蠋蝽病毒Arma chinensis virus-1基因組克隆及序列分析
2022-01-08 09:26:00孫梅雪張長華賈芳曌徐蓬軍任廣偉
煙草科技 2021年12期
關鍵詞:分析
孫梅雪,張長華,高 強,李 煒,賈芳曌,徐蓬軍*,任廣偉*
1.中國農業科學院煙草研究所,山東省青島市科苑經四路11號 266101
2.貴州省煙草公司遵義市公司,貴州省遵義市匯川區人民路341號 563000
3.山東臨沂煙草有限公司沂水分公司,山東省沂水縣長安中路 276400
蠋蝽(Arma chinensis),又名蠋敵,隸屬半翅目(Hemiptera)蝽科(Pentatomidae)益蝽亞科(Asopinae)蠋蝽屬(Arma),其成蟲和若蟲均可捕食黏蟲、刺蛾、葉甲、蚧殼蟲和蚜蟲等40 余種常見害蟲[1]。蠋蝽分布較廣,國內主要分布于黑龍江、北京、云南、貴州等近20 個省份[2-4]。蠋蝽的食性廣、適應性強,可人工規模化飼養,是害蟲生物防治的一類重要天敵資源。
蠋蝽最初用于林業害蟲的防治,且防效較好[5-6]。近年來,貴州省煙草公司遵義市公司率先建立了蠋蝽繁育基地,構建了蠋蝽規模化繁育技術體系。該技術體系現已成為煙草行業綠色防控重大專項推薦的關鍵技術之一,并在全國主要煙區進行了示范推廣。目前,在蠋蝽的生物學特性[7]、捕食能力、人工飼料和規模化繁育技術等方面已有較為完善的研究。唐藝婷等[8]研究表明蠋蝽5 齡若蟲和雌蟲對斜紋夜蛾具有較好的捕食能力。人工飼料在蠋蝽規模化飼養過程中起著至關重要的作用,鄒德玉等[9-10]通過對飼喂人工飼料的蠋蝽和飼喂柞蠶蛹的蠋蝽分別進行轉錄組測序分析,發現了一些與營養相關的代謝途徑和差異表達基因,獲得了更多關于人工飼料配方的信息反饋。就飼養密度而言,雷庭等[11]研究表明室內飼養蠋蝽應嚴格控制飼養密度,密度過高會導致種群自殘,產卵前期推遲,產卵量降低。然而,蠋蝽規模化繁育時常出現蠋蝽因染病導致個體大量死亡的問題,嚴重威脅蠋蝽的規模化生產及推廣應用,但有關蠋蝽病原物方面的研究還鮮見報道。為此,本研究中采用高通量測序技術在蠋蝽體內鑒定出一種新病毒,將其命名為Arma chinensis virus-1(AcV-1),并分析了AcV-1 的基因組序列,旨在明確其基因組結構特征及遺傳進化情況,為蠋蝽病原物防控提供依據,從而提高蠋蝽成活率,為蠋蝽規模化繁育提供技術支撐。
1 材料與方法
1.1 材料
供試蠋蝽樣品于2019 年5 月采自貴州省煙草公司遵義市公司天敵繁育基地(27.45°N,106.53°E)。收集的蠋蝽樣本(200 頭)包括若蟲和成蟲(每個時期各100 頭),樣本保存于中國農業科學院煙草研究所。
1.2 方法
1.2.1 文庫的構建
