基于流刺網調查的海陽近岸口蝦蛄漁業生物學研究*
2022-01-14 07:48:16鑫王蕾邱盛堯劉淑德董秀強
漁業科學進展 2022年1期
徐 鑫王 蕾邱盛堯① 劉淑德董秀強
(1.煙臺大學海洋學院 山東 煙臺 264005;2.山東省水生生物資源養護管理中心 山東 煙臺 264005)
口蝦蛄(Oratosquillaoratoria),俗稱皮皮蝦,螳螂蝦等,常居于海底泥沙礫的U形洞穴中(王春琳等,1996),在我國沿海各地均有分布(徐海龍等, 2010),為我國沿海重要的經濟蝦蟹類(劉海映等,2013)。近年來,口蝦蛄已成為山東海陽近岸流刺網漁業的主要漁獲物(李春芝等,2019),經濟價值的提高導致對其捕撈強度不斷增強(劉修澤等,2014),其種群面臨前所未有的生存壓力。為保護海陽近岸口蝦蛄資源,迫切需要了解其群體現狀及生活史特征。
20世紀80年代,日本的口蝦蛄漁業因過度捕撈造成產量急劇下降(Ohtomi et al, 2005),資源的劇烈波動引起政府和學者的關注。生物學家通過生態控制實驗和野外調查,對分布于日本周邊幾個海灣內的口蝦蛄的生長、繁殖和種群動態等進行研究發現,雖然生活于不同海域的口蝦蛄具有相似的生活史過程,但在生長規律(Hamano et al, 1987; Kodama et al, 2010)、繁殖策略(Kodama et al,2006、2003;Ohtomiet al,1988)、生命周期和攝食組成(Hamano et al, 1992)等方面存在差異。國內學者對口蝦蛄的研究主要集中在漁業生物學(鄧景耀等, 1992;盛福利, 2009)、繁殖與攝食(劉海映等, 2013;王春琳等, 2002;徐善良等, 1996;薛梅等, 2016)、形態性狀(張新明等, 2020)及資源分布(劉修澤等, 2014;李春芝等, 2019)等方面。李明坤等(2019)分析了山東南部近海口蝦蛄的空間分布特征及季節變化,孫東昱等(2021)研究了海陽與煙臺近岸口蝦蛄的形態比較,但還未見關于海陽近岸口蝦蛄群體狀況的研究報道。本研究通過對海陽近岸口蝦蛄的月度跟蹤調查,分析海陽近岸口蝦蛄的群體結構組成、生長與繁殖等漁業生物學特征,掌握其群體動態變化規律,旨在為山東南部海域口蝦蛄資源的科學管理和合理的開發利用提供依據。
1 材料與方法
1.1 調查方法和樣品采集
本研究所用口蝦蛄樣品為2018年3月—2019年9月在山東半島南部海陽近海的流刺網漁獲(2018年6—8月及2019年2月、5—8月除外),每月隨機取5 kg樣品,共計1538尾。生物學測量1514尾,其中,雌性744尾,雄性770尾(表1)。流刺網網具為口蝦蛄漂流三重刺網,上綱長度為27.00 m,側綱為3.50 m,網目為60.00 mm。
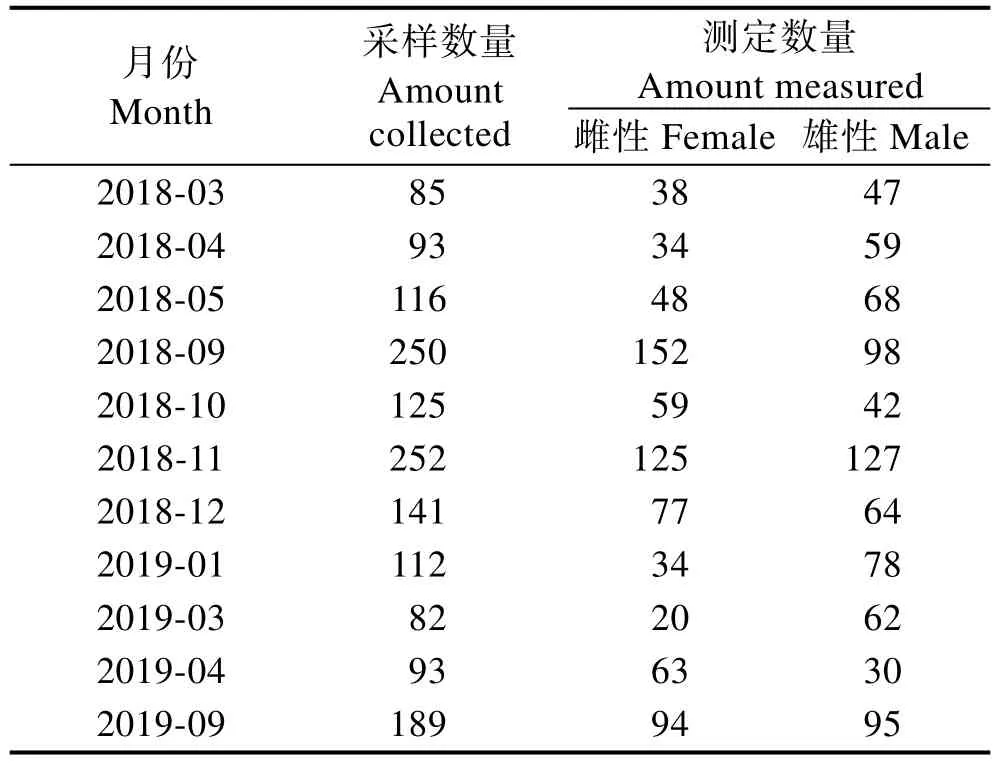
表1 海陽近岸流刺網口蝦蛄各月樣品采集數量Tab.1Sample quantity of O. oratoria collected from Haiyang offshore gillnet in each month
1.2 材料處理與數據分析
