發酵十字花科蔬菜中硫代葡萄糖苷代謝研究進展
2022-01-06 02:32:00趙吉春譚正衛顏鑫藝周海燕雷小娟
食品科學 2021年23期
趙吉春,余 潔,譚正衛,顏鑫藝,周海燕,雷小娟,明 建,*
(1.西南大學食品科學學院,重慶 400715;2.重慶市渝北中學校,重慶 401120)
大白菜(Brassica rapaspp.Pekinensis)、甘藍(Brassica oleraceaL. var.capitata)、紫甘藍(Brassica oleraceavar.capitataf. rubra)等十字花科蔬菜是制作四川泡菜、東北酸菜、德國酸菜和韓國泡菜等各種發酵蔬菜制品的主要原材料[1-4]。這些蔬菜不僅含有豐富的膳食纖維、維生素、鈣、鐵、鋅等礦物元素,還具有能夠促進人體健康的各類植物化學物質,例如硫苷、黃酮和花青素等[5-8]。
蔬菜的發酵依賴于原材料攜帶的乳酸菌以及人工接種發酵劑的生長繁殖活動。在異型發酵乳酸菌和同型發酵乳酸菌交替作用下,蔬菜中糖類被轉化為乳酸、乙酸等代謝產物,降低了體系pH值。一方面,發酵抑制腐敗菌生長,延長蔬菜產品的保質期;另一方面,發酵可產生有機酸、氨基酸等營養成分以及特征性風味和滋味[8]。此外,乳酸菌發酵也促進了植物化學物質的生物轉化。
硫苷是十字花科蔬菜中一類重要的特征性含硫化合物,但自身活性較低,能夠在黑芥子酶催化下發生降解。蔬菜發酵過程中,硫苷被降解生成異硫氰酸鹽、腈類、環腈類等活性代謝產物,且不同產物的生理活性存在較大差異,其中異硫氰酸鹽的活性最高。研究顯示乳酸菌對硫苷具有轉化能力,且存在菌株差異性,并受到多種外在因素影響。本文總結了硫苷及其代謝產物生理功能的最新研究進展,重點研究分析了各類發酵蔬菜中乳酸菌對硫苷的代謝作用,比較不同種屬乳酸菌對發酵蔬菜硫苷代謝產物的影響差異,并討論了鹽含量、pH值、金屬離子以及加熱處理等因素對硫苷代謝的影響,為調控發酵蔬菜中硫苷向高活性的代謝產物方向轉化提供理論依據。
1 硫苷概述
硫苷是一類水溶性含硫的次生代謝產物,其結構主要由β-D-硫葡萄糖基、硫化肟基和可變氨基酸衍生側鏈R基團構成(圖1)。目前,已從十字花科蔬菜中鑒定出多達137 種硫苷化合物[9]。根據R基團結構不同,硫苷可以分為脂肪族硫苷(R基團為甲硫氨酸、丙氨酸、擷氨酸、亮氨酸和異亮氨酸)、芳香族硫苷(R基團為苯丙氨酸和酪氨酸)和吲哚族硫苷(R基團為色氨酸),其中第一類約占已發現硫苷的50%[10]。十字花科蔬菜中常見硫苷有3-丁烯基硫苷、2-羥基-3-丁烯基硫苷、蘿卜硫苷、黑芥子苷和4-羥基-3-吲哚甲基硫苷等,具體到特定蔬菜,其硫苷組成與含量因栽培品種、栽培條件、采后貯藏條件以及蔬菜部位而異[7,11]。例如,大白菜硫苷以3-丁烯基硫苷和4-戊烯基硫苷為主,而西蘭花的主要硫苷有新葡萄糖蕓苔素、蕓苔葡糖硫苷和蘿卜硫苷[7,12](表1)。
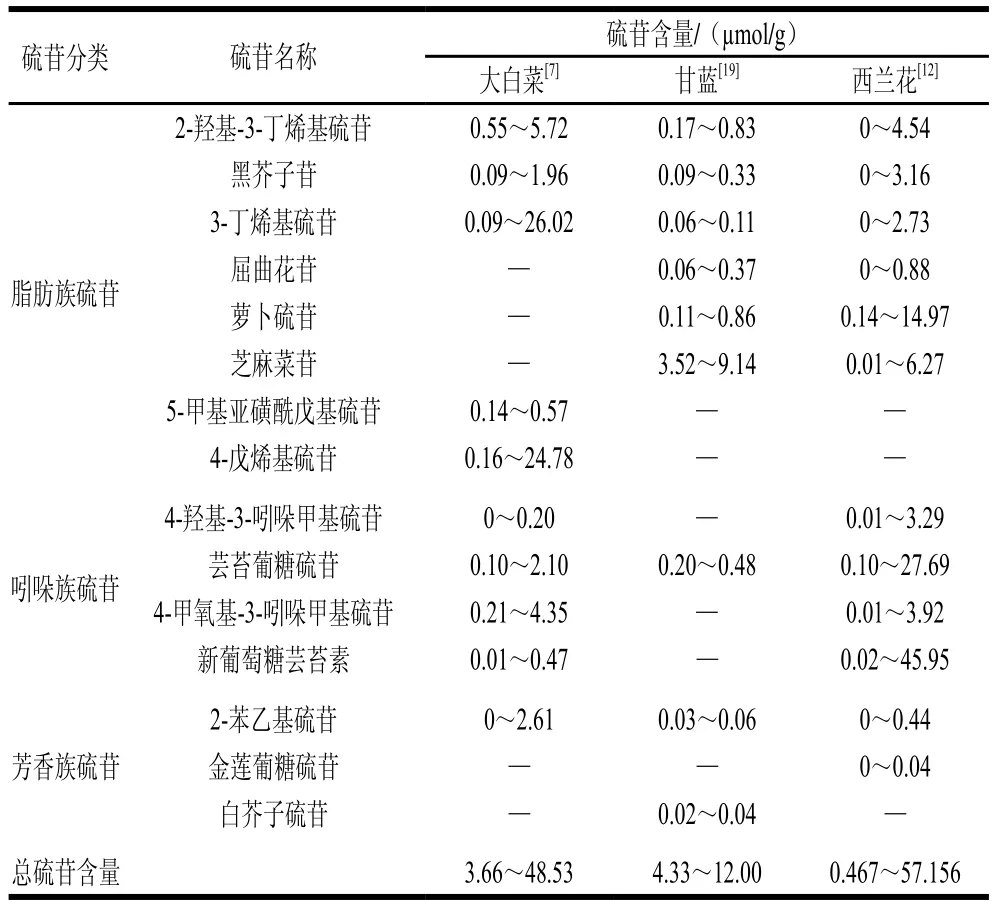
表1 大白菜、甘藍和西蘭花中硫苷種類與含量Table 1 Types and contents of glucosinolates in Chinese cabbage,cabbage and broccoli
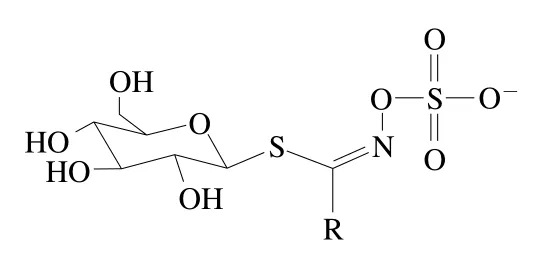
圖1 硫苷基本結構Fig. 1 Basic structure of glucosinolates
