(dǎo)高脂血癥、胰島素抵抗等代謝疾病,有研究表明BCFA可通過上調(diào)硬脂酰輔酶A去飽和酶的表達(dá),促進(jìn)9-十六碳烯酸的形成,進(jìn)一步驅(qū)動9-十六碳烯酸參與三酰基甘油的合成,從而緩解由棕櫚酸誘導(dǎo)的內(nèi)質(zhì)網(wǎng)應(yīng)激和細(xì)胞凋亡[46]。在人體脂肪酸中,BCFA和奇數(shù)碳鏈脂肪酸的含量雖較低,但對血脂有很大的調(diào)節(jié)功能。Mika等[47]通過氣相色譜-質(zhì)譜法對超重患者血清中的奇數(shù)碳鏈脂肪酸和支鏈脂肪酸進(jìn)行了綜合性研究。其中超重患者血清中的BCFA水平明顯低于對照組(圖3)。而且,血清中的總BCFA含量與胰島素、甘油三酯、18∶1/18∶0去飽和指數(shù)呈負(fù)相關(guān)。該研究首次證明了BCFA與肥胖之間的關(guān)系,在肥胖患者中,較低的BCFA含量可能導(dǎo)致血清中的甘油三酯水平升高。Su Xiong等[48]對肥胖患者術(shù)前和術(shù)后1 年的BCFA含量進(jìn)行檢測,表明肥胖患者在減肥后BCFA的含量增加,其結(jié)果與Mika等[47]的結(jié)果相一致。盡管BCFA在人體血液中的含量較少,但適當(dāng)攝入可能會產(chǎn)生許多健康益處,與通常補(bǔ)充的n-3多不飽和脂肪酸類似。
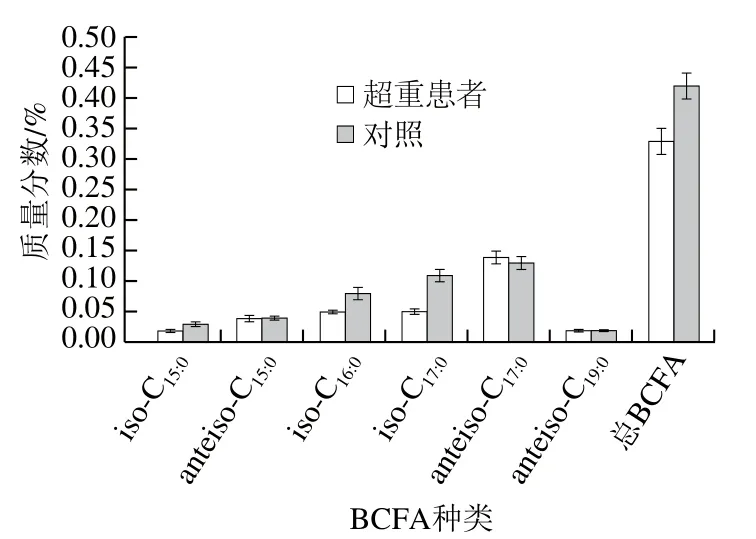
圖3 超重患者和對照組中BCFA含量[47]Fig. 3 Contents of BCFAs in overweight patients and control group[47]
Heimann等[49]對BCFA調(diào)節(jié)初級脂肪細(xì)胞的糖脂代謝進(jìn)行了研究,結(jié)果表明,BCFA對基礎(chǔ)脂肪生成和胰島素刺激的脂肪生成均有抑制作用。李姣[50]探討了BCFA的降脂作用及其機(jī)制,BCFA降低肝細(xì)胞內(nèi)脂質(zhì)含量的原因主要是抑制肝臟細(xì)胞部分脂肪合成關(guān)鍵酶的表達(dá),以及促進(jìn)脂肪分解關(guān)鍵酶表達(dá)。肝臟病變的重要原因之一就是脂肪沉積,非酒精性脂肪肝就是一種典型的由于肝細(xì)胞脂肪堆積誘導(dǎo)形成的病癥。賀宇佳等[51]通過肝細(xì)胞脂肪沉積模型探討了BCFA對肝細(xì)胞脂肪沉積的影響,研究結(jié)果與李姣[50]的一致,且BCFA對肝細(xì)胞脂肪沉積的抑制作用呈劑量依賴性。因此,BCFA可潛在改善脂肪代謝,減少肝細(xì)胞脂肪累積。
3 結(jié) 語
BCFA在人體中的含量雖少,卻發(fā)揮著重要的作用,尤其對于嬰幼兒的腸道健康。BCFAs在嬰兒配方奶粉中的添加對預(yù)防新生兒NEC至關(guān)重要,因此,BCFAs是嬰兒配方奶粉中不可缺少的營養(yǎng)素。最新研究表明,利用尿素絡(luò)合法從羊毛脂游離脂肪酸中得到的BCFA濃縮物主要成分是anteiso-C15:0和anteiso-C17:0,說明BCFA濃縮物的組成和類型與母乳脂肪酸相似,在提高嬰幼兒配方奶粉質(zhì)量方面具有潛在的應(yīng)用前景[52]。
目前關(guān)于BCFA的應(yīng)用研究較為少見,通過對其功能特性的研究可以看出BCFA的各種生理功能對治療某些疾病方面十分有益,說明BCFA有很大的應(yīng)用空間,尤其在嬰幼兒特醫(yī)食品以及抗癌方面。反芻動物制品是人們攝取BCFA的主要來源,但我國對于乳制品的消費(fèi)量相較于歐美地區(qū)低很多。所以,對于含BCFA的功能性食品或特殊醫(yī)療用途配方食品應(yīng)受到相關(guān)研究者的重視。另外,我國對于BCFA的研究還較為匱乏,其生理作用機(jī)制、代謝途徑以及不同結(jié)構(gòu)對其生理功能的影響等方面都尚不清楚。因此,BCFA這一領(lǐng)域還具有十分廣闊的研究前景。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
國際商務(wù)財(cái)會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
