糜子不同品種的光合能力和源庫關系類型對產量形成影響的研究
2022-01-15 07:22:04游文萍張東升劉昭霖宗毓錚郝興宇
激光生物學報 2021年6期
關鍵詞:產量
游文萍,張東升,劉昭霖,宗毓錚,郝興宇,李 萍
(山西農業大學農學院,太谷 030801)
源庫關系決定著作物產量的形成,因為作物葉面積和光合作用會通過影響作物對光的捕獲及利用進而影響作物碳水化合物的生產,這也常用于表征作物源的大小。而作物產量的形成(庫建成)主要受到光合碳同化物向果實器官累積分配[如收獲指數(harvest index,HI)]的影響[1-4]。之前對主要糧食作物育種的關注度在于作物HI的提高,而且相關研究幾乎已經達到了一個上限值。在當前作物育種過程中,提高作物產量的合理途徑被認為是調控作物光合作用過程和源庫關系,但這需要我們深入了解作物光合作用和源庫關系的遺傳變異,并明確它們與作物產量形成的關系。
作物光合作用受其遺傳特性、環境因素和管理措施的影響,也會受到作物源庫關系的反饋調節[1-2,5-6]。在育種中,葉片光合速率[如最大凈光合速率(Pmax)和凈光合速率(Pn)]的遺傳變異通常被作為選擇優良作物品種的前提。然而由于光合作用的多重效應,盡管某些物種(如小麥、玉米、大豆和鷹嘴豆)的葉片光合速率與產量存在正相關關系,但它們的產量并未顯著增加[7-12]。就光合性狀而言,小麥和棉花的氣孔導度與產量的相關性最強[12-13]。一般來說,光合遺傳進化也常常伴隨著光合速率和蒸騰作用對氣孔限制的降低。氣孔導度通常控制著蒸騰作用中碳吸收和水分損失之間的平衡,從而影響水分利用效率。而水分利用效率由于其強大的遺傳基礎,一直是作物選擇和育種的目標性狀[3-4,14-17]。此外,光合作用還受非氣孔因素[如光系統II(photosystem II,PSII)]的光化學效率的限制[18]。葉綠素熒光動力學作為PSII光化學效率的一種非常靈敏和內在的探針,在植物生理學各個領域內均有廣泛應用[19-21]。