黑芥子酶存在于所有含有硫苷的植物組織中,可以特異性地裂解硫連接的糖苷。在完整蔬菜組織中,硫苷存在于S細胞,而黑芥子酶位于相鄰的芥子酶細胞,彼此相互分隔。當蔬菜遭受切割、咀嚼等機械損傷時,完整組織結構被破壞,黑芥子酶和硫苷相互接觸發生反應。硫苷自身難以被人體直接吸收代謝,攝入體內后,絕大多數硫苷以完整形式通過上消化道而進入大腸,被腸道微生物代謝利用[13]。在發酵蔬菜中乳酸菌也可以代謝硫苷,提高硫苷化合物的生物活性。硫苷及其代謝產物也是發酵蔬菜中辛辣味、苦味等特征性風味的主要物質基礎之一[14-15]。眾多研究已表明硫苷主要代謝產物異硫氰酸鹽具有抗炎、提高抗氧酶活性、調節腸道菌群、降低罹患癌癥風險、降低糖尿病風險、緩解肥胖等多種生理功能[16-18]。
2 硫苷生理功能
目前,關于硫苷及其代謝產物生理功能的研究主要集中于抗癌和抗腫瘤作用,特別是拮抗結直腸癌研究報道最多。其次,硫苷的抗氧化作用及其機制也受到人們關注。另外,硫苷在調節腸道菌群、應對肥胖、糖尿病等慢性疾病中發揮的積極作用近年來也逐漸被人們重視。
2.1 抗氧化活性
Maryati等[20]在西蘭花提取液中接種保加利亞乳桿菌(Lactobacillus bulgaricus)、嗜熱鏈球菌(Streptococcus thermophilus)、嗜酸乳桿菌(Lactobacillus acidophilus)和兩歧雙歧桿菌(Bifidobacterium bifidum)混合發酵劑,20%發酵劑接種量發酵24 h,發酵西蘭花的抗氧化活性達到最大,此時總酚含量降低,提示發酵產物抗氧化能力的物質基礎可能不是多酚,而是與西蘭花硫苷等活性物質具有相關性。Lee等[7]測定了62個大白菜品種提取物的總硫苷含量與抗氧化能力,結果發現硫苷粗提物的1,1-二苯基-2-三硝基苯肼自由基清除能力、羥自由基清除能力和鐵還原能力均呈現濃度依賴關系,但與總硫苷含量不具有顯著相關性。芥藍的植物活性物質與抗氧化能力相關性分析結果顯示芥蘭的抗氧化能力與屈曲花苷、蘿卜硫苷和蕓苔葡糖硫苷等硫苷含量具有高度正相關性,提示特定硫苷對芥蘭總抗氧化能力具有貢獻[21]。
現在普遍認為絕大多數硫苷不具有清除自由基、中斷氧化鏈式反應等直接氧化性,其抗氧化能力主要表現在間接抗氧化性。例如,蘿卜硫苷是西蘭花中含量最高的硫苷之一,其代謝產物蘿卜硫素是一種重要的異硫氰酸鹽,通過激活核轉錄因子2(nuclear transcription factor 2,Nrf2)信號通路而誘導II相酶,例如谷胱甘肽-S-轉移酶、谷胱甘肽還原酶和NAD(P)H:醌氧化還原酶(NAD(P)H: quinone oxidereductase,NQO1)、環氧化物酶和血紅素加氧酶,這些酶有助于細胞抵御親電試劑的攻擊從而緩解氧化損傷[22]。蘿卜硫素通過核轉錄因子2-Kelch樣環氧氯丙烷相關蛋白1(nuclear transcription factor 2-Kelch-like ECH-associating protein 1,Nrf2-keap1)系統增強了抗氧化酶活性,從而緩解幽門螺旋桿菌感染誘導的胃黏膜氧化損傷[23-24]。芝麻菜苷及其代謝產物4-甲硫基丁基異硫氰酸酯(erucin,ERN)除了具有間接抗氧化性外,還可以有效分解過氧化氫和烷基氫過氧化物,可被視為過氧化物清除能力型抗氧化劑[25]。