每10 頭蠋蝽為一組,在液氮下混合研磨,每組取0.1 g 粉末置于1.5 mL 無菌無酶離心管中,加入1 mLReagent(美國Thermo Fisher 公司),剩余粉末儲存于-80 ℃冰箱備用。加入TRIzol?Reagent 的樣品4 ℃12 000 r/min 離心5 min,將每份樣品中的50 μL 上清液混合在一起提取總RNA。用Oligo(dT)(美國Thermo Fisher 公司)提取mRNA,然后在片段化緩沖液(美國Thermo Fisher 公司)中將mRNA 剪切成短片段。隨后,使用TruSeq RNA Sample Prep Kit(美國Thermo Fisher 公司),參照其說明書,用合適的插入片段[約(200±50)bp]構建文庫。使用Illumina HiSeq?儀器(美國Illumina 公司)對樣品進行RNA-seq 分析[12]。
1.2.2 組裝和注釋
使用Illumina HiSeqTM儀器進行測序時,選擇成對末端和讀取長度為150 nt 的選項,而不選用鏈特異性選項。使用Trinity 2.0.6 修剪接頭,舍棄低質量讀取并使用清晰高質量讀取從頭組裝[13],使用BLASTx(E-value ≤1×10-5)將組裝的重疊群與非冗余蛋白質數據庫、STRING、Swissprot 和KEGG 比對,進行功能注釋。
1.2.3 病毒檢測、基因組擴增
(1)RNA 提取和反轉錄
取儲存于-80 ℃冰箱的蠋蝽粉末樣品0.1 g 于1.5 mL 無菌無酶離心管中,加入TRIzol?Reagent混勻,用于提取總RNA,具體操作參照說明書進行。提取的RNA 用50~100 μL DEPC 水溶解,檢測RNA 濃度和純度后,置于-80 ℃冰箱中保存。
(2)AcV-1 基因組擴增
根據RNA-Seq 得到的AcV-1 基因組已知序列,設計7 對特異性引物(表1),以反轉錄產物cDNA 為模板進行PCR 擴增,各片段擴增體系:TaKaRa LA Taq 0.5 μL、10×LA Taq Buffer II(Mg2+Plus)5 μL、dNTP Mixture(2.5 mM each)8 μL、上下游引物各1 μL、cDNA 2 μL、ddH2O 32.5 μL,總體積50 μL。擴增條件:94 ℃5 min;94 ℃30 s,55 ℃30 s,72 ℃1.5 min,循環35 次;72 ℃7 min。AcV-5R 和AcV-3F1/AcV-3F2分別用于5′和3′cDNA 末端快速擴增(Rapid amplification of cDNA ends,RACE),使用SMARTer?RACE 5′/3′Kit(美國TaKaRa 公司),按照產品說明進行實驗。上述PCR 產物于1.0%瓊脂糖凝膠中電泳檢測,目的片段參照EasyPure?Quick Gel Extraction Kit[全式金(北京)生物技術有限公司]說明進行切膠回收。回收目的片段與載體pEASY-T1 25 ℃連接30 min,熱激法轉化大腸桿菌DH5α感受態細胞,涂布于含氨芐青霉素(Ampicillin,Amp)的培養基過夜培養,挑取平板上的克隆進行菌落PCR,將PCR 結果中大小正確條帶對應的菌液樣品送上海派森諾生物科技股份有限公司進行序列測定。

表1 AcV-1 基因組擴增引物Tab.1 Amplificɑtion primers used for AcV-1 genome
1.2.4 基因組結構分析及系統發育分析