葉綠素熒光參數中的最大光化學量子效率(Fv/Fm)值已被廣泛用于測定并評價逆境條件對光合作用的影響。實際光化學量子效率(ФPSII)值可以作為電子通過PSII傳遞的定量指標,這與PSII的光化學效率有關[22]。光化學猝滅系數(qP)通常用來表示反應中心的開放程度[23]。非光化學猝滅系數(qN)反映了光周期中非光化學過程的激活,如反式類囊體pH梯度、光抑制過程和非輻射能量耗散熱[24]。
糜子(Panicum miliaceumL.)作為東亞地區過去10 000年里一直種植的作物[25],其所含營養極為豐富,主要富含膳食纖維、儲存蛋白、抗性淀粉、必需氨基酸、維生素和其他生物活性物質[26-27]。糜子對水分和營養的低需求使得它能夠在中國北部、中亞和俄羅斯等地區半干旱貧瘠的環境中生長[28-29]。目前遺傳改良在減輕干旱條件下作物產量損失方面做出了巨大貢獻[30],前提是需要我們明確影響作物產量形成的遺傳策略和篩選相關遺傳性狀。盡管C3(如小麥和棉花)和C4(如玉米)作物的育種已取得了更多進展[12,17,31-33],但未來C4糜子高產抗逆優良品種的選育仍然需要我們更深入探究糜子光合及其源庫關系的遺傳變異規律,并明確它們與糜子產量形成的關系。
1 材料與方法
1.1 試驗地點
田間試驗于2012年在北京昌平中國農業科學院(北緯40.13°,東經116.14°)試驗站進行。該區氣候為受季風影響的大陸性氣候,春季干燥,夏季炎熱(k?PPen-geiger分類[34])。糜子生長季時的降水量為(286±16)mm,氣溫為(22.9±3.2)℃。土壤質地為褐色潮土(pH值8.3),有機質含量為12.30 g/kg,土壤全氮含量為0.81 g/kg,土壤總磷含量為0.69 g/kg,土壤全鉀含量為14.50 g/kg。
1.2 試驗詳情