Cedrowski等[26-27]對蘿卜硫素和ERN可以通過誘導II相細胞保護酶而調節細胞氧化還原狀態但卻不具備自由基捕獲能力的原因進行了研究。研究結果發現,在100 ℃以下,蘿卜硫素和ERN均不能抑制亞麻酸的氧化,但是在150 ℃以上卻能增加大豆卵磷脂的氧化穩定性。結合氣相色譜-質譜檢測結果,作者推斷ERN的甲基磺酰基和蘿卜硫素的甲基亞磺酰基經140 ℃以上高溫轉化為次磺酸,與過氧自由基發生反應,阻斷自由氧化鏈式反應。甲基亞磺酰基還可能經熱降解釋放出亞磺酰基自由基,從而與烷基過氧自由基發生反應。次磺酸與過氧自由基反應也會產生亞磺酰基自由基。
2.2 調節腸道菌群
硫苷以及富含硫苷的西蘭花對腸道菌群具有調節作用。一項探索性研究顯示攝食富含硫苷的酸菜使得受試人群腸道菌群組成發生變化,與酸菜發酵相關的乳酸菌(例如植物乳桿菌(Lactobacillus plantarum)和短乳桿菌(Lactobacillus brevis))豐度顯著上升[28]。在Yanaka等[29]開展的一項安慰劑對照半開放標簽干預實驗中,慢性功能性便秘人群每日攝食20 g富含硫苷的西蘭花嫩芽或不含硫苷的苜蓿芽,持續4 周。與食用苜蓿芽相比,攝食西蘭花嫩芽可以顯著降低便秘評分系統得分,提高排便頻率,縮短排便用時,并且提高了腸道菌群中產丁酸的雙歧桿菌屬(Bifidobacterium)和ClostridiumXVIa的比例。蘿卜硫素干預結腸癌的研究也顯示攝入西蘭花芽持續2 周,能提高人體腸道菌群中雙歧桿菌屬和ClostridiumXVIa的比例[30]。
攝入蘿卜硫苷可以降低高脂膳食小鼠糞便菌群的厚壁菌門(Firmicutes)與擬桿菌門(Bacteroidetes)之比,減少與高脂飲食呈正相關的菌群豐度,例如毛螺菌科(Lachnospiraceae)和脫硫弧菌科(Desulfovibrionaceae)[31],這與富含硫苷的西蘭花對健康人群腸道微生物的影響一致[32]。此外,小鼠喂養西蘭花實驗也顯示攝入西蘭花可以顯著提高腸道菌群厚壁菌門和擬桿菌門的相對豐度,降低變形菌門(Proteobacteria)豐度,提高盲腸和結腸內容物類黑芥子酶活性以及異硫氰酸鹽含量[33]。蘿卜硫苷攝入可以恢復N-丁基-N-(4-羥基丁基)-亞硝胺誘導的膀胱癌小鼠模型腸道菌群紊亂,顯著增加脆弱擬桿菌(Bacteroides fragilis)和Clostridium cluster I的比例,提高結腸中丁酸水平,并通過上調緊密連接蛋白和胰高血糖素樣肽2(glucagon-like peptide 2,GLP2)的表達來修復結腸和盲腸黏膜上皮的損傷[34]。
2.3 抗癌作用
流行病學調查顯示攝入富含硫苷的十字花科蔬菜與肺癌、膀胱癌、結腸癌、前列腺癌等癌癥發生率存在負相關關系,攝入高含量的十字花科蔬菜可以將結直腸癌、胃癌罹患風險分別降低8%和19%[35-38]。一項超過8 000 例樣本的文獻薈萃分析也顯示十字花科蔬菜攝入與結腸腺瘤發生風險降低存在相關性[39]。在拮抗癌癥方面,硫苷對結直腸癌的作用尤為受到研究者的關注。