序列測定完成后,根據擴增引物去除兩端的載體序列,通過拼接獲得AcV-1 的基因組序列。使用ORF Finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)搜索病毒基因組的ORF,用Clustal W軟件來比對核苷酸序列和氨基酸序列,用NCBI 保守域搜索預測保守域。使用BLAST(http://www.ncbi.nlm.nih.gov/BLAST/)進行序列多重比對及同源性分析。將獲得的新病毒基因組序列與雙順反子病毒科(Dicistroviridae)目前接受的15 種雙順反子病毒的相應氨基酸序列進行比對,用MAGE-X軟件進行聚類分析并構建系統發育樹[14],采用鄰接 法(Neighbor-joining,NJ)分別構建ORF1 和ORF2 預測氨基酸的系統發育樹,距離模型選擇差異位點比例(p-distance),并使用自舉法(Bootstrap)1 000 次進行穩定性檢驗;采用最大似然法(Maximum likelihood,ML),以隸屬伴生豇豆病毒科(Secoviridae)的水稻東格魯球狀病毒(Rice tungro spherical virus)、歐防風黃點病毒(Parsnip yellow fleck virus)和隸屬傳染性軟腐病病毒科(Iflaviridae)的褐飛虱蜜露病毒-2(Nilaparvata lugens honeydew virus-2)為外群(Outgroup),構建保守結構域RdRp 的系統發育樹,其可信度使用1 000 次自舉重復驗證。
2 結果及分析
2.1 蠋蝽中AcV-1 高通量測序
RNA-Seq 數據提交到NCBI Sequence Read Archive(SRA)數據庫(登錄號:SRR10098905)。通過RNA-seq獲得了60億個堿基,總共產生了27 058個重疊群,其中包括21 972 個單基因。通過功能注釋的方法,發現了一個組裝的重疊群(長度為9 176 nt),通過Blast 分析,發現其與蟋蟀麻痹病毒(Cricket paralysis virus,CrPV)分離物CrPV-3(GenBank:KP974707.1)全基因組的核苷酸序列相似性達89.4%,將該重疊群命名為AcV-contig1。
2.2 AcV-1 基因組序列分析
成功擴增了AcV-1 基因組的3′端和5′端(圖1a、圖1b),拼接后得到的AcV-1 基因組序列長度為9 192 nt[不包含Poly(A)],堿基組成分別為33.3% A、27.6% T、18.2% C、20.9% G(圖1c)。該序列已上傳至GenBank(登錄號為MW846634)。拼接的AcV-1 病毒基因組序列與RNA-seq 獲得的序列相比無堿基變異,但在5′端多出25 個堿基,另外,可能由于RNA-seq 測序拼接的問題,3′端存在2 個堿基的缺失。基因組序列與CrPV(GenBank:KP974707.1)的結構和長度相似,其中核苷酸相似性最高達89.40%,表明本研究中獲得了含有完整讀碼框的AcV-1 基因組序列。