本試驗選用生長在我國半干旱地區不同地方的9個糜子品種作為供試試驗品種,分別為內蒙古紅糜子(M1)、雁黍7號(M2)、伊糜5號(M3)、紫桿白黍(M4)、靖遠中集青糜(M5)、白黍子(M6)、寧糜14(M7)、一點青(M8)、昌圖紅糜子(M9),它們均來自中國北京農作物種質資源中心。試驗采用完全隨機設計,重復3次,每組地塊面積為30 m2。糜子種植密度為80株/m2,行距為0.3 m,其在糜子整個生長季節均不額外灌溉和施肥。
1.3 氣體交換測量
使用便攜式氣體交換系統(LI-COR 6400;LI-COR,Lincoln,Neb.)進行氣體交換測量。葉室內的CO2濃度由LI-COR的CO2注射系統控制在400 μmol/mol,光量子通量密度(photosynthetic photon flux density,PPFD)為1 600 μmol/m2/s,由內置的LED燈(紅/藍)提供,葉室溫度設定為25℃。在營養生長和生殖生長過渡期(不同品種糜子播種后的40 d左右),于晴天上午9:00—11:30在作物冠層內隨機選取6片最上部完全展開的糜子旗葉,測定其Pn、氣孔導度(gs)、蒸騰速率(E)和胞間 CO2濃度(Ci)等氣體交換參數,然后計算糜子的氣孔水汽壓(gsw=1.6×gs),由此計算單葉水分利用效率(WUE=Pn/E)和內在水分利用效率(iWUE=Pn/gsw)。另外在每個糜子樣品上取3片完全展開的旗葉,測定不同品種糜子的光響應曲線。葉室內CO2濃度為400 μmol/mol,初始PPFD設置為1 600 μmol/m2/s,依次為1 300、1 100、900、700、500、300、200、100、50 μmol/m2/s,在光合速率達到穩定狀態時進行數據記錄。
1.4 葉綠素熒光測量
在測定氣體交換參數的同時,利用微型脈沖振幅調制熒光分析儀(Mini-PAM,Walz,Effeltrich,Germany)測定相同樣葉的葉綠素熒光參數[Fv/Fm、ФPSII、電子傳遞速率(electron transport rate,ETR)、qP和qN],評估葉片PSII的光合性能[35-36]。所有的葉綠素熒光參數根據Rascher等[37]和Huang等[38]描述的公式進行計算。