通過給予小鼠化學致癌劑氧化偶氮甲烷(15 mg/kg)進行3~6 次造模,同時每日飼喂蘿卜硫苷,連續24 周,結果顯示蘿卜硫苷抑制了氧化偶氮甲烷誘導的異常隱窩病灶(aberrant crypt foci,ACF)和結腸腫瘤的形成。患有結腸腺瘤的病人每兩天攝入一次富含硫苷的50 g生西蘭花芽,持續6個月,減少了結腸ACF數量[30]。Ho等[18]通過質粒將源于辣根的黑芥子酶基因導入腸道共生菌Escherichia coliNissle 1917,并經由YebF蛋白分泌標簽將黑芥子酶分泌至胞外,從而將黑芥子苷轉化為異硫氰酸烯丙酯(allyl isothiocyanate,AITC)。同時,該工程菌被導入了源于解沒食子酸鏈球菌(Streptococcus Gallolyticus)的類組蛋白(histone-like protein A,HlpA)基因,獲得與結腸癌細胞表面的硫酸乙酰肝素蛋白聚糖特異性地結合的能力,并在細胞表面分泌黑芥子酶將宿主攝入的硫苷轉化為異硫氰酸鹽,從而進入癌細胞。工程菌和硫苷組合在體外實驗中可以將超過95%的鼠源和人源結直腸癌細胞系殺死,而對人源胃癌細胞系、乳腺癌細胞系和食管平滑肌細胞系沒有抑制作用。工程菌和西蘭花/黑芥子苷膳食聯合喂養顯著降低了結直腸癌小鼠模型的腫瘤發生的風險,結直腸區域的腫瘤數量減少了75%以上。
2.4 緩解代謝綜合征
近年來,十字花科蔬菜硫苷及其活性代謝產物在降低脂肪積累、提高胰島素敏感性、降低炎癥等方面表現出良好特性,因而在緩解代謝綜合征方面的潛力得到了研究者的關注[40]。Xu Xinxing等[31]在飼料中添加西蘭花凍干粉或蘿卜硫苷,喂養高脂膳食小鼠8 周,發現蘿卜硫苷喂養可以有效地抑制小鼠體質量、肝臟質量增加,減少附睪脂肪、腸系膜脂肪和腎周脂肪脂質積累,同時降低了氧化應激水平,上調肝臟中與脂質代謝相關的過氧化物酶體增殖物激活受體α(peroxisome proliferator-activated receptor α,PPARα)、肉堿棕櫚酰轉移酶-1(carnitine palmitoyl transterase-1,CPT1)、過氧化物酶體酰基輔酶A氧化酶(acyl-coenzyme A oxidase,ACOX)基因表達,而飼喂西蘭花對高脂膳食小鼠脂質代謝的改善作用要弱于飼喂蘿卜硫苷的作用。Liu Yaoli等[41]將蘿卜硫素(10 mg/kg)以腹腔注射方式每日給予高脂膳食誘導的肥胖小鼠,連續30 d后,減少了小鼠體質量增加和脂肪細胞增大,并且抑制了脂肪形成關鍵基因表達和脂質在前體脂肪細胞C3H10T1/2中積累。同時,蘿卜硫素也促進了肥胖小鼠白色脂肪組織棕色化,降低了細胞和線粒體氧化應激,提高了胰島素敏感度。
此外,在肥胖誘導的炎癥反應中,Toll樣受體(Toll-like receptors,TLR)-骨髓樣分化因子88(myeloid differentiation primary response 88,MyD88)信號通路居于中心位置。兩種硫苷物質異硫氰酸苯乙酯(phenethyl isothiocyanate)和3,3’-二吲哚甲烷(3,3’-diindolylmethane)膳食飼喂高脂高膽固醇膳食的肥胖小鼠,顯著降低了小鼠脾臟TRR2/4、MyD88、IL-6、NF-κB和環氧化酶2(cyclooxygenase 2,COX-2)mRNA表達水平,即兩種硫苷物質通過調節TLR-MyD88信號通路緩解了脾臟炎癥[42]。
