山茱萸果實發育過程中單寧物質的分布與積累特征
2022-01-15 05:35:16陳旋勐顏永剛
西北植物學報 2021年11期
陳旋勐,張 崗,高 靜,顏永剛,魏 瑤,陳 瑩
(陜西中醫藥大學 藥學院/陜西省秦嶺中草藥應用開發工程技術研究中心,陜西咸陽 712046)
山茱萸為山茱萸科植物山茱萸(CornusofficinalisSieb.et Zucc.)的干燥成熟果肉,又稱山萸肉,是中國大宗道地中藥材品種之一[1],主產于陜西、山西、河南、甘肅等地[2]。山茱萸味酸澀,性微溫,具有補益肝腎,收澀固脫等功效,常用于治療眩暈耳鳴、腰膝酸痛、陽痿遺精、遺尿尿頻、崩漏帶下、大汗虛脫、內熱消渴等癥[3]。山茱萸化學成分主要包含多糖、有機酸、酚類、苷類、環烯醚萜苷、皂苷、鞣質等[4-5]。現代藥理學研究表明,山茱萸在抗炎、糖尿病的防治、心腦血管的保護、神經保護、抗腫瘤、抗氧化和抗衰老等方面均表現出較好的活性[5-7]。
單寧,又名單寧酸、鞣酸,是廣泛存在于植物體內的次生代謝產物,在植物防紫外線以及抵御病原體侵害等方面都發揮著重要的作用[8]。除此之外,單寧物質能與人體口腔黏膜蛋白結合產生強烈的收斂性,引起“澀感”,過量的單寧攝入后,單寧會與唾液蛋白酶及腸胃中的膳食蛋白質發生交聯,導致蛋白質代謝紊亂,影響蛋白質的可消化性和利用率[9];但單寧的適量攝入對多種酶和病菌具有明顯的抑制作用[10]。單寧的存在是導致果實澀感的主要原因,與山茱萸果實“味酸澀,性微溫”的性味特點相一致。目前,對山茱萸的研究主要集中在炮制加工、化學成分的提取測定、質量評價、藥理作用及臨床應用等方面[11-14],而對于山茱萸果實中單寧物質的研究報道較少。廖景平等[15]在對山茱萸果皮的解剖學研究中提到單寧主要積累在山茱萸果實中果皮體積膨大的薄壁細胞,即單寧細胞中。喬琦等[16]研究發現山茱萸中果皮的色素塊中積累了高濃度的皂甙類和多糖等藥用成分。植物中單寧的含量會受到遺傳、環境變量以及植物成熟度等多方面的影響[17-20]。但是,對于山茱萸果實發育過程中單寧細胞發育及單寧物質的積累形式仍缺乏深入研究,本實驗選取不同發育時期的山茱萸果實作為研究對象,對山茱萸果實發育過程中單寧物質的分布、單寧含量和單寧細胞數目變化以及單寧物質積累特點進行探究,為深入研究山茱萸果實澀味調控機制奠定實驗基礎。
1 材料和方法
1.1 實驗材料
山茱萸果實為2018年3月至今采自于陜西中醫藥大學藥用植物園15年生的健康山茱萸植株,經陜西中醫藥大學顏永剛教授鑒定為山茱萸科山茱萸屬的山茱萸。
1.2 實驗方法
1.2.1 果實形態觀察在光學顯微鏡Nikon SMZ745T下對山茱萸果實發育過程中果實的外部形態及內部結構變化進行觀察并拍照記錄。