1.5 葉面積、干物質量、產量和產量構成因素
測定了氣體交換和熒光動力學參數后,計數供試糜子品種單株葉片數量,測定葉片長度(length,L)和寬度(width,W),根據LA=L×W×a(a表示糜子的形狀系數,本試驗中為0.75)計算單株葉面積(leaf area,LA)。在成熟期,糜子于2012年9月11日進行人工收獲。風干后,從各處理樣品中隨機抽取10株糜子,測定株高、穗長、單株籽粒數和籽粒千粒重。通過每株籽粒重和整株干重計算單株產量和干物質量。HI是單株產量和單株干物質量的比值。
1.6 統計分析
本試驗所有統計分析在統計軟件R(https://cran.r-project.org/)平臺進行。方差分析為單因素ANOVA分析,多重比較采用P=0.05水平的最小顯著性差異檢驗。變異系數為平均值與標準差之比。用非線性混合效應模型擬合實測的光響應數據。選用非直角雙曲線函數,并利用R中的nlme包對其參數進行估計。采用R中的corrplot包進行Pearson相關性分析以明確氣體交換參數、熒光動力學參數、葉面積、干物質量、產量以及產量構成因素間的相關性,通過t檢驗確定相關分析的顯著性差異(P≤0.05)。
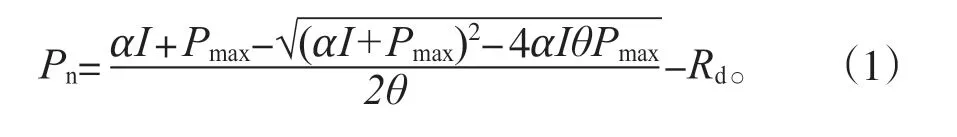
其中Pn為葉片凈光合速率(μmol/m2/s),I是被吸收的PPFD(μmol/m2/s),估算為入射PPFD與葉片吸收系數0.88的乘積。Pmax是指葉片最大凈光合速率(μmol/m2/s),α是表觀量子效率,θ是曲率參數,反映光合曲線彎曲程度的凸度,Rd是暗呼吸(μmol/m2/s)。
2 結果與分析
2.1 Pmax和氣體交換參數