硫苷雖然具有諸多生理功能,但其潛在安全問題同樣也應受到重視。硫苷自身不具有毒性,但其降解產物并非如此。例如,菜籽粕中硫苷代謝產物噁唑烷硫酮會損傷甲狀腺對碘的吸收能力,導致血液甲狀腺素濃度下降;硫氰酸鹽會競爭性抑制甲狀腺吸收碘,導致甲狀腺功能減退;腈類會增加肝臟抗氧化酶和脫毒酶的代謝活性,導致肝臟發生腫大增生[43]。菜籽粕含有豐富蛋白質,是一種理想的動物飼料。然而,硫苷是菜籽粕的抗營養因子,因此需要對菜籽粕進行脫毒處理。Latimer等[44]考察了5 種不同硫苷的代謝產物腈對人和牛肝細胞毒性,受試化合物最大濃度為2 mmol/L,孵育24 h條件下,沒有影響HepG2細胞和牛干細胞活性,也沒有抑制三磷酸腺苷結合盒轉運體(ATP-binding cassette transporter)G2轉運熒光基質的能力。蔬菜中硫苷含量較低,對其硫苷的安全問題目前關注較少,在未來研究中需要加強發酵蔬菜中硫苷安全性的研究。
3 發酵蔬菜中硫苷的代謝
十字花科蔬菜組織完整時,硫苷與黑芥子酶彼此相互隔離,植物組織中硫苷處于惰性狀態[45]。制作發酵蔬菜時,蔬菜組織經受切割、加工,硫苷與黑芥子酶接觸,硫苷在黑芥子酶作用下轉化成異硫氰酸鹽、硫氰酸鹽、環腈類、腈類、噁唑烷等代謝產物[46]。在發酵過程中,除了蔬菜內源性黑芥子酶代謝作用,蔬菜自身攜帶的乳酸菌以及添加的發酵劑同樣在硫苷代謝方面發揮著重要的作用。
截至目前,已知具有硫苷代謝能力的微生物主要來源于土壤、人體糞便和發酵食品[47-51]。其中,來自發酵蔬菜的乳酸菌主要有乳桿菌類(敏捷乳桿菌(Lactobacillus agilis)、格式乳桿菌(Lactobacillus gasseri)、嗜酸乳桿菌、干酪乳桿菌(Lactobacillus casei)、植物乳桿菌、副干酪乳桿菌(Lactobacillus paracasei))、腸膜明串珠菌(Leuconostoc mesenteroides)、雙歧桿菌(假小鏈雙歧桿菌(Bifidobacterium pseudocatenulatum)、青春雙歧桿菌(Bifidobacterium adolescentis)和長雙歧桿菌(Bifidobacterium longum))、乳酸乳球菌(Lactococcus lactis)和腸桿菌(陰溝腸桿菌(Enterobacter cloacae)和大腸桿菌(Escherichia coli))[46-47,52-55]等。
乳酸菌對十字花科蔬菜硫苷的代謝作用,可以促進植物組織硫苷物質的釋放,也提高硫苷向異硫氰酸鹽等活性代謝產物的生物轉化。乳酸菌利用蔬菜碳水化合物代謝產生乳酸、乙酸等,降低了體系pH值,也會影響硫苷的代謝以及代謝產物的生成。
3.1 乳酸菌發酵促進蔬菜中硫苷釋放
經過微生物發酵,某些硫苷從十字花科蔬菜植物組織中釋放,從而增加了含量。西蘭花泥經121 ℃、3 min高溫滅酶滅菌處理后,屈曲花苷、2-羥基-3-丁烯基硫苷和蘿卜硫苷含量低至無法定量,經植物乳桿菌和腸膜明串珠菌不同菌株接種發酵15 h,屈曲花苷、2-羥基-3-丁烯基硫苷和蘿卜硫苷的含量分別增加至16.1~56.2、24.5~65.9、54.8~358.7 μg/g,且不同菌株之間存在顯著差異[54]。這種現象出現可能原因是在高溫滅酶時,游離態硫苷也被降解,因此幾乎無法檢出,而經過乳酸菌發酵,某些微生物代謝酶可以改變西蘭花基質的理化特性,比如裂解植物細胞壁,提高了發酵西蘭花中硫苷的可提取性,使得檢出硫苷含量顯著增加。
3.2 乳酸菌發酵促進蔬菜中硫苷生物轉化