1.2.2 石蠟切片法使用50% FAA固定液對所采集的山茱萸果實固定,梯度酒精逐級脫水,二甲苯透明,浸蠟,包埋,Lecia RM2135型手搖式切片機切片,切片厚度8 μm~10 μm,在40 ℃的電熱鼓風干燥箱中烘片,番紅-固綠染色,中性樹膠封片,在Leica DM750 型光學顯微鏡下觀察,并拍照記錄。
1.2.3 組織化學染色單寧顯色反應:三氯化鐵中的鐵離子能夠與單寧中的酚羥基結合,發生顯色反應。取脫蠟后的山茱萸果實縱切片,在三氯化鐵溶液中浸染25~30 min,用Leica DM750 型顯微鏡觀察染色結果,并拍照記錄。用以鑒別單寧物質在山茱萸果實中的分布。
過碘酸-Schiff試劑在組織學上可以用來檢測組織中的多糖物質,高碘酸是一種氧化劑,能夠將多糖分子中相鄰的二醇基氧化成二醛基,醛基能與Schiff試劑反應生成紅色不溶性復合物。取脫蠟后的山茱萸果實縱切片,在過碘酸液中浸染10 min,自來水沖洗10 min,Schiff試劑染色10 min,流水沖洗5 min,用Leica DM750 型顯微鏡觀察染色結果,并拍照記錄。
1.2.4 透射電鏡法使用Adobe Photophoto CC作圖示意山茱萸果實透射電鏡的取樣部位,將不同發育時期山茱萸果實中果皮部分果肉(圖1所示區域)切成2~3 mm的小塊,使用2.5%的戊二醛固定液(pH=7.0的磷酸緩沖液配置)4 ℃預固定24 h,然后使用磷酸緩沖液漂洗3次,每次20 min,0.5%的鋨酸(pH=7.0的磷酸緩沖液配置)固定2 h,用磷酸緩沖液漂洗3次,每次20 min,梯度酒精脫水,環氧丙烷過渡,Epon812包埋劑包埋,60 ℃烘箱聚合24 h,使用Leica EM UC7超薄切片機切片,切片厚度為1~2 μm,制成半薄切片,經甲苯胺藍染色,Leica DM750顯微鏡觀察并拍照。根據半薄切片定位情況,使用Leica EM UC7超薄切片機對包埋好的樣品進行切片,切片厚度為60~80 nm,經醋酸雙氧鈾和檸檬酸鉛雙染色,在日立透射電子顯微鏡H-7650下觀察并拍照。
1.2.5 含量測定可溶性單寧含量測定采用Folin-Ciocaileu法,參考Ikegami[21]和張凡[22]等的方法。取5.0 g山茱萸果肉,以80%甲醇溶液為提取液,按照樣品質量(g):提取液體積(mL)為1∶4的比例處理樣品,充分勻漿后轉入EP管中;5 000 r/min,常溫離心10 min,取上清液至50 mL容量瓶中,以20 mL 80%甲醇清洗殘渣,重復上述步驟,在50 mL容量瓶中定容;隨后移至500 mL的容量瓶中定容將其稀釋10倍;取1 mL樣品,加入7.5 mL蒸餾水和0.5 mL酚類試劑,3 min后加入1 mL飽和碳酸鈉,1 h后在746 nm下測定吸光度,由單寧酸標準曲線計算對應單寧含量。
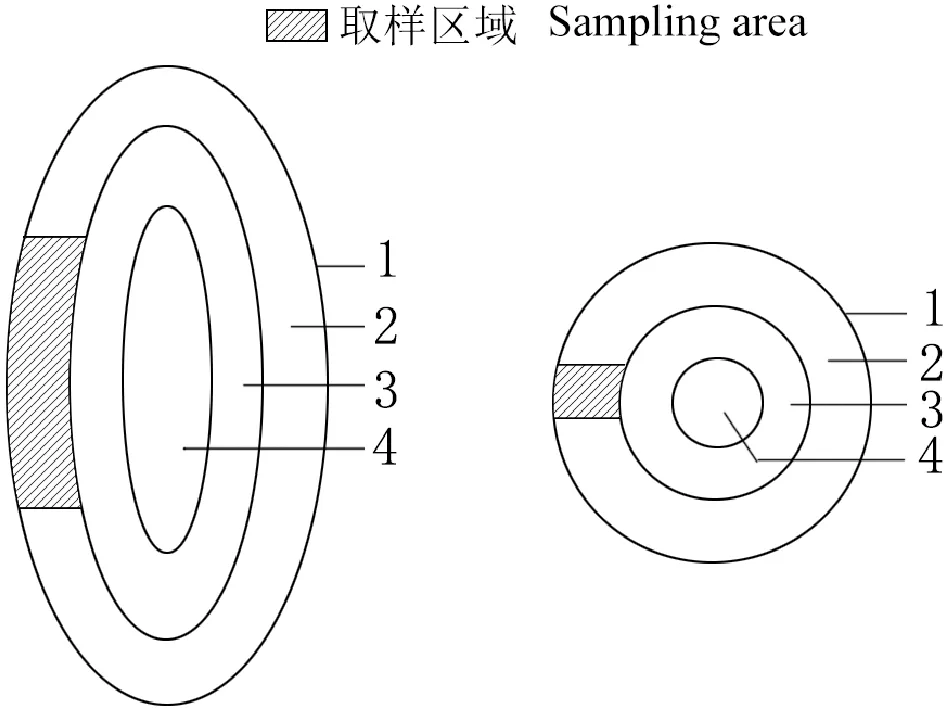