用非直角雙曲線函數擬合光合作用對光的響應,結果表明,R2大于0.9時,擬合效果良好(圖1)。不同品種糜子的Pmax和氣體交換參數有顯著差異,而α和Rd差異不顯著(表1和表2),表明糜子利用強光的能力具有明顯的遺傳變異。就氣體交換參數而言,3個糜子品種M6、M8和M9的Pn相對最高,為高光合品種。而M1和M2的Pn相對最低,為低光合品種。M3、M4、M5和M7的光合能力處于中等水平,為中等光合品種(表2)。Pn與gs(P<0.05)和Ci(P<0.05)相關,而gs與E和WUE有較好的相關性(圖2)。
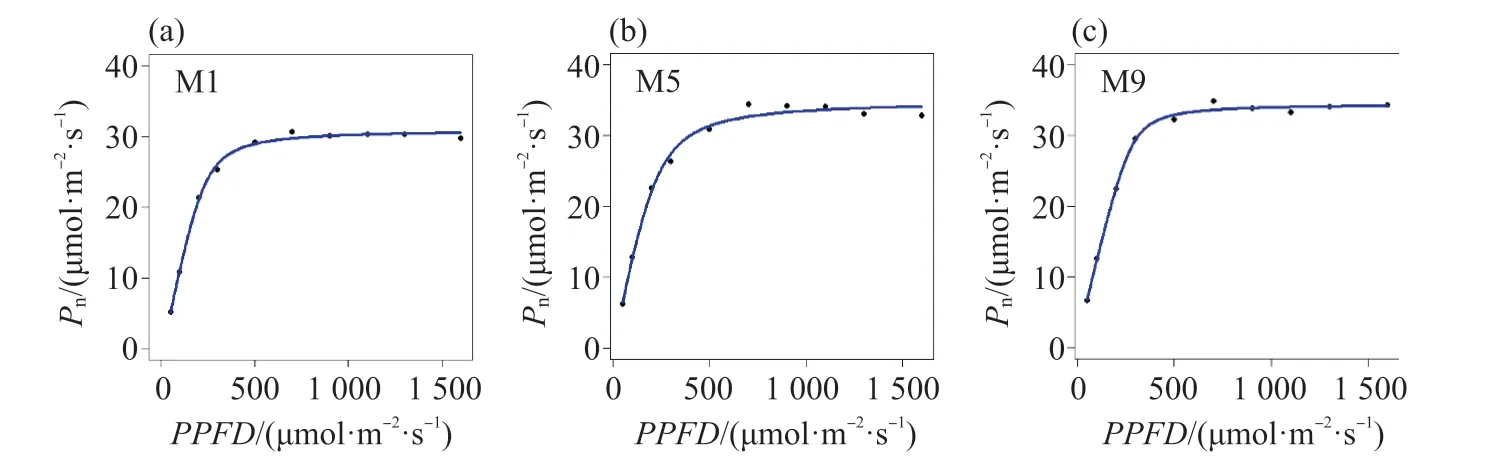
圖1 不同品種糜子的光響應曲線Fig.1 Photosynthetic light response curves of different broomcorn millet varieties
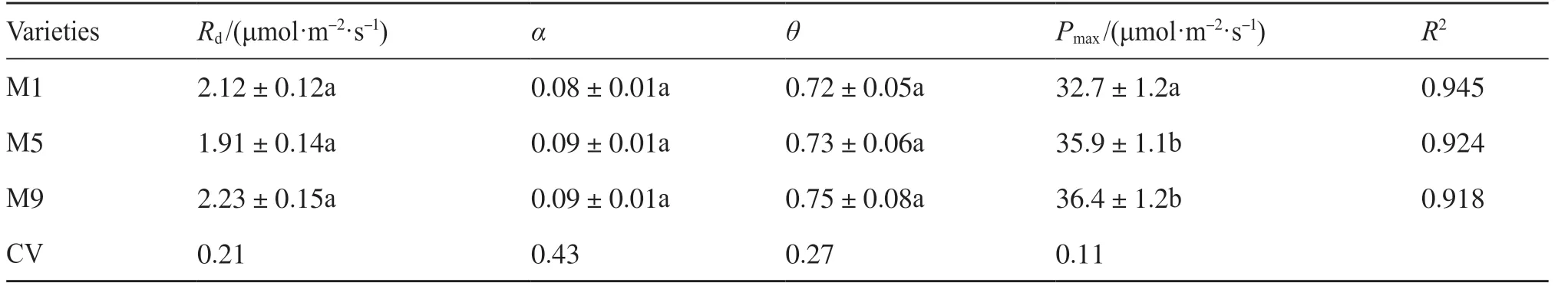
表1 不同品種糜子的光響應曲線參數Tab.1 The parameters of photosynthetic light response curve for different broomcorn millet varieties
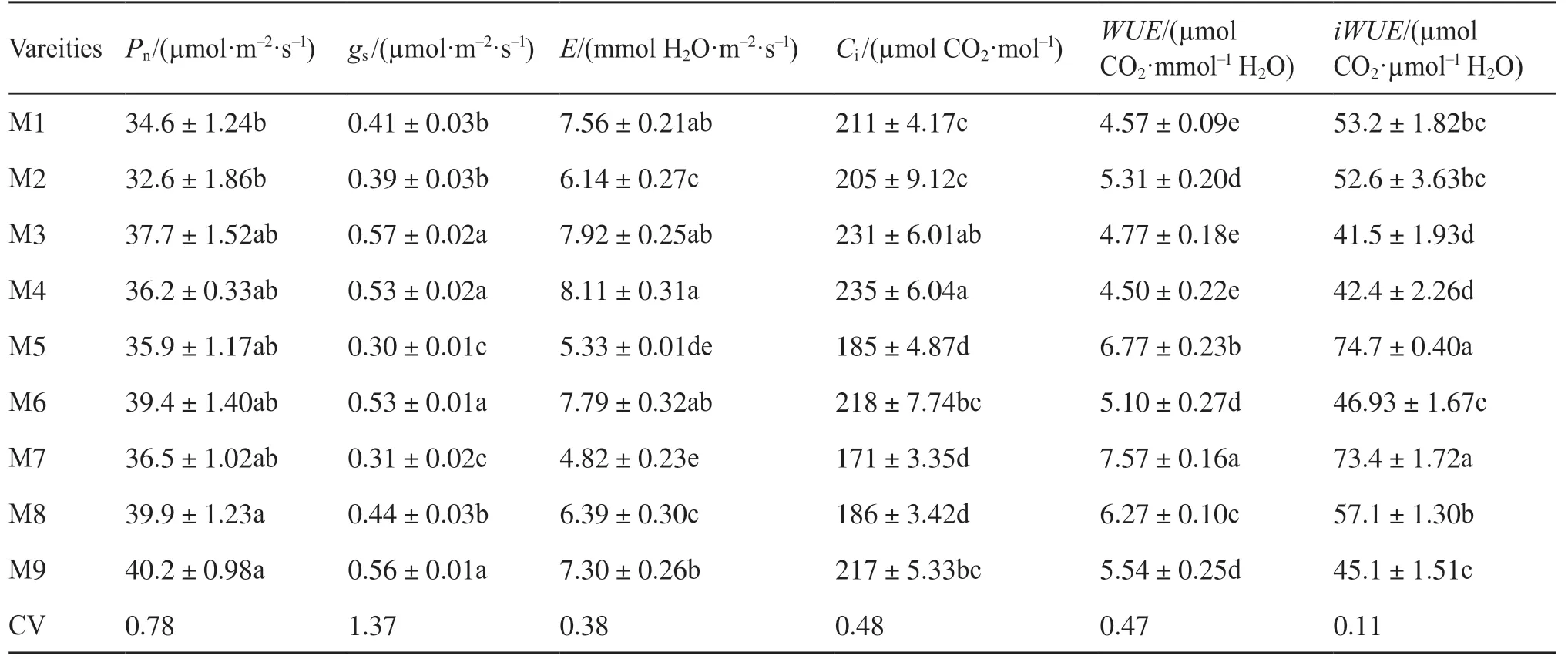
表2 不同品種糜子的氣體交換參數Tab.2 The gas exchange parameters of different broomcorn millet varieties
2.2 葉綠素熒光參數、光合、葉面積和HI與產量的相關性
不同品種的糜子PSII光化學效率具有顯著的差異。除Fv/Fm和qN外,糜子的ФPSII、ETR和qP表現出顯著的品種差異(表3)。在所有測試的糜子品種中,Pn與ETR、ФPSII和qP呈正相關(P< 0.05),E與ETR、ФPSII呈顯著負相關(P< 0.01)(圖2)。
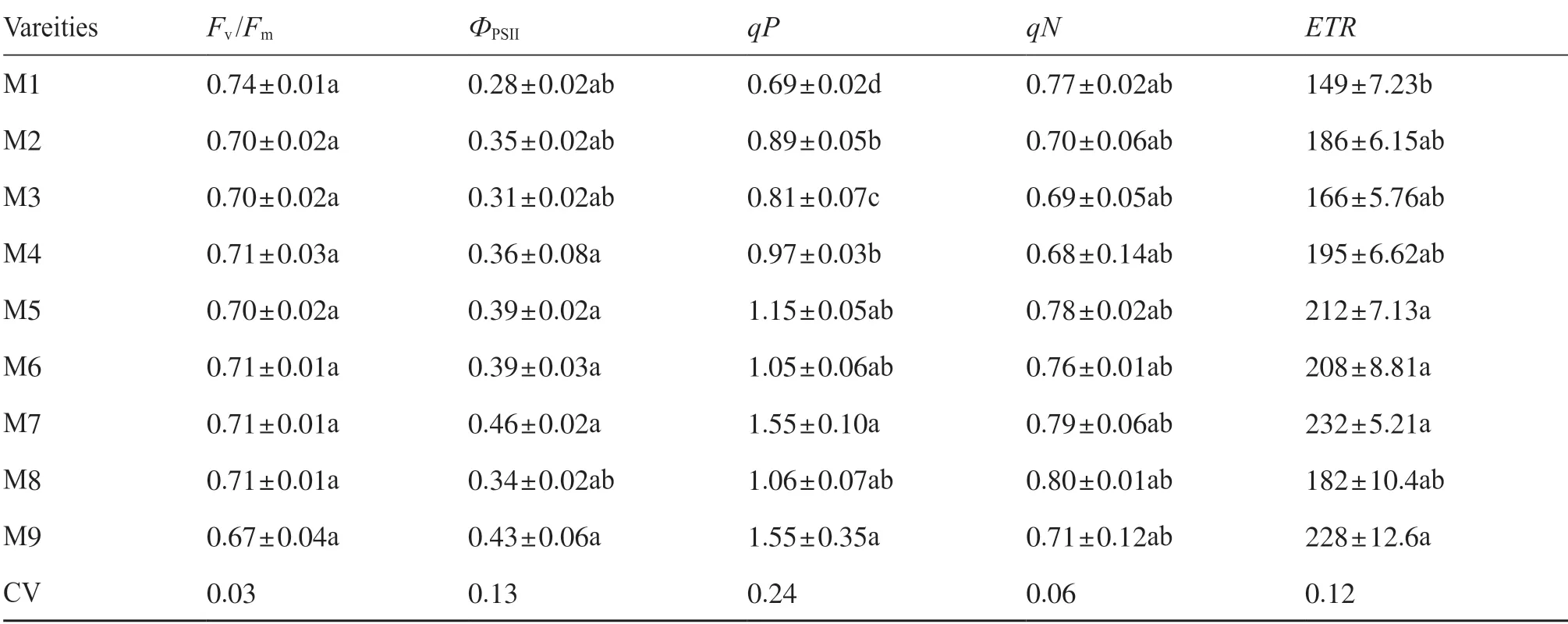
表3 不同品種糜子的葉綠素熒光參數Tab.3 The chlorophyll fluorescence parameters of different broomcorn millet varieties
