修剪措施對紫葉紫薇花蕾中內源激素和碳氮營養含量的影響
2022-01-15 05:35:38羅雪夢王曉明曾慧杰李永欣
西北植物學報 2021年11期
羅雪夢,王曉明,曾慧杰,李永欣,唐 麗
(1 中南林業科技大學 林學院,長沙 410004;2 湖南省林業科學院,長沙 410004)
紫薇(LagerstroemiaindicaL.)屬于千屈菜科(Lythraceae)紫薇屬(Lagerstroemia)植物,是中國夏季重要的園林觀賞樹種。大量關于果樹生長的報道中,常采用修剪措施以調整樹枝結構、改變樹體營養分配、增加果實產量等[1-2]。關于紫薇采用修剪措施的報道已有很多,如剪除第一次開花的花穗實現北京地區紫薇花期持續至國慶節前后[3],通過修剪4/5當年生枝與施肥結合實現廈門地區紫薇盛花天數顯著增加[4],達到實現二次開花、花期延長的目的。還有學者研究紫薇一年多次花開的技術措施,如沙飛等[5]研究證明紫薇在夏季采用輪次修剪處理后能夠獲得一年花開四度的最佳狀態。
修剪措施對不同植株不同器官內的生長激素會產生不同程度的影響,通過改變內源激素比例促進或延遲成花進程[6]。蘇曼琳等[1]發現文冠果(XanthocerassorbifoliaBunge)經不同程度短截后不同部位芽內源激素含量有所變化,脫落酸(ABA)含量下降,而赤霉素(GA3)、吲哚乙酸(IAA)和玉米素(ZT)含量均上升。艾沙江·買買提等[7]研究‘富士’蘋果(Malus×domesticaBorkh.)枝條不同方式修剪后芽內源激素平衡,發現ZR和GA3含量顯著提高,促進萌發成枝。修剪后植株各部分之間建立新的營養代謝關系,碳素主要供給生殖生長,氮素主要供給營養生長,如紅花玉蘭(Magnoliawufengensis)不同程度地夏季修剪均能提高可溶性蛋白和可溶性糖含量[8],不同程度短截后薄殼山核桃(CaryaillinoensisK.Koch)新枝葉片及枝條的碳氮比值隨短截程度的增大而提高[9]。紫薇夏剪是目前重要的花期調控措施和生產實踐經驗,目前關于紫薇修剪的報道基本集中在處理后樹勢生長和開花特征等方面,鮮見紫薇修剪后激素水平和營養代謝變化的有關報道。
紫葉紫薇(purple-leafLagerstroemiaindica)枝干直立性強,花色大紅色,成熟葉黑紫色至紫紅色,是極具觀賞價值的選育良種。本研究通過對紫葉紫薇主花枝進行重度修剪,探究修剪措施對花芽內源激素和碳氮代謝的影響,旨在為紫葉紫薇花期調控實踐提供進一步理論依據,以支持紫薇修剪技術的實踐應用與推廣。
1 材料和方法
1.1 試驗地概況
田間試驗地位于湖南省長沙市雨花區湖南省林業科學院林木新品種創制中試基地,地處113°01′E,28°06′N,屬低山丘陵地貌,地勢較平緩,坡度15~20°。屬亞熱帶季風性濕潤氣候,年均氣溫17 ℃,年均降雨量1 400 mm,年均日照時數1 500 h。試驗地土壤為四紀酸性紅壤,pH 4.0~5.5。
1.2 材料與處理