1. 外果皮;2. 中果皮;3. 內果皮;4. 種子圖1 透射電鏡取樣部位示意圖1. Exocarp; 2. Mesocarp; 3. Endocarp; 4. SeedsFig.1 Schematic diagram of sampling position of transmission electron microscope
不溶性單寧含量測定參考Oshida等[23]的方法。用1%(v/v)HCl-甲醇溶液溶解測定完可溶性單寧含量的果肉殘渣,靜置30 min,5 000 r/min、常溫離心10 min;取上清液,用1%(v/v)HCl-甲醇溶液再次清洗殘渣,重復以上步驟;最后取上清液定容到50 mL,后續測定步驟與可溶性單寧含量測定方法相同。
1.2.6 數據分析用Excel 2016、Graphpad prism 8、SPSS 25.0進行數據統計和差異顯著性分析。
2 結果與分析
2.1 果實形態結構觀察
對山茱萸果實的外部形態以及內部結構觀察并結合果實性狀測定結果,將山茱萸果實的發育劃分為幼果期、中果期、成熟期3個時期。
幼果期(開花至花后90 d)果皮顏色為綠色,果實縱、橫徑差異明顯,果實縱徑最大為14.6392 mm,果實橫徑最大為6.4728 mm (圖2);中果期(花后90~180 d)果皮顏色在較長一段時間內為綠色,后期果皮顏色由綠轉黃,果實橫徑較幼果期末期增加了2.3828 mm(圖2);成熟期(花后180~210 d)果皮顏色由黃色逐步轉為紅色,全紅之后果皮顏色進一步加深,果實縱、橫徑差異較小,果實縱徑保持在14.3076~14.6376 mm,果實橫徑8.6656~8.8556 mm (圖2)。
2.2 單寧物質的組織化學定位
山茱萸果實為核果,主要由果皮和種子兩部分組成,果皮由外到內依次是外果皮、中果皮和內果皮(圖3,A)。山茱萸果實外果皮由一層排列整齊的矩形細胞組成。中果皮體積膨大的薄壁細胞中積累的棕色物質經三氯化鐵染色后顯藍黑色 (圖3, C),表明含有單寧;經過碘酸-Schiff試劑反應顯色呈紫紅色(圖3,D),表明含有多糖。在山茱萸果實內果皮中有一些細胞體積較大,與周圍細胞差異明顯,即異細胞[15],它是由內果皮特化的細胞發育而來的,縱切面觀其形態呈橢圓形或圓形。
2.3 單寧含量及單寧細胞數目變化
對不同發育時期山茱萸果實縱切觀察結果如下:
幼果期(開花至花后90 d):幼果期早期山茱萸果實中果皮薄壁細胞大小約為0.0119 mm,尚未發現有單寧物質的積累(圖4,A)。隨著果實進一步發育,中果皮體積膨大的薄壁細胞中逐漸開始積累單寧物質(圖4,B),單寧物質經番紅-固綠對染呈紫紅色,單寧細胞數目也不斷增多(圖4,B-D)。幼果期總單寧和可溶性單寧含量變化呈增長趨勢,不溶性單寧含量變化不明顯(圖5)。其中開花后30~60 d總單寧含量平均日增長速率為48.3836%,可溶性單寧含量平均日增長速率為48.222%;花后60~90 d,總單寧含量的平均日增長速率為8.455%,可溶性單寧含量平均日增長速率為9.5370%。