該病毒基因組包含2 個非重疊的ORFs,ORF1 含有5 316 個核苷酸,該ORF 起始于717 nt,終止于6 032 nt,分子質量為203.68 kDa,pI 為6.67,編碼一個長度為1 771 個氨基酸的非結構蛋白,包含3 個保守結構域,分別是依賴RNA 的RNA 聚合酶(RNA-dependent RNA polymerase,RdRp)、蛋白酶(Protease,P)、RNA 解旋酶(RNA helicase,HEL)。對ORF1 推導出氨基酸序列進行BLASTp 分析,結果表明其與雙順反子病毒(Dicistroviruses)和微小核糖核酸病毒(Picorna-like viruses)的非結構蛋白關系較近,其中與CrPV 的非結構蛋白(UniProtKB/Swiss-Prot:AKA63263.1,相似性97.74%,覆蓋率100%)的相似性最高。ORF2含有2 529個核苷酸,該ORF起始于6 384 nt,終止于8 912 nt,分子質量為94.3 kDa,pI 為7.2,編碼一個包含842 個氨基酸的結構蛋白,包括蟋蟀麻痹病毒樣衣殼蛋白、蟋蟀麻痹病毒VP4 衣殼蛋白以及兩個小核糖核酸病毒樣衣殼蛋白。2 個ORFs 中間由一個長度為351 nt 的基因間隔區(Intergenic region,IGR)隔開,其兩端為非翻譯區(Untranslated regions,UTRs),5′和3′非翻譯區分別含有716個和280個核苷酸,且3′端具有Poly(A)尾。BLASTp 分析表明,ORF2 同樣也是與雙順反子病毒和微小核糖核酸病毒關系較近,其中與CrPV 的結構蛋白(UniProtKB/Swiss-Prot:AKA63264.1,相似性93.70%,覆蓋率99%)的相似性最高。
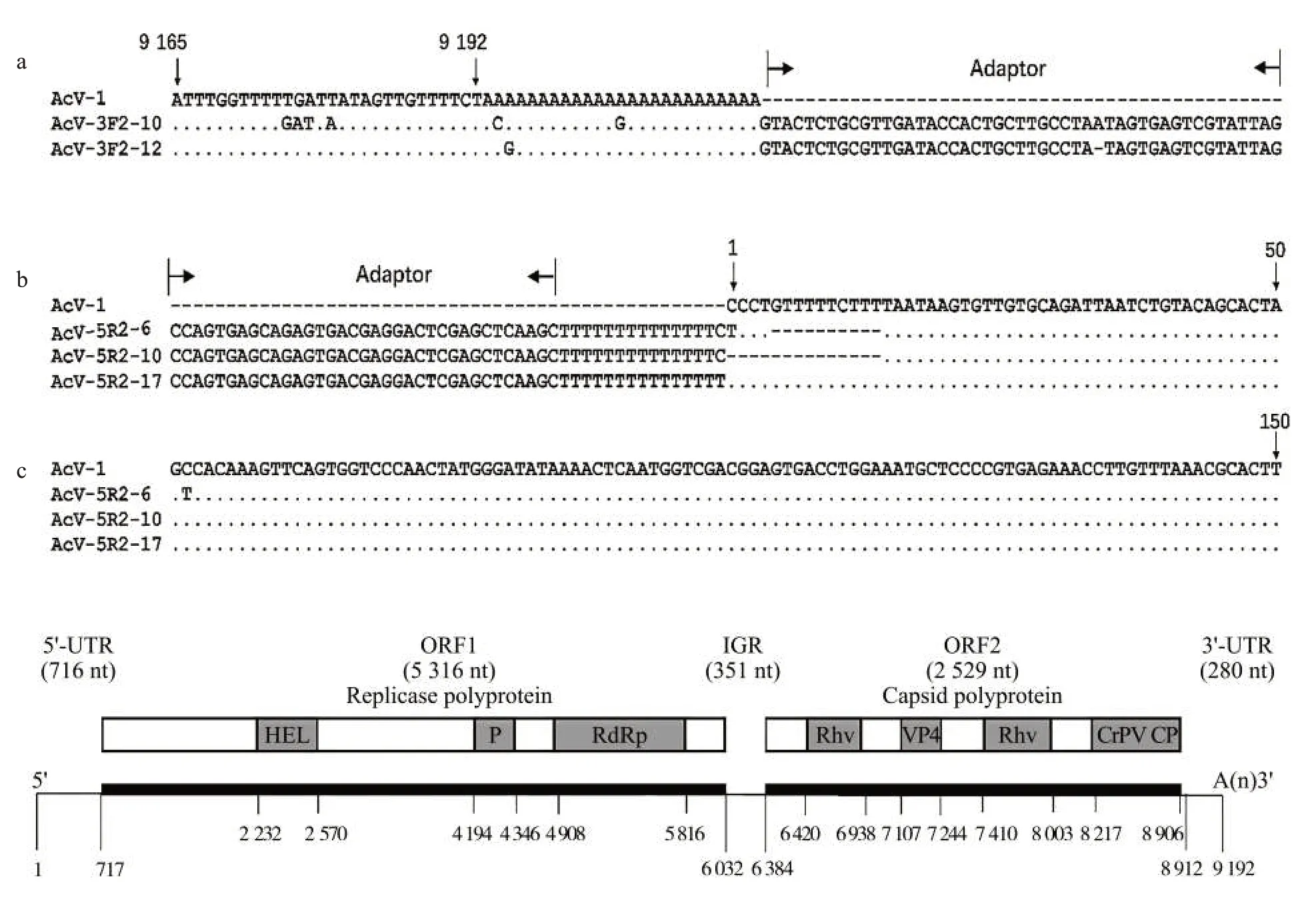
圖1 AcV-1 基因組3′端/5′端擴增及基因組結構分析Fig.1 Amplification of the 3′/5′ends and structural analysis of AcV-1 genome
2.3 AcV-1 系統發育分析
通過Blastp 對推導的ORF1 和ORF2 氨基酸序列進行同源性分析,結果顯示其與雙順反子病毒科中的CrPV 具有高度相似性。采用系統發育分析方法對推導的ORF1、ORF2 以及RdRp 的氨基酸序列進行了系統發育分析,進一步確定AcV-1 和雙順反子病毒科之間的關系。
采用最大似然法構建保守結構域RdRp 的系統發育樹時,應首先對模型進行選擇,經過篩選,此次分析選擇貝葉斯信息標準得分最低的LG+伽馬分布(Gamma distributed,G)模型。基于RdRp的ML 樹表明,AcV-1 與褐飛虱C 病毒(Nilaparvata lugens C virus,NLCV)聚集在一起,但自舉值(54,圖2)較低,而基于ORF1 和ORF2 的氨基酸序列的NJ 樹表明,AcV-1 與CrPV 聚集在一起且具有較高自舉值(分別為100 和81,圖3a、圖3b)。
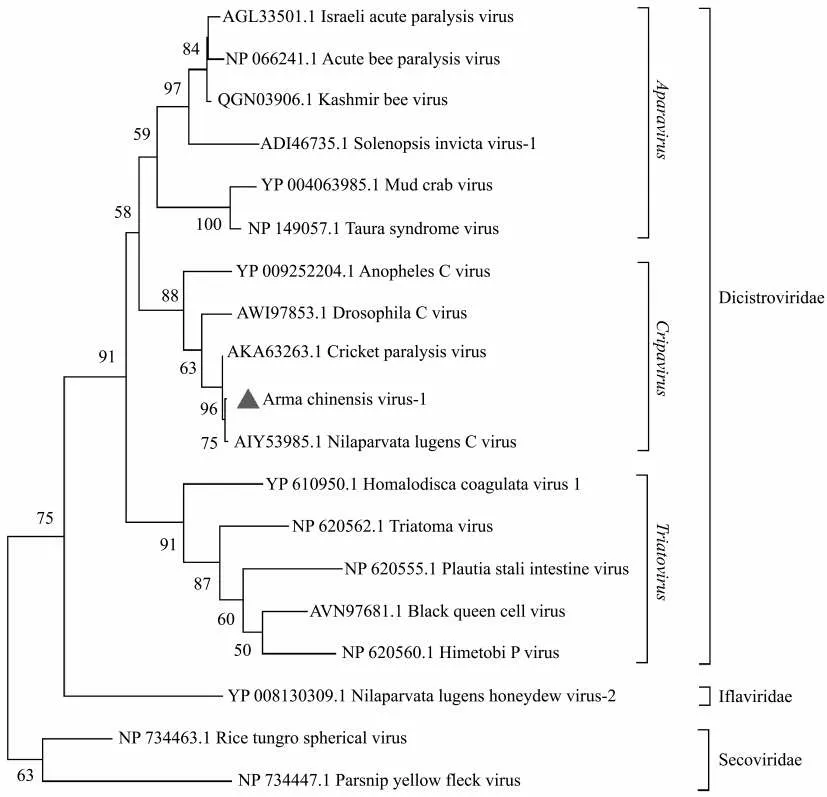