1.2.1 體長及體質量 口蝦蛄樣品根據有無交接肢區分雌、雄(徐善良等,1996),利用游標卡尺測量其體長(L),精度為0.01 mm;利用分析天平測量其體質量(W),精度為0.01 g。
1.2.2 性比及性腺發育 對口蝦蛄進行性別鑒定,解剖雌性個體,口蝦蛄卵巢發育階段劃分參考王春琳等(1996)的方法,大致可分為5期:
Ⅰ期(未發育期):卵巢呈細線狀分布,無色素分布,緊貼在消化道上,寬度為1~2 mm。
Ⅱ期(發育期):卵巢開始發育,向兩側擴大,呈薄帶狀,半透明,淺黃色,開始有少量色素分布。
Ⅱ'期(恢復期)為排卵后不久的個體擁有:卵巢灰黃色,有較多色素沉積,大小與Ⅱ期卵巢一致。
Ⅲ期(成熟前期):卵巢迅速生長,縱向已發展到尾節。輕壓卵巢有彈性,解剖鏡下能見到清晰的卵粒,不能分離。
Ⅳ期(成熟期):卵巢已進入成熟期,幾乎充滿整個腹腔背部。卵巢呈橙黃色,卵粒已能分離,為不規則圓形。為即將產卵的個體擁有。
Ⅴ期(產卵期):卵極度膨大,通過甲殼隱約能見到橙黃色卵巢,解剖卵巢,卵子能自動散開。
卵子產出后,抱卵于胸部附肢上,同時,排空的卵巢開始萎縮吸收,逐漸進入Ⅱ′期(恢復期),待下次繁殖季節再次發育。
1.2.3 攝食強度 參考徐善良等(1996)劃分的口蝦蛄攝食強度等級觀察胃囊的充實程度,分為4級:0級為空胃,1級為胃中食物含量較少,2級為胃含物較多但不飽滿,3級為胃含物很多、飽滿且胃壁膨脹。
1.2.4 體長–體質量關系 對體長–體質量關系運用冪函數回歸方法進行分析。體長和體質量呈冪函數增長關系,其關系式為:

式中,W為體質量(g),L為體長(mm),a、b為系數。
1.2.5 體長相對生長速率 本研究參考甲殼動物群體中區分不同世代的體長頻率分布法(Peterson,1892),以每月流刺網漁獲樣本中優勢體長組的中值為參考點,選取體長組中值±5 mm的口蝦蛄樣本,作為體長相對增長率樣本的計算對象,計算月間體長相對增長速率,計算公式(Changet al, 2012)如下:
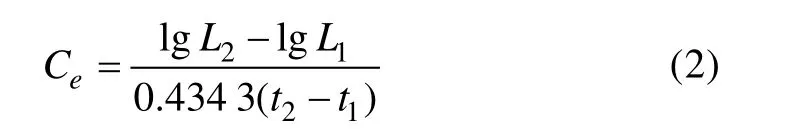
式中,0.434 3為自然對數轉換為以10為底數一般對數的系數;L1和L2為計算生長速率的前一月和當月的口蝦蛄體長;t1和t2為以生長開始起,即需要計算生長比速的前一月和當月。
1.2.6 統計檢驗 使用t檢驗分析比較雌、雄月度平均體長及體質量間的差異,使用K-S檢驗分析比較雌、雄月度優勢體長組及優勢體質量組的分布差異。對口蝦蛄雌、雄比例,以1∶1為期望值進行卡方檢驗(x2=0.50,df=2)。
2 結果
2.1 體長組成
2018年3月—2019年9月,流刺網口蝦蛄樣品體長范圍為62.00~180.40mm。雌性平均體長為(122.01±13.90)mm,2018年3月平均體長最大,為(131.49±8.68)mm;9月平均體長最小,為(106.49±8.62) mm。雄性平均體長為(129.27±13.99) mm,2019年1月平均體長最大,為(139.38±8.02)mm。2018年9月平均體長最小,為(117.04±9.93) mm。除2019年4月外,其余月份雄性平均體長均大于雌性(圖 2)。t檢驗結果顯示,2018年 4、12月雌、雄樣品平均體長差異不顯著(P>0.05),其余月份均存在顯著差異(P<0.05)。