試驗材料為5年生紫葉紫薇品種‘丹紅紫葉’(Lagerstroemiaindica‘Ebony Embers’)。平均樹高約3 m,樹勢中等,長勢良好。選取生長勢一致的‘丹紅紫葉’設2個處理,一是每植株的雙杈花枝的其中一個枝條進行重度修剪(剪去當年生枝長度的2/3),二是以植株不進行修剪為對照(CK)。每個處理3株,單株小區,3次重復。修剪處理于夏季一次花末花期進行,修剪后常規養護管理。
1.3 樣品采集與處理
對照植株于花芽膨大期、花蕾成熟期、初花期、盛花期、末花期各采樣一次,處理植株枝條于修剪后當年二次開花同樣的花發育階段各采樣一次;在修剪處理枝條及對照植株上隨機摘取頂端花蕾,用于內源激素含量測定的樣品液氮速凍后存于-75 ℃冰箱中待測,用于可溶性糖、淀粉、全氮含量測定的樣品在105 ℃烘箱中殺青30 min,80 ℃繼續烘干48 h,粉碎機研磨過80目篩,裝入密封袋待測。
1.4 測定指標及方法
1.4.1 內源激素含量稱取鮮樣0.5 g,液氮研磨為粉末狀,激素的分離純化參照饒丹丹[10]的方法進行,采用高效液相色譜法(HPLC)測定樣品中赤霉素(GA3)、吲哚乙酸(IAA)、脫落酸(ABA)、玉米素核苷(ZR)的含量,高效液相色譜儀為島津LC-20AT,色譜柱為Hypersil BDS C18色譜柱,流動相為甲醇-0.75%冰乙酸水溶液,色譜柱溫度為30 ℃,進樣量為10 μL,檢測波長為254 nm,流速為1.0 mL·min-1。
1.4.2 可溶性糖和淀粉含量稱取樣品粉末0.1 g,采用蒽酮比色法測定樣品中可溶性糖和淀粉的吸光度,再根據葡萄糖標準曲線計算可溶性糖和淀粉含量(mg·g-1)[11],并計算非結構性碳水化合物含量(可溶性糖含量+淀粉含量)。
1.4.3 全氮含量稱取樣品粉末0.2 g,采用硫酸-雙氧水消解,凱氏法測定樣品中全氮含量[12],并計算碳氮比(C/N,非結構性碳水化合物含量/全氮含量)。
1.5 數據分析
采用Excel2010進行數據統計與作圖,SPSS25進行數據分析。
2 結果與分析
2.1 修剪處理對紫葉紫薇花蕾內源激素含量的影響
2.1.1 玉米素核苷(ZR)含量由圖1,①可知,修剪處理紫葉紫薇在開花過程中花蕾的ZR含量均比同期不修剪對照顯著提高,其在花蕾成熟期ZR含量較對照顯著增加了263.98%(P<0.05)。修剪處理組花蕾中ZR含量在花芽膨大期至花蕾成熟期上升,之后逐漸下降,盛花期至末花期再次上升,但整個開花過程ZR含量變化并不顯著(P>0.05)。對照組花蕾中ZR含量隨生育期逐漸下降,從花芽膨大期至初花期變化不顯著,至盛花期開始顯著降低,盛花期和末花期ZR含量分別比初花期顯著降低了57.82%和67.23%,而盛花期和末花期之間無顯著差異(P>0.05)。說明紫葉紫薇經重度修剪后會顯著提高花蕾中ZR水平,也明顯改變了其在花芽發育過程中動態變化規律。
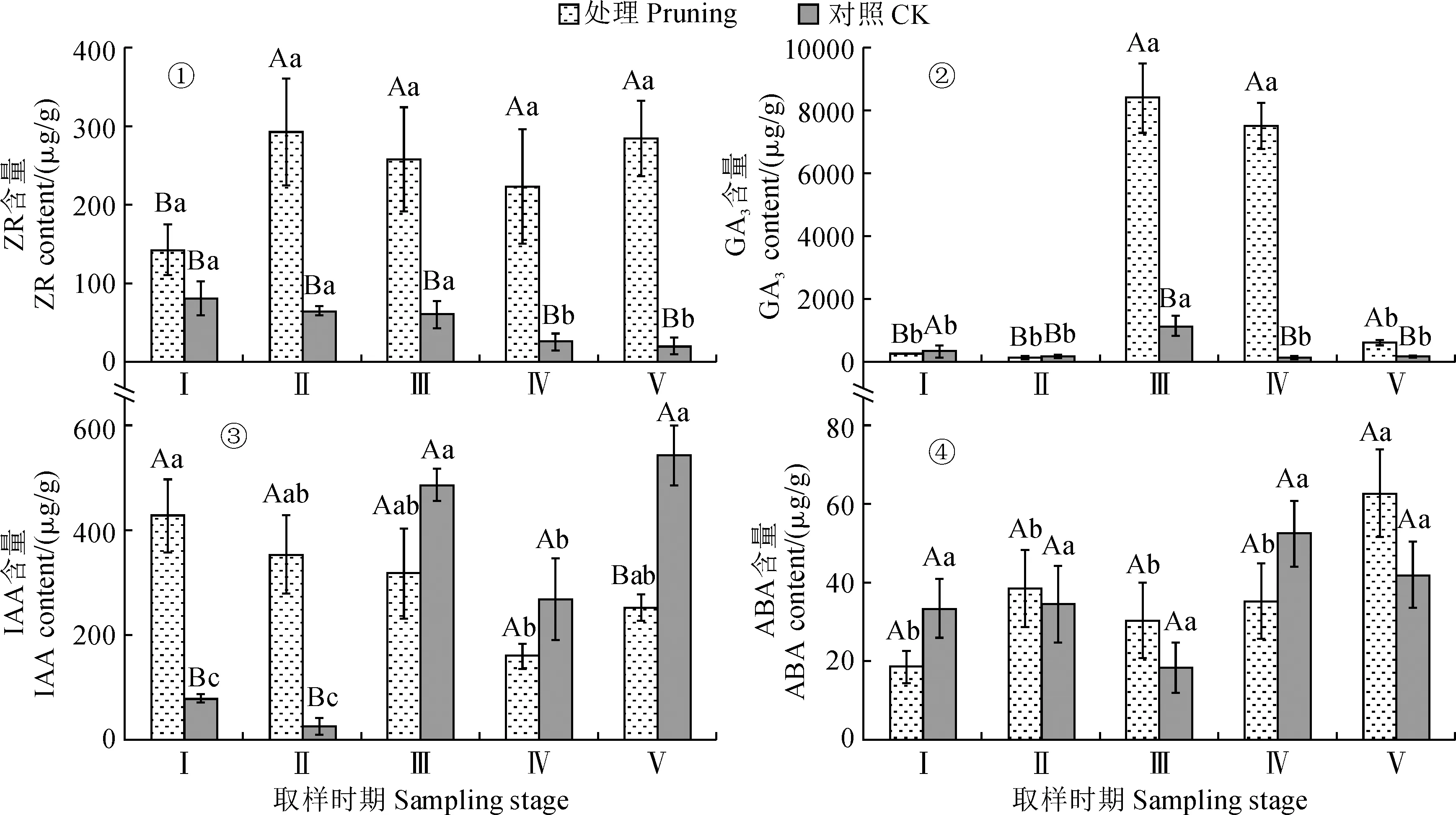