圖2 基于RdRp 的ML 樹Fig.2 Maximum-likelihood(ML)tree based on RdRp

圖3 基于ORF1/ORF2 的NJ 樹Fig.3 Neighbor-joining(NJ)tree based on ORF1/ORF2
2.4 不同來源蠋蝽感染AcV-1 情況檢測
利用特異性引物AcV-1-F/AcV-1-R(AcV-1-F:CACAGGGGATTTTACTGAAGG,AcV-1-k:CGTC GGGTGTACCTGTAGAAA;擴增片段長度為725 bp)通過RT-PCR 方法對供試蠋蝽樣品中AcV-1 侵染情況進行檢測。結果表明,146 份樣品中,55 份樣品呈AcV-1 陽性,即感染率為37.67%(表2)。

表2 不同來源蠋蝽感染AcV-1 情況檢測Tab.2 Infection rates of AcV-1 in Arma chinensis from different sources
3 討論
本研究中利用RNA-seq 測序技術在蠋蝽體內發現的AcV-1 是一種新雙順反子病毒。雙順反子病毒是一類小型無包膜RNA 病毒,此類病毒含有一個長度約8~10 kb 的正義單鏈RNA 基因組[15]。雙順反子病毒科分為3 個屬:蜜蜂急性麻痹病毒屬(Aparavirus),蟋蟀麻痹病毒屬(Cripavirus)和吸血獵蝽病毒屬(Triatovirus),該分類基于IGR-IRES表現出的系統發育差異和獨特特征[16-17],通過病毒蛋白衣殼和RdRp 的系統發育分析也支持了這種分類[18]。蠋蝽病毒AcV-1 基于ORF1 和ORF2 氨基酸序列的NJ 樹表明,該病毒與CrPV 聚在一起,而基于RdRp 氨基酸序列的ML 樹表明,其與NLCV 聚在一起。上述聚類分析結果表明AcV-1與CrPV、NLCV 具有很高的同源性,但又不盡相同,推測其為蟋蟀麻痹病毒屬的一個新成員。
利用檢測引物AcV-1-F/AcV-1-R 檢測146 份蠋蝽樣本,其中有55 份樣本呈陽性,即感染率為37.67%。昆蟲與病毒之間存在多種互作關系,可以將這些互作關系分為3 大類:致病、共生和潛伏侵染[19-21],科學利用寄主與病毒之間的關系,有益于農作物病蟲害綠色防控技術的發展。目前AcV-1 的功能尚不清楚,通過帶毒蠋蝽成活率及系統發育分析結果,推測該病毒可能無致死功能,但其對蠋蝽生長發育和生殖是否有顯著影響及其與蠋蝽的互作機制尚待進一步研究。若為有害病毒,可根據其傳播途徑等構建防控技術體系;若為有益病毒,可進一步加以利用,為蠋蝽商業化應用奠定基礎。
4 結論
本研究中在蠋蝽體內發現了一種新的雙順反子病毒,并將其命名為Arma chinensis virus-1(AcV-1)。該病毒含有完整讀碼框的基因組序列,長度為9 192 nt(不包含Poly A),包含2 個ORFs。ORF1 編碼非結構蛋白,包含3 個保守結構域,分別是RdRp、蛋白酶和RNA 解旋酶;ORF2 主要編碼衣殼蛋白。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