雌性口蝦蛄全年優勢體長為100~140 mm,占總尾數的84.96%。優勢體長組月間變化明顯,其中,2018年3、12月及2019年1、4月優勢體長組為120~140 mm;2018年4、5月及2019年3月優勢體長組為120~150 mm;2018年9月、2019年9月優勢體長組分別為100~120 mm及100~130 mm;2018年10月與11月優勢體長組為110~140 mm(圖1)。
雄性口蝦蛄的全年優勢體長為110~150 mm,占總尾數的 88.99%。優勢體長組的月間變化較明顯,其中,2018年4、5、10、11月及2019年3月的優勢體長組為120~150 mm;2018年3月及2019年1月的優勢體長組為130~150mm;2018年9月的優勢體長組為100~130 mm,12月的優勢體長為120~140mm;2019年 9月及 4月的優勢體長組均為110~140mm(圖1)。K-S檢驗表明,除2018年12月外,其余月份雌、雄樣品體長分布均存在顯著性差異(P<0.05)。
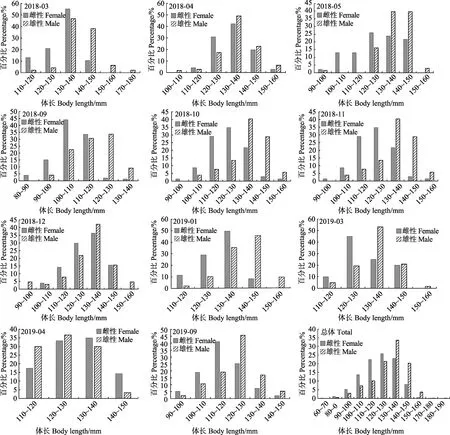
圖1 海陽近岸口蝦蛄體長分布的月間變化Fig.1Monthly variation in body length distribution of O. oratoria in the inshore waters of Haiyang
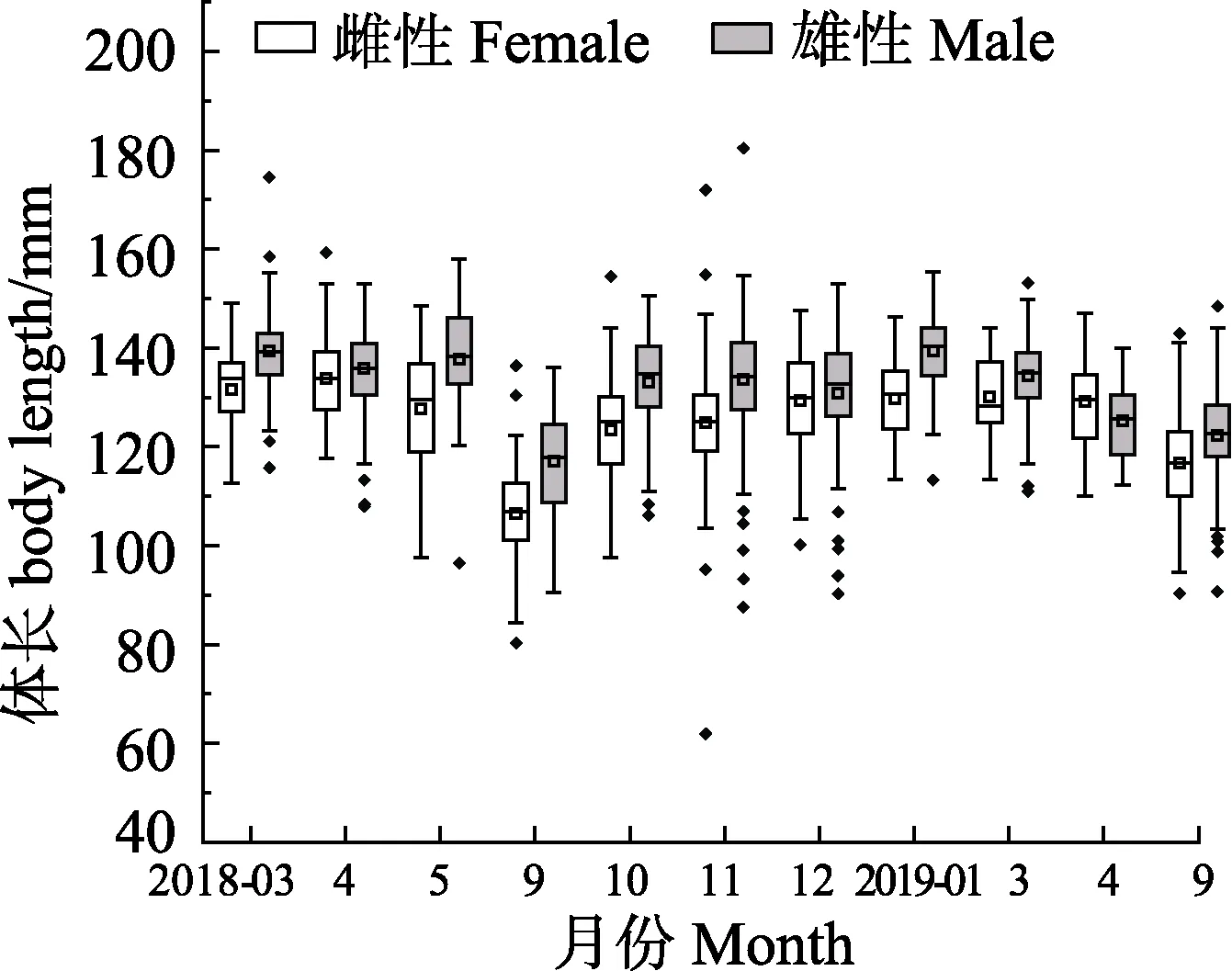
圖2 海陽近岸口蝦蛄雌性與雄性體長月間分布箱線圖Fig.2Boxplot of body length of female and male O. oratoria in the inshore waters of Haiyang
2.2 體質量組成
2018年3月—2019年9月口蝦蛄流刺網全年體質量范圍為8.30~64.20 g,雌性全年平均體質量為(25.79±8.87) g,2018年3月平均體質量最大,為(35.96±6.76) g;2018年9月平均體質量最小,為(16.44±3.65) g。雄性全年平均體質量為(32.49±10.46) g;2018年1月平均體質量最大,為(41.13±7.14)g,2018年 9月平均體質量最小,為(22.99±5.79) g。各月雌性平均體質量均小于雄性(圖3)。t檢驗結果表明,除2019年4月雌、雄樣品平均體質量差異不顯著(P>0.05)外,其余月份均存在顯著性差異(P<0.05)。
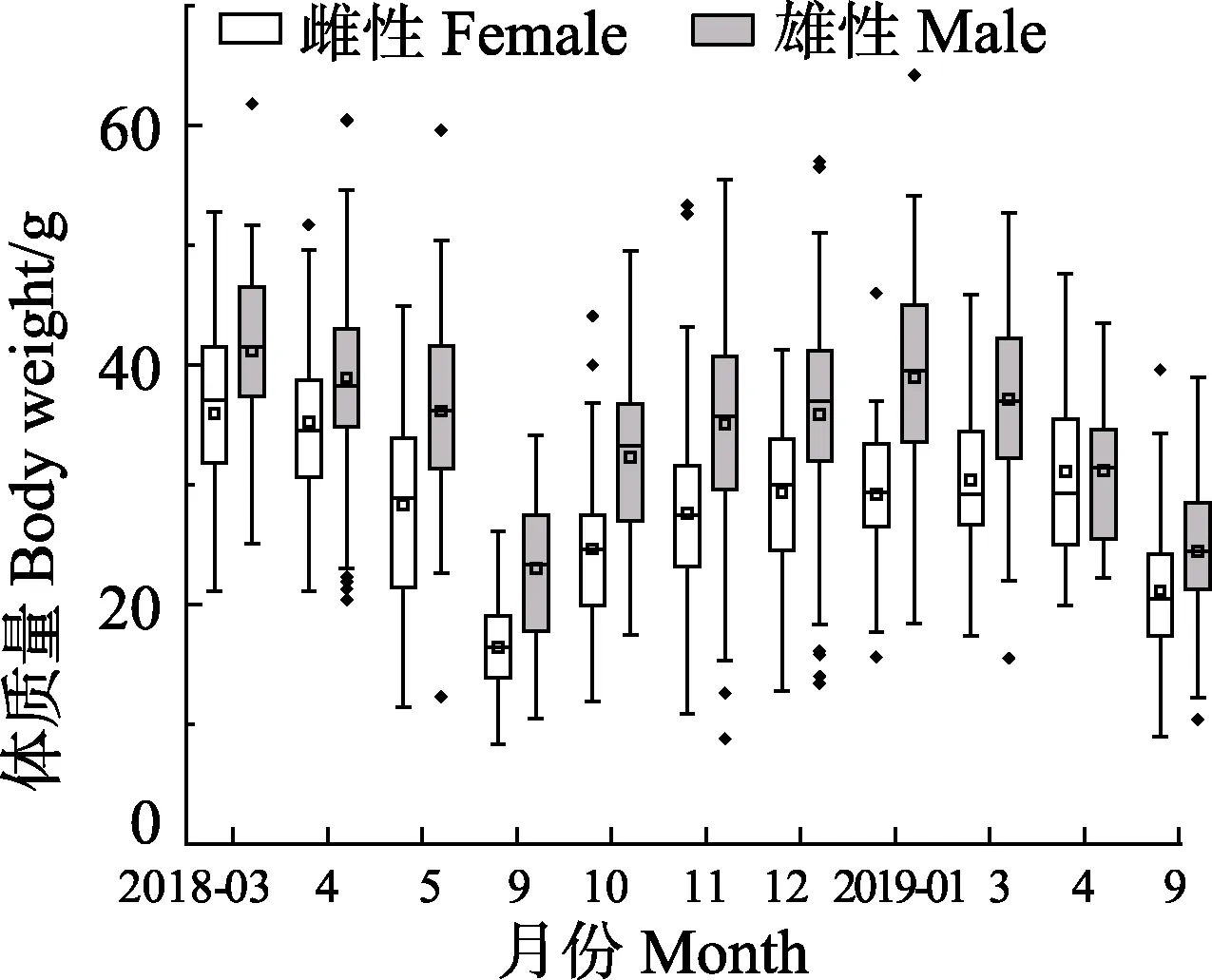
圖3 海陽近岸雌性與雄性口蝦蛄體質量月間分布箱線圖Fig.3Boxplot of body weight of female and male O. oratoria in the inshore waters of Haiyang