圖2 不同發育時期山茱萸果實形態結構變化Fig.2 Changes of morphological structure of C. officinalis fruit at different development stages

Ep. 外果皮;Me. 中果皮;En. 內果皮;Pa. 薄壁細胞;TC. 單寧細胞;Id. 異細胞;黑色箭頭→指示單寧細胞;A. 番紅-固綠染色,示山茱萸果實縱切面結構;B. 未染色,示陰性對照;C. 三氯化鐵染色,示單寧物質染色結果呈藍黑色;D. PAS反應,示單寧物質染色結果呈紫紅色圖3 山茱萸果實中單寧細胞分布Ep. Epicarp; Me. Mesocarp; En. Endocarp; Pa. Parenchyma cell; TC. Tannin cell; Id. Idioblast; Black arrow indicates tannin cell;A. Saffron-fast green staining shows the longitudinal section structure of C. officinalis fruit; B. No staining, showing negative control; C. FeCl3 staining showed that tannin were black-blue; D. PAS reaction showed that tannin was purplish redFig.3 Tannin cell distribution in C. officinalis fruit
中果期(花后90~180 d):山茱萸果實中單寧細胞的數目減少(圖4,E)。總單寧和不溶性單寧的含量均呈先增后減的趨勢,可溶性單寧的含量不斷減少(圖5)。其中花后90~150 d可溶性單寧的含量變化與不溶性單寧的含量變化呈負相關。可溶性單寧含量的平均日減速率為9.9570%,不溶性單寧含量的平均日增速率為11.8882%;花后150~180 d總單寧含量、可溶性單寧含量和不溶性單寧含量均呈下降趨勢,其中總單寧含量的下降速率最大為17.6180%。
成熟期(花后180~210 d):山茱萸果實單寧細胞數目減少(圖4,F)。總單寧、可溶性單寧和不溶性單寧的含量均呈下降趨勢(圖5),其中,總單寧和可溶性單寧含量的平均日下降速率均大于不溶性單寧含量的平均日下降速率,總單寧、可溶性單寧和不溶性單寧平均日下降速率分別為31.3680%、23.9127%和7.4553%。

不同小寫字母表示花后天數間存在顯著差異(P<0.05)圖5 山茱萸果實發育過程中單寧含量變化Different normal letter means significant difference among days after flowering (P<0.05)Fig.5 Changes of tannin content during fruit development of C. officinalis