研究顯示自然發酵或乳酸菌接種發酵后,西蘭花、甘藍等十字花科蔬菜中硫苷轉化為抗壞血酸原、異硫氰酸鹽和腈類等代謝產物[53,56-57]。甘藍中主要硫苷為黑芥子苷、蘿卜硫苷、屈曲花苷和3-丁烯基硫苷,經自然發酵后,代謝形成吲哚-3-甲醇、吲哚-3-乙腈和抗壞血酸原[56]。Palani等[49]研究發現,經過5 d自然發酵,甘藍中90%硫苷降解成活性物質抗壞血酸原和吲哚-3-甲醇。蕓苔葡糖硫苷被代謝成吲哚-3-甲醇,在酸性環境下,吲哚-3-甲醇與抗壞血酸結合形成抗壞血酸原[58]。Tolonen等[57]報道在發酵2 周內,甘藍硫苷全部降解為異硫氰酸鹽、吲哚-3-甲醇、噁唑烷硫酮、烯丙基腈和腈類。此外,也有研究報道甘藍發酵還會形成3-甲基亞磺酰基丙基異硫氰酸酯(iberin,IBR)、烯丙基腈、異硫氰酸烯丙酯和蘿卜硫素等揮發性代謝產物[59](表2)。
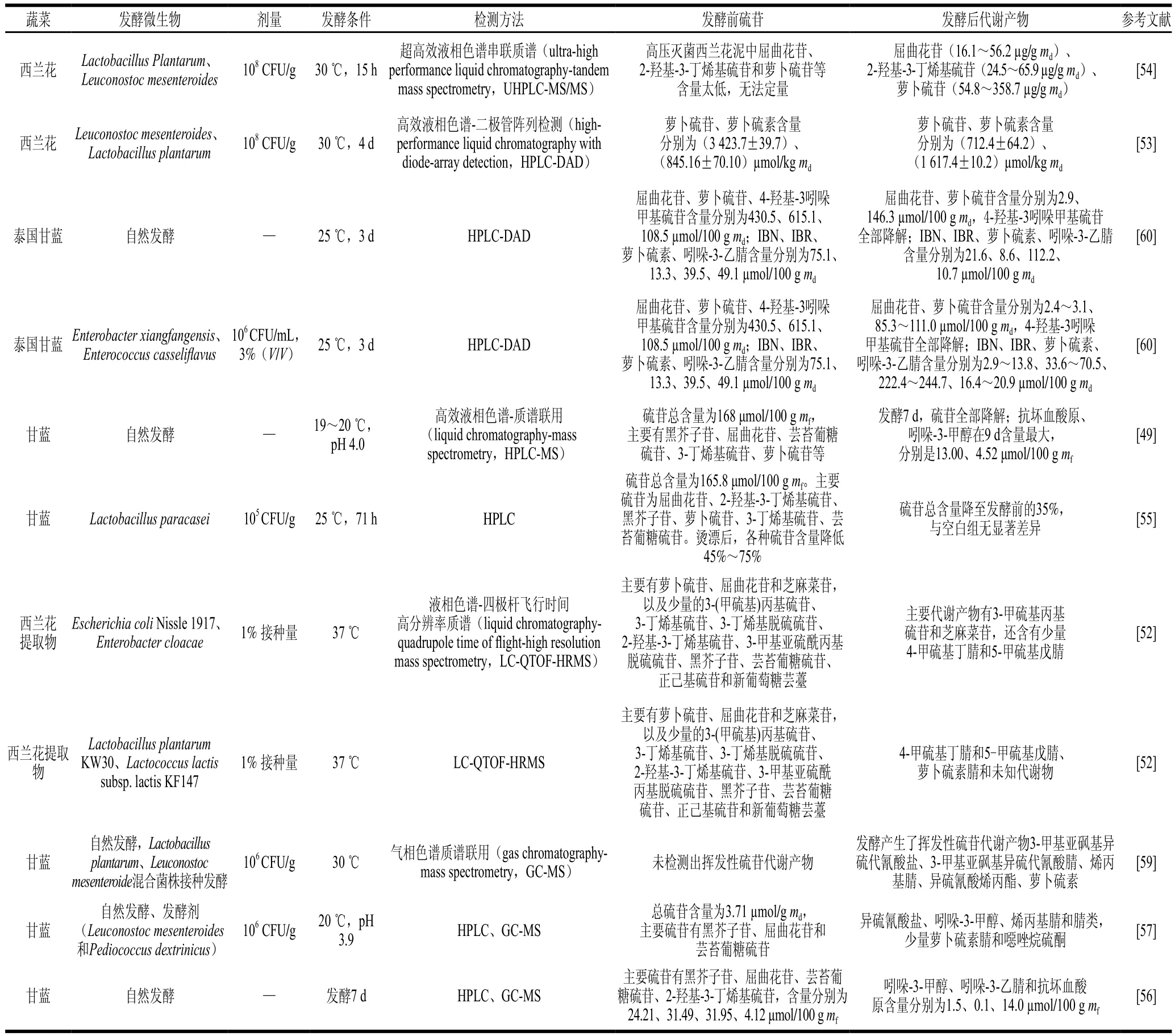
表2 發酵蔬菜中硫苷的代謝變化Table 2 Metabolism of glucosinolates in fermented vegetables
Mullaney等[52]研究發現腸桿菌科發酵將大部分西蘭花屈曲花苷和蘿卜硫苷轉化為3-甲硫基丙基硫苷和芝麻菜苷,以及少量的4-甲硫基丁腈和5-甲硫基戊腈;而植物乳桿菌、乳酸乳球菌發酵產生4-甲硫基丁腈、5-甲硫基戊腈、蘿卜硫素腈,并使其進一步代謝為未知物質,這提示微生物種屬是影響硫苷代謝產物的一個重要因素。腸膜明串珠菌作為發酵劑時,酸菜中蘿卜硫素含量顯著高于以植物乳桿菌為發酵劑的泡菜中含量[59]。香坊腸桿菌(Enterobacter xiangfangensis)4A-2A3.1(EX)和酪黃腸球菌(Enterococcus casseliflavus)SB2X2(EC)發酵泰國甘藍,硫苷代謝為4-甲基亞磺酰基丁腈(iberin nitrile,IBN)、IBR、蘿卜硫素和吲哚-3-乙腈。其中EX組IBR含量顯著高于EC組[60]。近期,一項研究顯示,腸道微生物多形擬桿菌(Bacteroides thetaiotaomicron)具有特殊操縱子是將硫苷轉化為異硫氰酸鹽的一個重要的條件,進一步說明微生物在代謝硫苷方面具有菌株特異性[46]。
4 影響發酵十字花科蔬菜硫苷代謝的因素
十字花科蔬菜硫苷代謝既與蔬菜自身硫苷種類和豐度相關,也受到發酵蔬菜體系pH值、金屬離子和特定蛋白等因素影響,同時也與蔬菜原材料前處理方法有關。
4.1 蔬菜中硫苷種類和含量差異
大白菜、甘藍、西蘭花等十字蔬菜的硫苷在種類和含量存在較大差異(表1)。即使同一物種,不同栽培品種之間的硫苷差異也十分明顯。例如,62個大白菜品種的總硫苷含量最大可相差13 倍,而具體到特定種類硫苷,差異更為明顯。以吲哚-3-甲基硫苷含量為例,80個西蘭花品種中最高含量是最低含量的270 倍。
除了蔬菜品種,蔬菜栽培條件也會造成蔬菜組織中硫苷含量的差異。Martinez-Villaluenga等[61]報道屈曲花苷、黑芥子苷和吲哚-3-甲基硫苷是甘藍中主要的硫苷,且與栽培季節(夏季或冬季)無關,但冬季收獲甘藍的吲哚-3-甲基硫苷(抗壞血酸原前體物質)含量要高于夏季甘藍。
4.2 氯化鈉含量