在所有供試品種中,糜子單株產量與HI、單株干物質量、穗長、單穗粒數呈極顯著正相關,與粒重無顯著相關性。不同品種糜子的單株干物質量和產量與Pn、gs、Ci和E均呈顯著負相關,與iWUE、WUE、qP、qN和ФPSII呈顯著正相關。糜子單株干物質量與單株葉面積呈現顯著正相關(圖2)。
2.3 葉面積、株高、干物質量和HI
葉面積和株高影響植株的光截獲,進而影響碳水化合物的生產。糜子單株葉面積以M2和M3顯著最大,M1和M9顯著最小,M4、M5、M6、M7和M8居中(圖3a)。糜子株高以M1、M2、M7和M8顯著較高,M9最低,其他品種的株高居中(圖3b)。產量由干物質量和HI的值決定。M5、M7和M9的干物質量顯著較大,而M1、M3和M6顯著較小,其他品種居中(圖4a)。糜子的HI值以M1、M3、M6、M9顯著較低,M4、M5、M7、M8顯著較高(圖4b)。
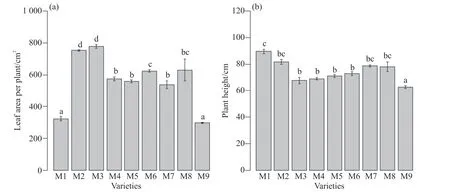
圖3 不同品種糜子的單株葉面積和株高Fig.3 The leaf area per plant and plant height of different broomcorn millet varieties
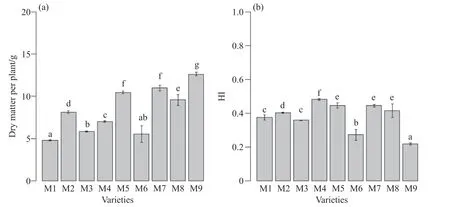
圖4 不同糜子品種單株干物質量及HIFig.4 The dry matter per plant and HI of different broomcorn millet varieties
2.4 產量和產量構成因素
單株產量與穗長、單穗粒數呈極顯著正相關,與粒重無顯著相關性(圖2)。產量和每穗粒數的變異系數分別為0.39和0.38,高于粒重的變異系數0.05。單穗粒數越少,單株產量越低,如M1、M3、M6、M9,單穗粒數均小于400粒,單株產量均小于3.0 g。單穗粒數多的品種單株產量高,特別是M5和M7,單穗粒數大于600粒,單株產量最高(表4)。