雌性口蝦蛄全年優勢體質量為10~40 g,占總尾數的91.38%。優勢體質量組月間變化明顯,其中,2018年3月優勢體質量組為20~50 g;2018年4、5月優勢體質量組分別為30~40 g、20~40 g;2018年9月及次年9月優勢體質量均為10~20 g;10月的優勢體質量組為10~40 g;2018年11月—次年4月的優勢體質量組均為20~40 g。雄性口蝦蛄全年優勢體質量為20~50 g,占總尾數的82.97%。優勢體質量組月間變化較明顯,其中,2018年3月及4月的優勢體質量組為30~50 g;2018年5月及11月的優勢體質量組為20~50g;2018年9月的優勢體質量組分別為10~30 g,10月及次年4月優勢體質量均為20~40 g;12月—次年3月優勢體質量組均為30~50 g;2019年9月優勢體質量組為20~30 g。K-S檢驗表明,除2019年4月外,其余月份雌、雄樣品體質量分布均存在顯著性差異(P<0.05)。
2.3 性比及性腺發育
本研究共測量1514尾流刺網口蝦蛄樣品,其中,雌性個體744尾,雄性個體770尾,雌、雄比為1∶1.03。卡方檢驗表明,雌、雄個體比例符合1∶1(P=0.094>0.05)。口蝦蛄各月份性比變化幅度明顯,2018年 9、10、12月及 2019年4、9月性比均>1,其余月份均<1;其中,2019年 4月的性比最大,為2.1∶1;3月性比最小,為 0.32∶1(圖 4)。口蝦蛄性比月間變化顯著(P<0.05),2018年9月及次年1、3、4月性比不符合1∶1,其他月份均符合1∶1(P>0.05)。
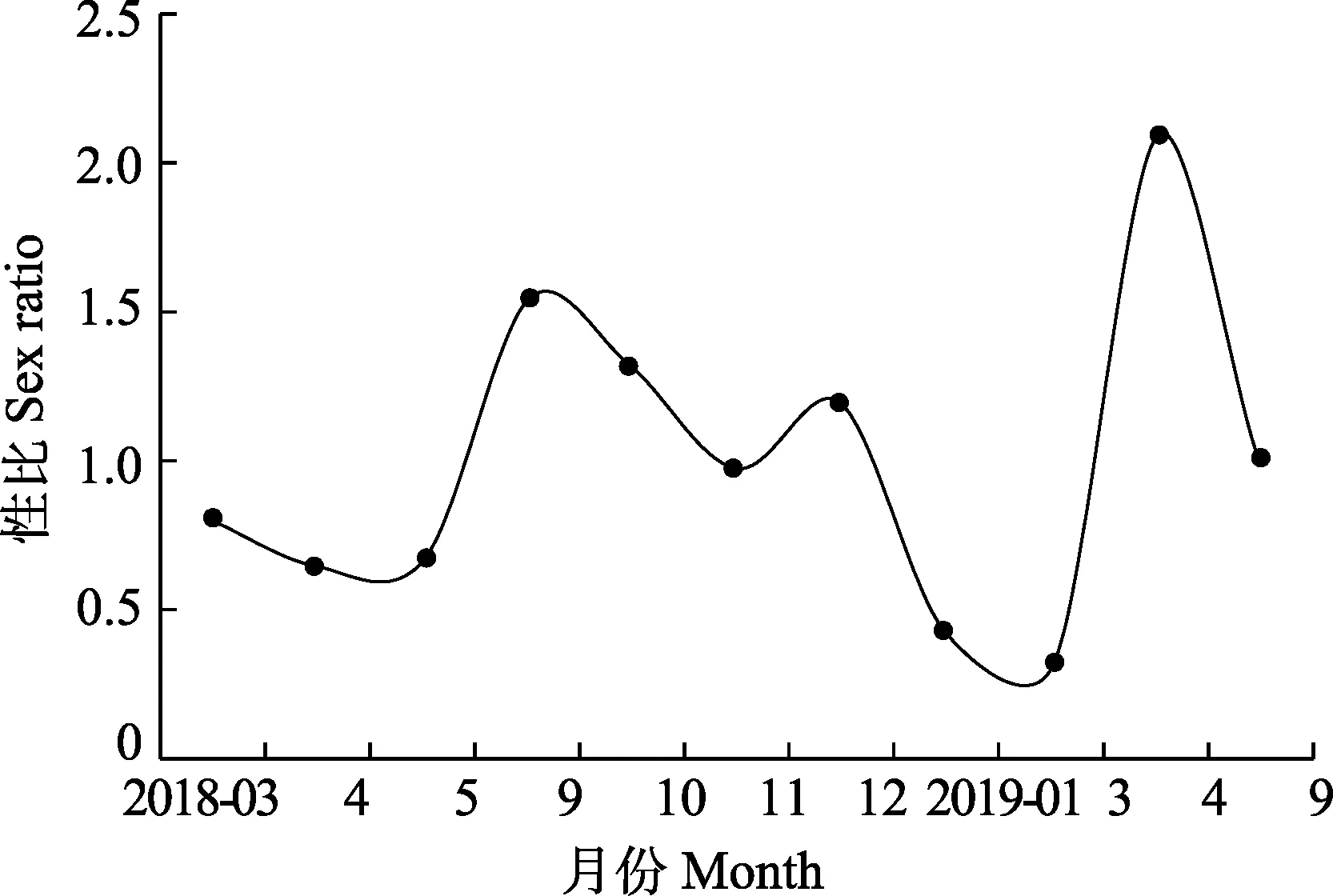
圖4 海陽近岸口蝦蛄性比月變化Fig.4Monthly variation of sex ratio of O. oratoria in the inshore waters of Haiyang
雌性口蝦蛄全年樣品生殖腺成熟度在Ⅰ~Ⅳ期,因為Ⅴ期個體處于排卵期,居于穴中進行產卵活動,流刺網很難捕捉,故未發現Ⅴ期個體。全年中,Ⅲ期和Ⅳ期個體主要出現在2個時間段,分別為4—5月和10月—次年1月(圖5)。
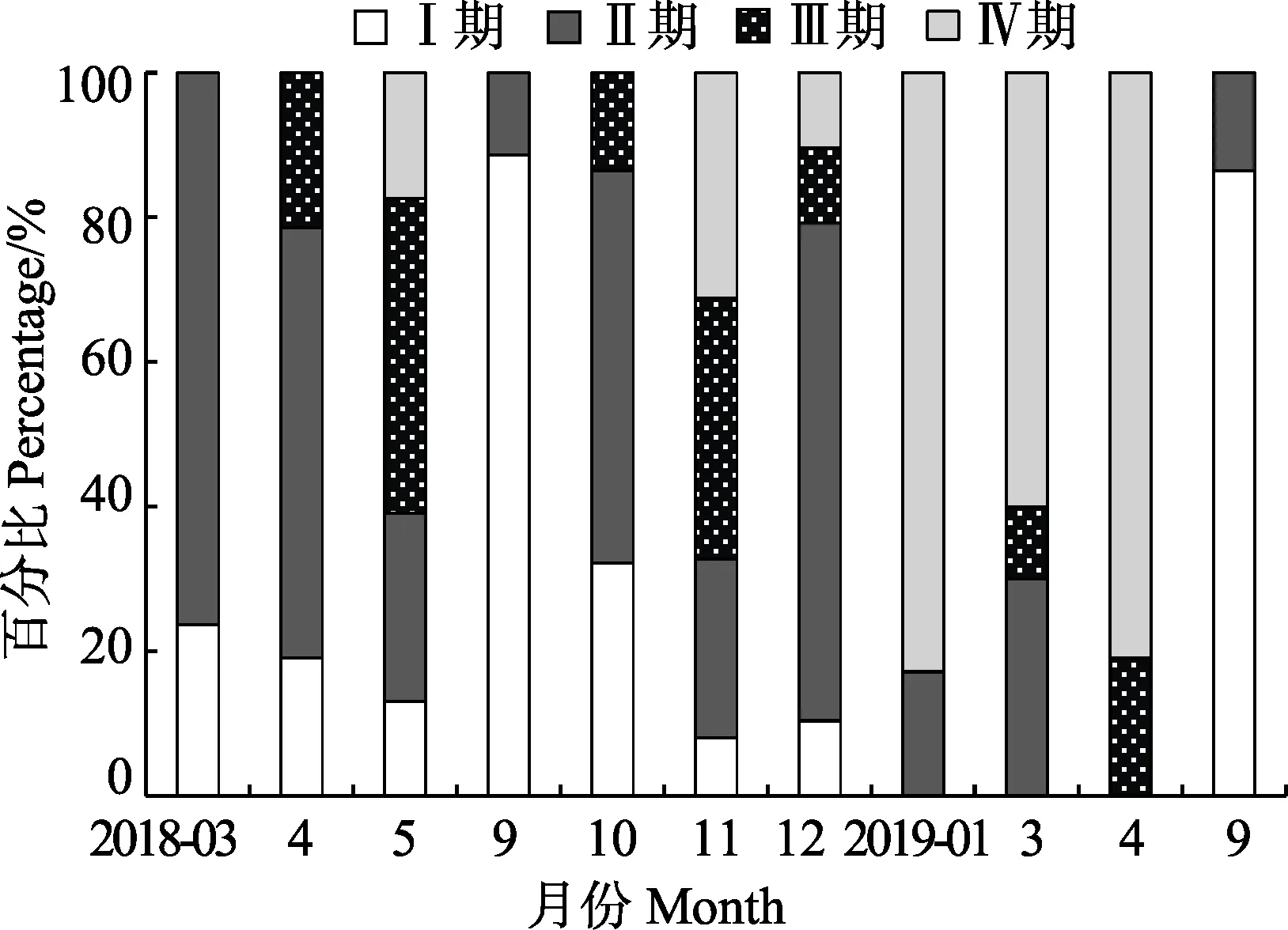
圖5 海陽近岸雌性口蝦蛄性腺發育程度月變化Fig.5Monthly variation of gonadal development of O. oratoria in the inshore waters of Haiyang
第1個時間段從4月開始,雌性口蝦蛄開始出現Ⅲ期的個體,占雌性總體的比例為21.43%,5月Ⅲ期占雌性總體的比例增加到43.48%,且出現17.39%的Ⅳ期個體,9月初未發現Ⅲ期和Ⅳ期個體,Ⅰ期個體占比高達86.67%。
第2個時間段從10月開始,同時出現Ⅲ期和Ⅳ期個體,分別占雌性總比例的47.89%和16.90%,11月Ⅲ期個體減少到36.00%,Ⅳ期個體增加至31.20%,12月Ⅲ期與Ⅳ期個體均減小至10.39%,到次年1月Ⅲ期的個體全部消失,Ⅳ期個體達到最高值82.86%。次年3月與4月均出現Ⅲ期與Ⅳ期個體,其中,3月Ⅳ期個體占60.00%,4月Ⅳ期個體占比高達80.95%。