發酵蔬菜中氯化鈉濃度對硫苷代謝產物以及含量具有重要影響。Martinez-Villaluenga等[61]發現添加了0.5%氯化鈉的酸菜發酵一周后,抗壞血酸原含量顯著高于添加了1.5%氯化鈉的酸菜,無論甘藍栽培季節是夏天還是冬天、自然發酵還是發酵劑發酵,趨勢均一致。Pe?as等[62]研究也發現無論是自然發酵還是接種了植物乳桿菌和腸膜明串球菌混合菌株發酵7 d的酸菜,0.5%氯化鈉組酸菜的吲哚-3-甲醇和吲哚-3-乙腈的含量均顯著高于1.5%氯化鈉酸菜中的含量。東北酸菜發酵30 d,與高鹽發酵組(2.5%和3.5%)相比,低鹽發酵組(0.5%和1.5%)的4-戊烯基異硫氰酸酯、1-異硫代氰酸丁酯、3-丁烯基異硫氰酸酯等揮發性異硫氰酸鹽含量降速更快,隨著異硫氰酸鹽含量降低,硫苷的另一類代謝產物腈類含量同時增加[51]。異硫氰酸鹽和腈類的形成與體系pH值密切相關,而鹽含量會影響微生物種群的類型,進而影響其產酸能力[8]。此外,高濃度氯化鈉對黑芥子酶活性的抑制作用可能是其影響硫苷代謝的另一個途徑[63]。
在自然發酵和接種乳酸菌發酵甘藍的研究中發現,只有植物乳桿菌作為發酵時,更高鹽含量(1.5%與0.5%相比)發酵體系的IBR、烯丙基腈、蘿卜硫素等揮發性硫苷代謝產物含量會顯著增加,這可能與菌株耐受滲透壓的能力有關[59]。植物乳桿菌接種發酵的酸菜,0.5%氯化鈉組的微生物數量(包括好氧嗜常溫菌和乳酸菌)顯著高于1.5%氯化鈉的酸菜,而腸膜明串珠菌發酵酸菜的微生物數量在不同鹽含量組別間沒有差異[64]。
4.3 特殊蛋白
硫苷在黑芥子酶催化下先裂解釋放β-D-葡萄糖,生成不穩定的糖苷配基。然后,糖苷配基通過洛森重排形成異硫氰酸鹽,或者分解為腈類和環腈類等代謝產物。在黑芥子酶催化降解硫苷過程中,某些特殊蛋白起到了重要調控作用,這些蛋白主要有表皮特異硫蛋白(epithiospecifier protein,ESP)[65]、腈特異性蛋白(nitrile specifier protein,NSP)[66]、硫氰酸鹽形成蛋白(thiocyanate-forming protein,TFP)[67]和表皮特異修飾蛋白(epithiospecifier modifier protein,ESM)[68]。ESP作為黑芥子酶的非催化性輔酶因子,促進不飽和脂肪族糖苷配基形成環腈類和腈類,而TFP的存在促進生成硫氰酸鹽。NSP和TFP也會通過與不穩定糖苷配基結合生成腈類。特殊蛋白對硫苷代謝產物的影響往往與pH值相關,例如ESM在中性條件下會抑制環腈類和腈類形成,而形成異硫氰酸鹽。
4.4 金屬離子
研究表明在pH 3.5~7.5范圍內,鐵離子存在不會抑制黑芥子苷釋放葡萄糖。然而,在pH 4.5和5.5條件下,鐵離子會強烈抑制異硫氰酸鹽的形成,在pH 6.5條件下,這種抑制效應明顯降低,最終在pH 7.5條件下抑制作用完全消失[69]。現已證實,TFP蛋白會影響硫苷代謝產物的結構,它會催化糖苷配基轉化為異硫氰酸酯或腈類,而二價鐵離子是TFP蛋白的輔酶因子[70]。此外,鐵離子也會影響ESP和NSP的活性,誘導形成腈類。鐵離子對硫苷代謝影響研究主要見于植物體系中硫苷酶解反應,目前在發酵蔬菜硫苷代謝中作用尚待證實。在制作發酵酸菜時加入亞硒酸鈉,結果表明硒元素主要以硒甲基硒代半胱氨酸形式存在于酸菜中,酸菜中抗壞血酸原略有下降,而吲哚-3-甲醇和吲哚-3-乙腈含量大幅增加,與對照組相比分別增加74%和13%[71],然而,硒對硫苷代謝的作用機制尚待進一步研究。