CW. 細胞壁;V. 液泡;N. 細胞核;M. 線粒體;Tn. 單寧物質;VE. 囊泡;ICS. 細胞間隙;PD. 胞間連絲;C. 葉綠體;A. 單寧物質尚未積累;B-C. 細胞質的小液泡中積累單寧;D-E. 細胞質和中央液泡中積累單寧;F. 細胞壁周圍有質體存在圖6 山茱萸幼果期果實中單寧細胞發育CW. Cell wall; V. Vacuole; N. Nucleus; M. Mitochondria; Tn. Tannin substance; VE. Vesicle; ICS. Intercellular space; PD. Plasmodesmata; C. Chloroplast; A. Tannin has not yet accumulated; B-C.Tannin is accumulated in small vacuoles of cytoplasm; D-E. Tannin accumulated in cytoplasm and central vacuole; F. Plastids around the cell wallFig.6 Tannin cell development in C. officinalis fruit at young fruit stage
2.4 單寧物質積累特點
山茱萸果實幼果期早期,中果皮薄壁細胞內尚未觀察到單寧物質積累(圖4,A);超微觀察可見,薄壁細胞細胞壁厚度均勻,細胞質在整個細胞中所占比例較大,細胞核體積較大,核仁大且明顯,細胞內具有線粒體及體積大小不一的液泡 (圖6,A)。單寧物質最早積累在單寧細胞細胞質的小液泡中,小液泡間會發生相互融合,融合后的液泡中單寧物質積累增多(圖6,B-C)。隨著中央大液泡形成,單寧物質主要積累在單寧細胞細胞質的小液泡以及中央大液泡中,囊泡數目較多(圖6,D);單寧細胞進一步發育后,細胞核核膜、核仁消失,細胞壁周圍有質體存在,單寧物質主要積累在中央大液泡中(圖6,E-F)。
中果期時山茱萸果實中單寧細胞體積增大且形態不規則(圖4,E),單寧細胞壁厚度與幼果期時相比有明顯差異。中果期細胞壁厚度不均,細胞壁中段厚度較小,細胞間隙較大,單寧物質積累在體積較大的中央大液泡中(圖7,A)。單寧細胞進一步發育后,細胞壁厚度差異明顯,單寧物質周圍線粒體數目較多(圖7,B)。中果期后期,中央大液泡液泡膜周圍出現一些電子致密物質(圖7,C)。
成熟期單寧細胞壁膨大,中央大液泡液泡膜附近出現電子致密度較高的物質,單寧物質周圍線粒體數目較多(圖7,D)。