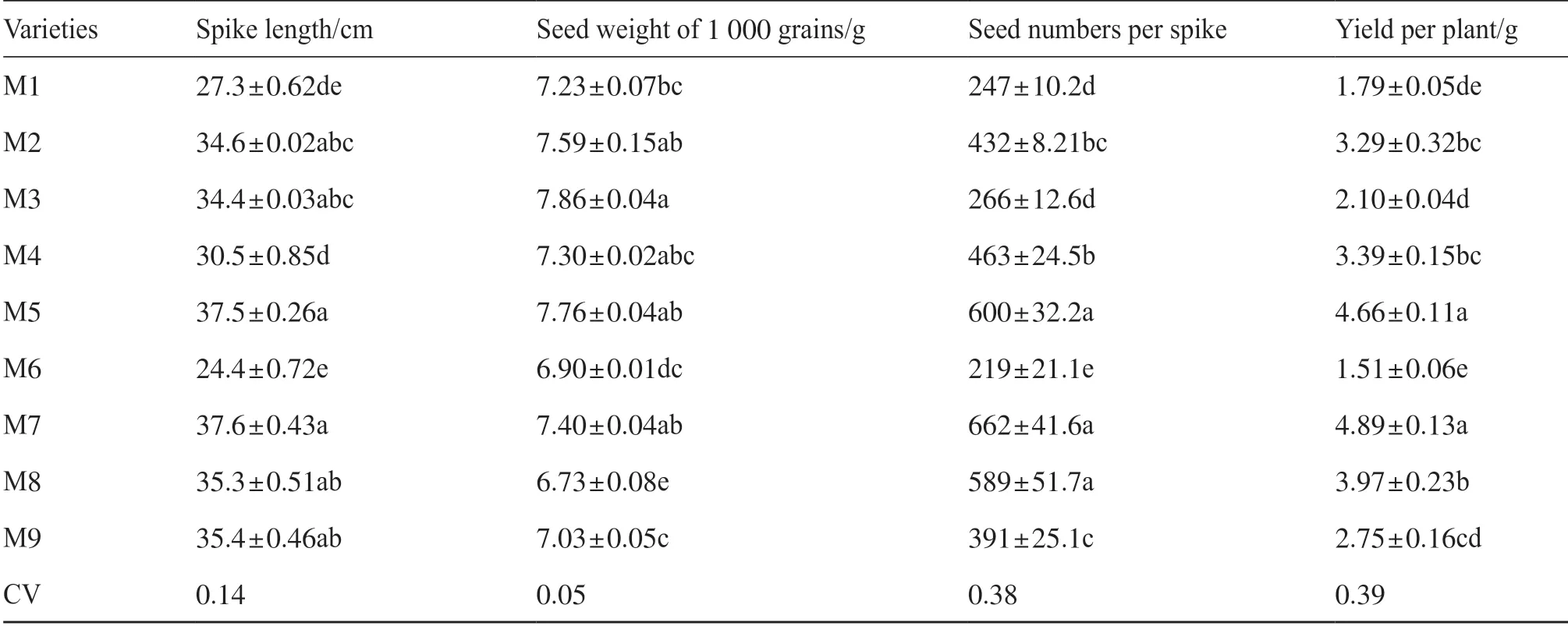
表4 不同品種糜子的產量構成因素及產量Tab.4 Yield components and yield of different broomcorn millet varieties
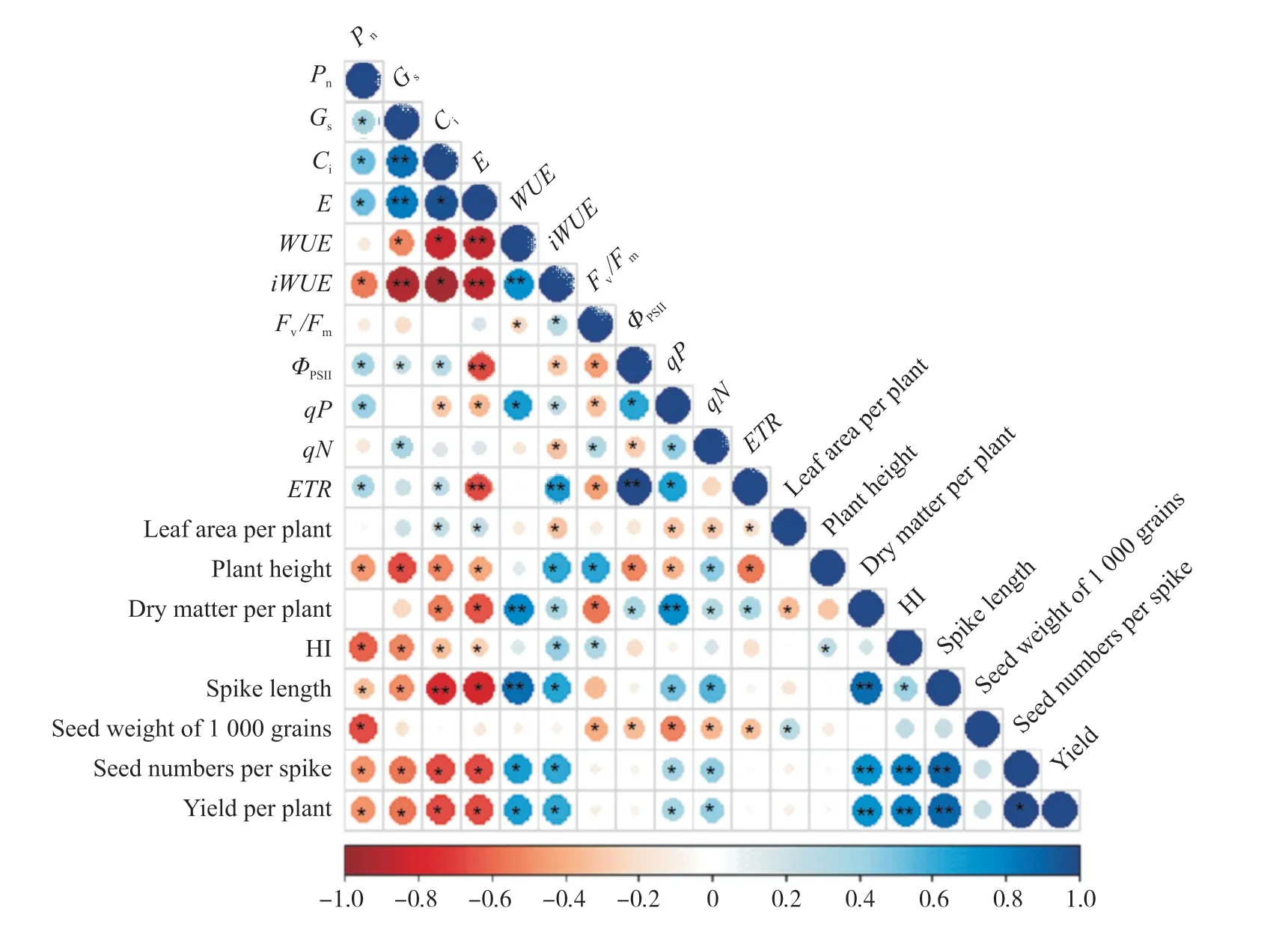
圖2 不同品種糜子的氣體交換參數、葉綠素熒光參數、單株葉面積、株高、單株干物質量、HI、單株產量及產量構成因素之間的皮爾遜相關系數(**P<0.01;*P<0.05)Fig.2 Pearson correlation coefficient between gas exchange parameters, chlorophyll fluorescence parameters, leaf area per plant, plant height, dry matter per plant, harvest index, yield per plant and yield components of different broomcorn millet cultivars (**P<0.01; *P<0.05)
3 討論
光合作用是作物碳水化合物形成的唯一途徑,在作物生長發育過中影響著碳水化合物的形成,也與產量有著密切關系[39-44],通過提高作物光合能力進而增加生物量和產量已經成為未來育種的主要目標。作物Pn和Pmax常常被認為具有一定的遺傳變異能力,能夠作為作物品種選育的關鍵光合性狀。一些研究發現,作物的Pn與產量之間存在顯著正相關,但這種關系似乎并不穩定,因為也有研究表明,包括小麥、水稻、大豆和棉花等這些作物的凈光合速率與產量缺乏正相關關系或呈現負相關關系[12]。在本研究中我們發現,Pn和Pmax的確表現出明顯的遺傳變異,但它們與單株產量呈顯著負相關(圖2)。該結果可能與葉面積引發的多效性有關,因為與單葉光合相比,作物產量與作物冠層光合能力表現出了更好的正相關關系[12]。