4.5 pH值
蔬菜發酵過程中,其中微生物存在動態變化過程。例如,酸菜發酵開始往往是以異型發酵乳酸菌占主導,主要是腸膜明串珠菌,隨著這些乳酸菌生長,產生大量乙酸和乳酸,發酵體系pH值隨之降低,當酸質量分數達到0.7%~1.0%、pH值低于4.5時,明串珠菌被酸耐受性更強的同型發酵乳酸菌所替代,主要是植物乳桿菌和短乳桿菌。同型發酵乳酸菌繼續產生乳酸,pH值進一步下降到3.4~3.7[72-73]。硫苷被降解時,首先與硫相連的糖苷被打開,生成葡萄糖與不穩定的糖苷配基,環境pH值與特殊蛋白共同作用對糖苷配基的下游產物形成具有重要的影響。一般來說,pH值接近中性時,糖苷配基容易發生結構重排;pH值較低時會抑制洛森重排。ESP存在會促進腈類形成,但是酸性(pH 4)或堿性(pH 8)下,ESP活性被抑制,反而有利用生成更高含量的異硫氰酸鹽[74]。然而,不同ESP亞型的最適pH值有所差異。Witzel等[75]從甘藍中鑒定出3 種不同亞型ESPs:BoESP1、BoESP2和BoESP3。BoESP1和BoESP2活性的最適pH值在6~7,而BoESP3在pH 4~7之間活性都保持穩定。
4.6 加熱處理
Sarvan等[55]研究發現在發酵之前沸水燙漂處理甘藍5 min,再接種副干酪乳桿發酵71 h,總硫苷含量為發酵前的35%,與傳統發酵方式相比,燙漂預處理起到了滅酶作用,使得發酵甘藍中保留了較多含量硫苷。西蘭花泥經過121 ℃高壓滅菌3 min,只檢測出屈曲花苷、2-羥基-3-丁烯基硫苷和蘿卜硫苷等幾種硫苷,且含量十分微少;而在西藍花中已被發現報道的5-甲基亞磺酰戊基硫苷、4-羥基-3-吲哚甲基硫苷、芝麻菜苷、蕓苔葡糖硫苷、4-甲氧基-3-吲哚甲基硫苷和新葡萄糖蕓苔素等硫苷在西蘭花泥中均未發現,可能是由于耐熱性差,硫苷經高壓處理已經被降解[54]。由此可見,加熱溫度會影響發酵蔬菜硫苷含量。ESP作為一種熱敏感蛋白,在室溫下會促進蘿卜硫苷更多形成蘿卜硫素腈,而不是蘿卜硫素。當新鮮西蘭花的芽或花在均質前于60 ℃下加熱,形成的西蘭花泥中蘿卜硫素含量增加,而蘿卜硫素腈降低,并且與ESP活性降低趨勢一致。在70 ℃及以上溫度下加熱,西蘭花中蘿卜硫素和蘿卜硫素腈含量均降低[76]。
5 結 語
硫苷及其代謝產物具有多種生理功能,因此受到人們廣泛關注。本文重點闡述了它們在抗氧化、抗癌、調節腸道菌群、緩解代謝綜合征等方面的最新研究進展。硫苷從惰性狀態轉化為具有生理功能的活性代謝產物,除了可以借助黑芥子酶的催化作用,微生物同樣也具有相似功能。在發酵十字花科蔬菜中,乳酸菌不僅可以代謝糖類,產生乳酸、乙酸等,也可以降解硫苷等植物化學物質,形成發酵蔬菜的特征性風味。乳酸菌對硫苷代謝作用具有菌株特異性,此外,與黑芥子酶作用類似,硫苷代謝還受到體系pH值、鹽含量、金屬離子等外在因素的影響。
然而,關于發酵蔬菜中硫苷代謝目前仍有許多問題亟待解決,主要體現在以下幾個方面:1)某些乳酸菌具有代謝硫苷的能力,但是相關的類黑芥子酶系仍未發現;2)發酵蔬菜中硫苷代謝往往是植物源黑芥子酶和乳酸菌共同作用的結果,兩者作用關系尚不明確;3)發酵蔬菜硫苷代謝受到鹽含量、金屬離子、pH值等因素影響,其作用機制也需要進一步明確;4)考慮到噁唑烷硫酮、腈類等硫苷代謝產物具有一定的毒性,發酵十字花科蔬菜硫苷的潛在安全問題需要得到重視。