CW. 細胞壁;V. 液泡;M. 線粒體;Tn. 單寧物質;ICS. 細胞間隙;PD. 胞間連絲;A-B. 單寧細胞壁變化;C-D. 單寧細胞中電子致密物質圖7 山茱萸中果期(A-C)和成熟期(D)果實中單寧細胞發育CW. Cell wall; V. Vacuole; M. Mitochondria; Tn. Tannin substance; ICS. Intercellular space; PD. Plasmodesmata.; A-B. Changes of tannin cell wall; C-D. Electron dense substances in tannin cellsFig.7 Tannin cell development in C. officinalis fruit at middle fruit stage (A-C) and mature fruit stage (D)
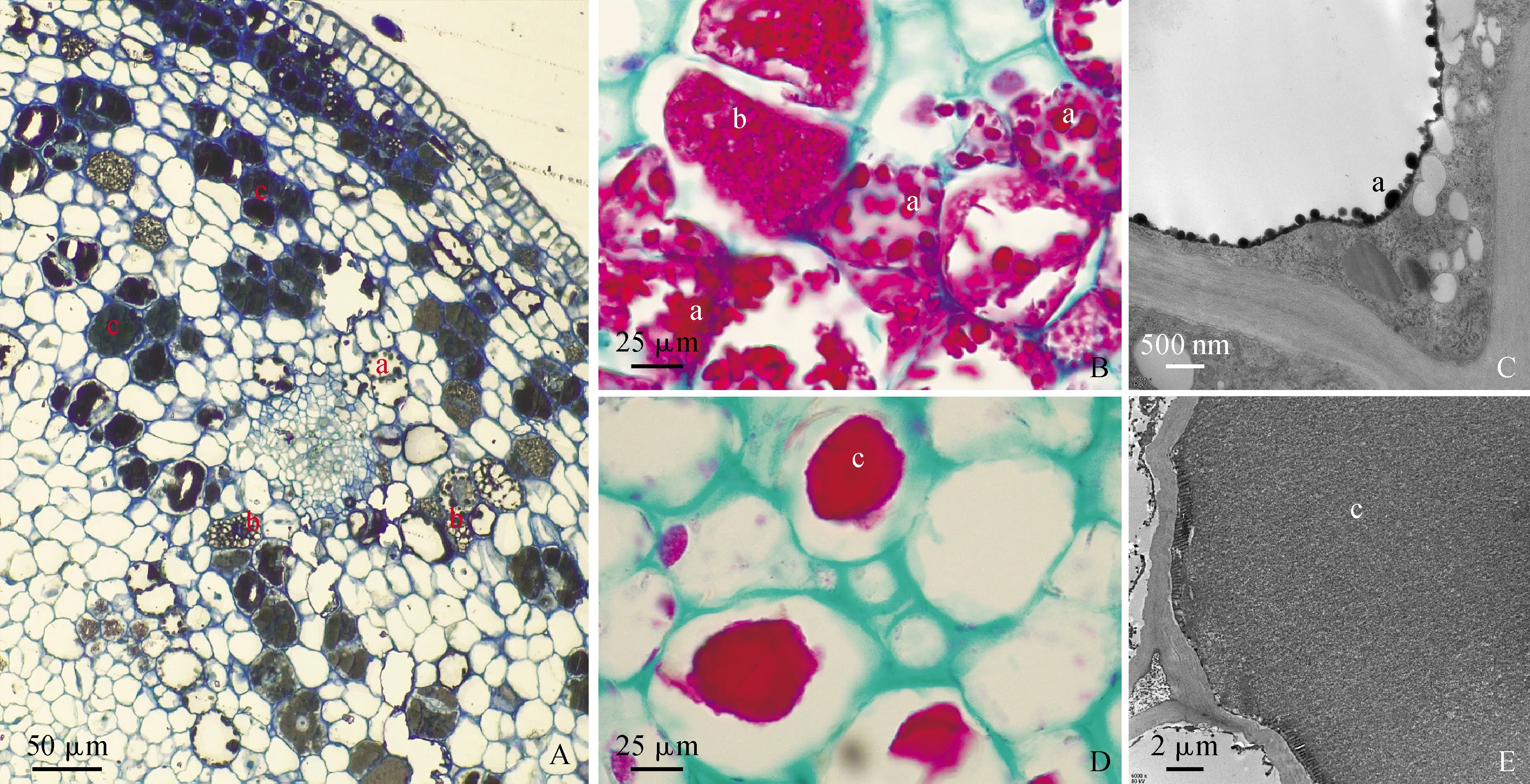
a. 顆粒狀;b. 不規則狀;c. 板塊狀;A. 半薄切片;B、D. 石蠟切片;C、E. 超薄切片圖8 單寧積累的不同形態a. Granular; b. Irregularities; c. Plate shape; A. Semi-thin section; B and D. Paraffin section; C and E. Ultrathin section; Fig.8 Different forms of tannin accumulation
山茱萸果實中單寧物質最早積累在細胞質的小液泡中,隨著單寧細胞進一步發育,單寧物質以顆粒狀(圖8,B-C)、不規則狀(圖8,B)、板塊狀(圖8, D-E)等不同形態積累在中央大液泡中,單寧物質積累的3種不同形態不僅在植物不同發育時期存在,在同一發育時期也會同時存在(圖8,A)。
3 討 論
3.1 單寧物質積累特性與山茱萸果實澀味變化相關
單寧是廣泛存在于植物體內的次生代謝產物[8],在不同植物中分布位置不同。葡萄、香蕉、石榴等果實中單寧主要積累在外果皮中[24],而在山茱萸果實中單寧物質主要積累在由中果皮薄壁細胞特化而來的單寧細胞中,這一研究結果與廖景平對山茱萸果實中單寧物質分布的研究結果相一致[15]。單寧物質經PAS反應顯色,表明含有多糖,單寧細胞中單寧和糖化合的這種形態有利于防止單寧對細胞質產生沉淀作用[15-16]。植物能夠協調自身發育特性以加強對環境的適應性[25-26],Jan Lorie M. Robil等發現隨著寶蓮花的生長發育單寧的含量也會發生變化,寶蓮花中單寧的分布方式與植物為適應環境的防御和繁殖特性相關[27]。山茱萸果實發育過程中單寧細胞數目的變化與單寧含量的測定結果相符,均呈先增后減的變化趨勢。這一變化趨勢可能與山茱萸果實對環境的適應性及果實澀味變化相關。