2.4 攝食強度
口蝦蛄的攝食強度在全年的波動較明顯。雌性口蝦蛄3—5月攝食強度逐漸減小,其中,攝食強度達1級的個體所占比例由34.21%降低至23.91%,2級所占比例由 34.21%降低至 21.74%,3級由 5.26%增至13.04%,整體攝食強度減至58.70%。9—10月攝食強度達3級個體數量由4.61%增加至20.29%,10月空胃率全年最低,為10.14%;11月—次年4月攝食強度呈先上升后下降的趨勢,空胃率均大于50%,且未發現攝食強度達3級的個體,4月空胃率達到全年最高(96.83%),且僅存在攝食強度為1級的個體。9月空胃率明顯下降,為50.53%(圖6)。
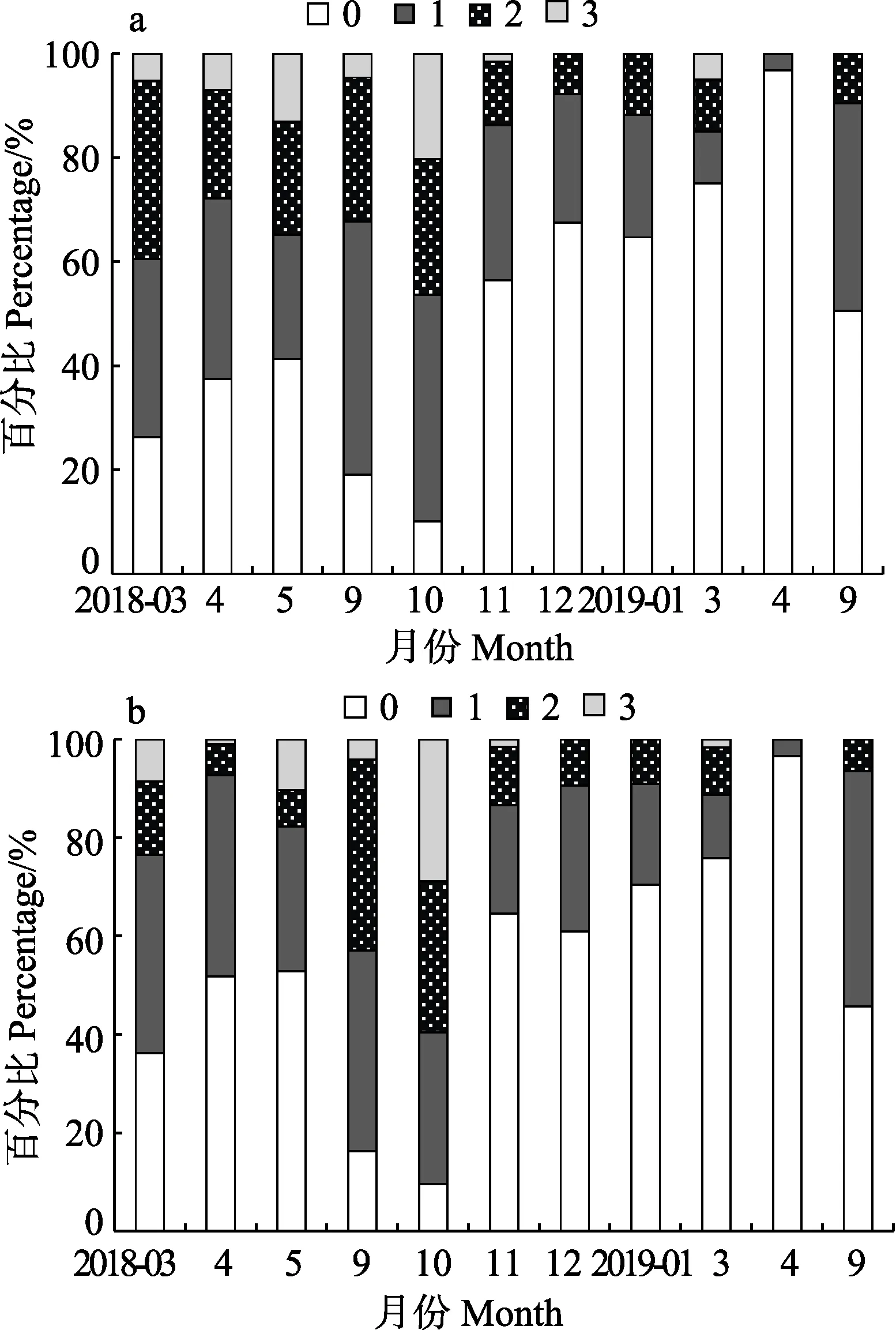
圖6 海陽近岸雌性(a)與雄性(b)口蝦蛄攝食強度的月變化Fig.6Monthly variation of feeding intensity of female (a)and male (b) O. oratoria in the inshore waters of Haiyang
雄性口蝦蛄與雌性個體攝食強度變化較為一致。雄性個體在3—5月攝食強度減小,攝食強度達1級和2級的個體數量分別減小至29.41%和7.35%,攝食強度達3級的個體數量由8.51%增加至10.29%,空胃率增加。9—10月空胃率逐漸下降,攝食強度達3級個體數量在10月占比最高,為28.85%,空胃率最低為9.62%;11月—次年4月,攝食強度呈逐漸下降的趨勢,4月最低,僅為3.33%,空胃率高達到96.83%;9月攝食強度明顯增加,但未發現達到3級的個體。
2.5 體長與體質量的關系
雌、雄口蝦蛄的體長與體質量均符合異速生長特征,其中,雌性口蝦蛄回歸方程為W=5.39×10–5L2.7098(R2=0.825 4, P<0.05),雄性口蝦蛄為 W=2.62×10–5L2.8744(R2=0.798 6, P<0.05)(圖 7)。
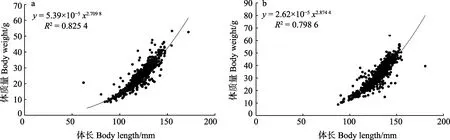
圖7 海陽近岸雌性(a)與雄性(b)口蝦蛄體長–體質量關系Fig.7Relationship between body length and weight of male (a) and female (b) O. oratoria in the coastal waters of Haiyang
2.6 體長相對增長率
雌性口蝦蛄月度體長相對增長速率整體呈先上升后下降的趨勢,4—5月由1.29增加至6.24,10月最高,為36.71,后呈下降趨勢,次年1月最低,為0.92。雄性口蝦蛄月度體長相對增長速率與雌性總體趨勢一致,均在10月達到最高值,次年1月相對增長速率較12月有所增加,由3.49增加至10.01(表2)。
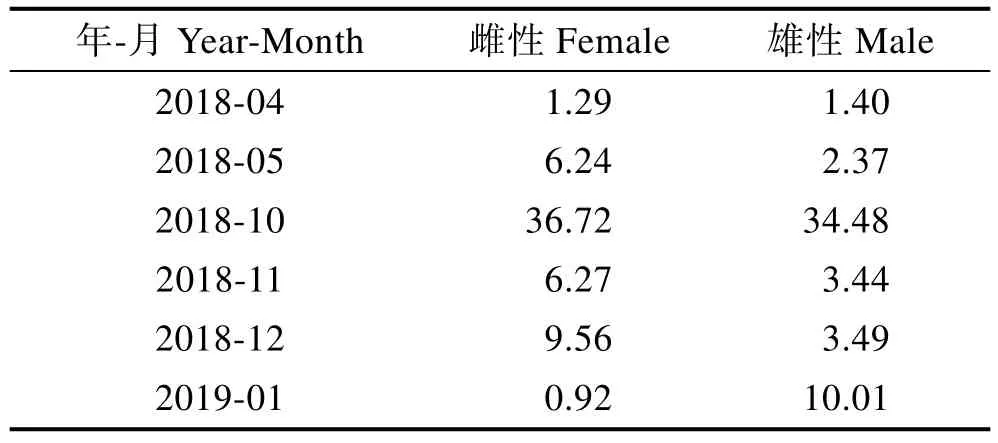
表2 海陽近岸雌性與雄性口蝦蛄體長相對增長速率Tab.2Relative growth rate of body length of male and female of O. oratoria in the coastal waters of Haiyang