WUE,尤其是iWUE,也一直被認為是作物品種選育的關鍵目標。許多研究都發現iWUE與作物產量之間存在穩定的相關性[45]。本試驗中,糜子的WUE和iWUE與單株產量表現出顯著正相關,這可能意味著水分利用效率(如WUE或iWUE)可以作為糜子品種選育的關鍵性狀。另外,盡管有研究表明,糜子的Pn與gs和Ci有顯著相關性[43],但我們的試驗發現糜子gs與E的相關性比gs與Pn的相關性更好,這意味著干旱地區糜子gs對水的敏感程度可能高于對CO2的敏感程度[46-48]。
植物源庫關系影響植物的生理代謝過程,從而影響同化物的生產和運輸。強源可以給庫提供更多的光合產物,而強庫可以調節源產生代謝物的輸出速度和輸出方向[49-50]。因此,源庫關系是影響作物碳水化合物生產和分配的重要因素,最終會影響作物產量的形成。在當前研究中,糜子源庫關系表現出明顯的品種依賴性。一些品種源弱庫強,產量較低,如內蒙古紅糜子(M1)的葉面積和葉光合速率較小,源限明顯,干物質累積較低,盡管HI相對較大,但單株粒數仍較少,產量較低。一些品種源強庫弱,產量較低,如白黍子(M6)的光合速率和單株葉面積都相對較高,而昌圖紅糜子(M9)光合速率最高,它們干物質累積多,但由于HI和穗粒數較小,即受到庫強度的限制,其產量也相對較小。一些源庫關系較好的品種,如靖遠中集青糜(M5)和寧糜14(M7),雖然它們的源強(葉光合速率和葉面積)和庫容(HI)并不是最大的,但是有相對較多的光合同化物在果實中分配,使得它們籽粒數最多,因此產量也最高。
本研究對不同品種糜子的光合生理作用和源庫關系進行了相關試驗和分析,為糜子的研究提供了一定的理論依據,但源庫關系與產量的關系是比較復雜的過程,中間還有很多生理活動需要我們更深入細致的研究。這有待我們在今后的研究中開展更深入更細致的科學試驗,為糜子的生產和育種工作提供保障。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