研究表明,果實澀味強弱受到單寧細胞的體積、數目、形狀及其生長發育進程的影響[28]。苯丙氨酸解氨酶是果實澀味物質合成途徑中的關鍵酶和限速酶,其活性高峰多出現在果實的幼果期和成熟期[28]。而自然脫澀的果實中乙醛和乙醇是單寧的凝固劑,乙醛作為細胞內不需轉運蛋白即可直接跨膜的小分子物質,在細胞膜完整時乙醛可直接跨越液泡膜,使液泡內可溶性單寧轉變為不可溶性單寧,從而降低果實澀味[29]。王玥等對不同品種柿果實的研究發現:柿果實成熟過程中可溶性單寧與水溶性果膠結合是引起柿果脫澀的因素之一[30]。在山茱萸果實幼果期(花后30~60 d)總單寧和可溶性單寧含量增加,單寧細胞數目逐漸增多,這可能與單寧合成過程中相關酶活性較高有關。中果期(花后120 d)時總單寧含量達到最高,中果期早期(花后90~150 d)山茱萸果實中可溶性單寧含量減少,不溶性單寧與可溶性單寧的含量變化呈負相關,可能存在有可溶性單寧向不溶性單寧轉化的趨勢,以降低果實澀味。中果期后期(花后150~180 d)單寧細胞數目減少,總單寧、可溶性單寧和不溶性單寧的含量均降低。隨著果實發育可溶性單寧可能與細胞內的一些物質結合形成復合體,從而使果實澀味降低[29]。成熟期時果實顏色鮮艷,總單寧、可溶性單寧以及不溶性單寧的含量均呈下降趨勢,果實的澀味降低,便于吸引鳥類等潛在的種子散布者對種子進行傳播[31]。
3.2 單寧物質積累的細胞學特性
單寧物質是植物長期進化過程中與環境相互作用的結果,在植物提高自身保護和生存競爭能力、協調與環境關系上充當重要角色[32-33],竹筍中單寧物質被大液泡擠至細胞壁附近的細胞質中,僅有少量進入液泡,積累量少,分布稀疏[34]。而在山茱萸果實中單寧物質早期積累在中果皮單寧細胞細胞質的小液泡中,小液泡間會發生相互融合,單寧物質積累增多,隨著單寧細胞的進一步發育,中央大液泡中積累單寧物質。Magda Andréia Tessmer等對‘金寶’和‘扶余’兩個柿品種中單寧物質的研究結果表明:在柿果實的同一發育時期存在有單寧物質積累的不同形態[35]。山茱萸果實中單寧物質積累的時空特征與柿果實相同,山茱萸果實中單寧物質積累的不同形態不僅在植物不同發育時期存在,在同一發育時期也會同時存在,單寧物質主要以顆粒狀、板塊狀以及不規則狀積累在單寧細胞中。單寧物質屬于類黃酮類次生代謝產物,其合成主要經過莽草酸途徑、苯丙烷途徑和類黃酮途徑最終形成[9]。縮合單寧的合成途徑已成為研究植物次生代謝基因表達及調控的模式途徑。在擬南芥ANS/LDOX突變體tds4中縮合單寧積累量減少,囊泡運輸紊亂,不能形成中央大液泡,縮合單寧前體的合成與運輸與囊泡運輸形成液泡的過程聯系緊密[36]。山茱萸果實的單寧細胞中可見明顯的小液泡和囊泡的存在,這可能與山茱萸果實中單寧前體的合成與運輸相關。單寧細胞發育過程中線粒體數量較多,可見單寧的積累與組織代謝能力加強相對應[37]。基于對不同發育時期山茱萸果實中單寧物質積累特征的研究,后續可結合代謝組及轉錄組等的分析結果以揭示山茱萸果實中單寧物質合成機制,進而為山茱萸果實澀味調控的研究提供理論依據和實驗基礎。