3 討論
3.1 關于采樣和計算方法的選擇
甲殼動物的生長需要通過規律性的蛻皮來實現,在蛻皮時,包含年齡信息的鈣化部分會丟失,因此,體長頻率法是漁業上廣為應用的一種可行的世代區分方法(Changet al, 2012)。對于甲殼動物來說,在漁具不具備選擇性的條件下,長度組分布曲線的高峰一般依次降低(李壯,2006),每個高峰代表1個世代,每個高峰的長度則代表著該年齡組的體長范圍。黃海的口蝦蛄群體每年產卵1次(劉海映等, 2013),因此,體長頻率分布圖中的每個高峰,即代表1個年齡組。與一般使用的拖網調查方法不同,本研究采用的是流刺網調查。流刺網雖然具有一定的選擇性,但這種選擇性只針對于一定體長范圍以上的個體。李顯森等(2016)在渤海進行的口蝦蛄三重刺網、單層刺網和底拖網漁獲組成的研究中發現,網目尺寸在50~60 mm的三重刺網和單層刺網捕獲的口蝦蛄體長范圍為78~166mm,而底拖網捕獲的口蝦蛄體長范圍為64~130 mm。流刺網對于體長較大的口蝦蛄個體選擇性較強,小個體不易被捕獲,尤其是1齡以下的,因為口蝦蛄的生物學最小型即初次性成熟體長為80 mm (徐善良等,1996)。根據前期對口蝦蛄拖網面上調查發現,9月拖網捕獲的口蝦蛄優勢體長組為95~110 mm和110~130 mm (圖8),其第2個體長分布高峰與流刺網捕獲的口蝦蛄體長分布高峰重合。根據口蝦蛄的生長曲線(王波等,1998),可判斷優勢體長組95~110mm為當年生的世代,第2個優勢體長組110~130 mm為生長1年后的世代。
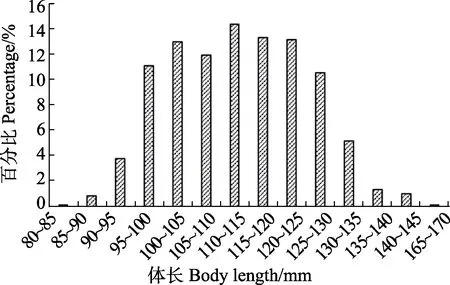
圖8 基于拖網調查的2019年9月海陽近岸海域口蝦蛄體長組成Fig.8Composition of body length of O. oratoria in Haiyang coastal waters based on trawl survey in September 2019
甲殼類應用體長頻率法分析時,常因較大年齡組中發育相對較慢的個體與前一年齡組發育較快的個體體長分布出現重疊,帶來分析誤差(Fournieret al,1991)。為了盡可能避免由此導致對不同年齡組的錯誤判斷,本研究選取優勢體長組中的組中值±5 mm作為體長相對增長率計算樣本的選擇范圍。計算月間體長相對增長速率時,所用的也是該范圍內的樣本的平均長度數值。
3.2 流刺網漁獲中口蝦蛄優勢體長組的生活史
流刺網對口蝦蛄群體的捕獲具有選擇性,其捕獲個體的93.1%以上為1齡以上個體(李顯森等, 2016),因此,本研究對海陽近岸流刺網所捕獲的口蝦蛄進行跟蹤采樣,并根據體長頻率分布劃分各月優勢體長組,分析其生長和攝食情況。
9—10月間,口蝦蛄攝食強度逐漸增強,空胃率逐漸下降(圖6),說明這一階段的口蝦蛄處于大量索餌育肥、加速生長階段。其中,10月雌、雄口蝦蛄體長增長最快(表 1)。口蝦蛄是甲殼動物,其生長需要通過周期性的蛻皮來實現,在蛻去舊的外骨骼前后,攝入水分,使柔軟的新皮膨脹,準確地說,這個過程只是一種膨脹(Lockwood,1968)。因此,9—10月間體長的快速增長表明口蝦蛄很可能正處于階段性蛻皮的過程中,10月中旬絕大多數口蝦蛄樣品外骨骼較軟、殘體較多也證實了這一推測。10月開始,雖然雌性平均攝食強度低于前期,但卵巢逐漸開始發育,Ⅳ期比例由16.90%逐漸增加到次年1月初的82.86%,考慮到流刺網捕獲的雌性口蝦蛄均為1齡以上個體,表明其在秋季有第2個卵巢發育高峰。薛梅等(2016)對皮口海域口蝦蛄的研究也發現,不同年齡口蝦蛄群體在性成熟的時期會有所差異,認為口蝦蛄的性成熟高峰期在5月和11月。口蝦蛄一般壽命為2—3年,經計算,海陽5月和12月2個性成熟高峰期的雌性的優勢組平均體長相近,分別為134.93 mm和130.47 mm,因此,判斷1齡以上性成熟口蝦蛄在春、夏季產卵,未滿1齡口蝦蛄在11月—次年1月期間達到性成熟,但未出現真正意義上的繁殖群體,卵巢發育積累的能量用于育肥越冬,次年5—7月進入真正產卵期。Kim等(2017)對韓國口蝦蛄進行研究時也發現,統營海域口蝦蛄主要產卵季節為夏季,但生殖腺指數在冬季開始升高,12月達到最高,并未發現冬季產卵現象。Ohtomi等(1988)同樣認為,日本東京灣口蝦蛄存在2個性成熟高峰期,但僅有1個繁殖期。
本調查中,1月中旬—3月初,自然海域很難捕獲口蝦蛄,說明其正處于越冬期。根據郭文場等(2011)的研究,越冬期口蝦蛄會在其 U型洞穴中段向地底延伸一段洞穴以抵御低溫,形成近Y字型的洞穴。徐海龍等(2010)認為,黃海北部海域口蝦蛄1月進入越冬期,而青島近岸口蝦蛄12月—次年1月處于越冬期(盛福利等, 2009),浙江北部海域口蝦蛄12月進入越冬期(徐善良等,1996),表明口蝦蛄進入越冬期的早晚跟海域平均水溫有關。越冬期結束后,口蝦蛄分批次進入漁場,開始大量索餌,為繁殖期做準備。生長發育較好的個體率先進入漁場,這也是 3—5月捕獲的口蝦蛄平均體長逐漸降低的原因。海陽近岸雌性口蝦蛄3月生殖腺發育以Ⅱ期為主,4月開始出現Ⅲ期個體,5月Ⅲ和Ⅳ期的個體占到總數的43.48%和17.39%,9月Ⅲ期及Ⅳ期個體消失(圖 5)。說明 3月開始,海陽近岸口蝦蛄生殖腺開始發育,5月進入繁殖旺盛期,8月繁殖進入尾聲。與此同時,3—5月雌、雄性比逐漸減小,一方面,可能因為繁殖期雌性個體在洞穴中的產卵和護卵;另一方面,產卵后高齡雌性個體大量死亡(王春琳等,2002)導致雌性個體減少,被網具捕獲的幾率較低。本研究中,2019年 4月口蝦蛄性比高達2.1,推測是由于產卵前期雌性口蝦蛄大量聚集索餌,導致雌性數量明顯多于雄性。渤海海域口蝦蛄繁殖前期同樣出現雌性個體較多的現象,產卵后性比明顯減小(鄧景耀等, 1988)。盛福利等(2009)對青島近岸口蝦蛄調查發現,5月下旬是口蝦蛄繁殖盛期,與本研究結果較為一致。遼東灣海域口蝦蛄產卵期為6—7月(劉修澤等, 2014),稍晚于本研究結果。推測是由于不同海域水溫及其他環境因素的差異導致口蝦蛄生殖腺發育起始時間和繁殖期持續時間不同,從而影響到生命周期(Ohtomi et al, 1988)。
因此,本研究推斷8—12月是海陽近岸口蝦蛄的生長期,其間10月經歷一次蛻皮,前一年出生的雌性生殖腺開始發育;1—2月為越冬期,3—5月生殖腺再度開始生長發育,5—7月間繁殖。這與王春琳等(1996)對浙江沿岸海域口蝦蛄生活周期的研究基本一致,但產卵階段早于6月中旬。
3.3 流刺網捕撈對海陽近岸口蝦蛄資源利用的分析
鄧景耀等(1992)對渤海海域口蝦蛄進行年齡組劃分,認為30~70 mm為當年生個體,70~110 mm為1齡組,90~150 mm為2齡組,150~175 mm為3齡組。根據本研究優勢體長分布(圖1),流刺網所捕獲的口蝦蛄大多為1齡以上及2齡個體。口蝦蛄1年可達性成熟(王波等,1998),第1次性成熟的生物學最小型為80 mm (徐善良等, 1996),以此為節點劃分,本研究中流刺網口蝦蛄99.88%已達性成熟,且完成了1次產卵。因此,本研究認為,流刺網捕撈對海陽近岸口蝦蛄初生世代資源具有較強的保護作用,對于整個自然群體的補充及延續是有利的。郭瑞等(2016)認為,60 mm三重刺網對遼東灣口蝦蛄具有較強的選擇性,所捕獲的口蝦蛄均大于最小可捕體長110 mm。而李顯森等(2016)在對渤海口蝦蛄三重刺網漁獲組成的研究中,對比了流刺網和拖網漁獲口蝦蛄的群體組成,其中,流刺網捕撈體長組為78~166 mm,拖網體長組成為64~130 mm;優勢體長組分別為120~130 mm和80~90 mm。由此推測,流刺網比拖網對口蝦蛄自然群體的延續更有利。
海陽的近岸口蝦蛄漁業有2個捕撈汛期——春汛和秋汛,春季捕撈以流刺網為主,秋季則以流刺網和拖網為主,這種捕撈方式對海陽當地的口蝦蛄漁業起到了較好的保護作用。根據2014—2016年的調查,海陽各類流刺網漁船的漁獲物中,口蝦蛄占比20.86%,為優勢種(李春芝等,2019)。以《渤海生物資源養護規定》的可捕標準為劃分幼體的標準,2019年 4月及9月對海陽近岸口蝦蛄資源的拖網調查中,4月幼體比例為 26.32%,9月幼體比例為 50.00%。渤海海域拖網口蝦蛄漁獲群體中幼體比例高達83.3%,而流刺網口蝦蛄漁獲中幼體比例僅為5.9%~9.0%(李顯森等,2016)。與渤海海域相比,海陽近岸拖網口蝦蛄漁獲的群體結構中,幼體比例明顯偏低,推測其群體結構更有利于自然群體的延續。
由此可見,流刺網較拖網能更好地保護海陽近岸口蝦蛄幼體,有利于其群體延續及漁業資源的持續發展。因此,本研究認為,應提倡流刺網捕撈代替拖網捕撈,盡量降低拖網捕撈對口蝦蛄資源造成的不利影響,減輕口蝦蛄自然群體的生存壓力,實現海陽近岸口蝦蛄漁業資源的可持續利用。
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
趣味(語文)(2020年3期)2020-07-27 01:42:46
中國生殖健康(2019年2期)2019-08-23 08:12:08
產品可靠性報告(2017年7期)2017-09-05 09:49:12
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
汽車觀察(2016年3期)2016-02-28 13:16:26
中國衛生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