Ⅰ.花芽膨大期;Ⅱ.花蕾成熟期;Ⅲ.初花期;Ⅳ.盛花期;Ⅴ.末花期;處理內不同小寫字母表示不同取樣時期間在0.05水平存在顯著性差異;同一取樣時期內不同大寫字母表示不同處理間在0.05水平存在顯著性差異(P<0.05)。下同圖1 修剪處理對紫葉紫薇花蕾內源激素含量的影響Ⅰ. Flower bud expansion period; Ⅱ. Flower bud maturity period; Ⅲ. Initial flowering period; Ⅳ. Full-flowering period; Ⅴ. Late flowering period;Different lowercase letters indicate a significant difference at the 0.05 level at different sampling time points for the same treatment, and different capital letters indicate a significant difference at the 0.05 level at same sampling time point for the different treatments(P<0.05). The same as belowFig.1 Effect of pruning treatment on endogenous hormone contents in flower buds of purple-leaf Lagerstroemia indica
2.1.2 赤霉素(GA3)含量紫葉紫薇花蕾GA3含量隨開花過程大致呈現“降-升-降”的變化趨勢(圖1,②)。其中,修剪處理的紫葉紫薇花蕾GA3水平從花芽膨大期至花蕾成熟期降低約47%;然后在初花期迅速上升,并在初花期出現GA3含量峰值,此時較花蕾成熟期增加了約60倍;隨后在盛花期GA3含量雖略降低,但仍維持在較高水平;末花期GA3含量比盛花期顯著下降了91.81%(P<0.05)。對照組花蕾中GA3水平從花芽膨大期至花蕾成熟期同樣降低約47%,然后在初花期比前一時期大幅顯著增加約5倍,但初花期至盛花期GA3含量顯著下降了88.62%(P<0.05),盛花期與末花期之間GA3含量變化不顯著(P>0.05)。同時,在紫葉紫薇花芽分化、開花過程中,修剪處理組花蕾中GA3含量在花芽膨大期顯著低于對照組,在初花期、盛花期、末花期GA3含量較對照組顯著升高了2~56倍(P<0.05)。說明紫葉紫薇經重度修剪后會先顯著降低花芽分化前期花蕾中GA3水平,而后顯著提高開花時期花蕾中GA3水平。
2.1.3 吲哚乙酸(IAA)含量圖1,③顯示,修剪處理的紫葉紫薇花蕾IAA含量在花芽分化期顯著高于不修剪對照4.4~12.8倍,其在初花期及盛花期低于對照組但差異不顯著,其在末花期顯著低于對照53.63%(P<0.05)。修剪處理組花蕾中IAA含量在花芽膨大期為最高值,之后在花蕾成熟期及初花期小幅度下降,其在初花期至盛花期顯著降低49.84%(P<0.05),而在末花期比盛花期顯著上升了58.01%。對照組花蕾中IAA含量隨開花過程呈現“降-升-降-升”變化趨勢,其在花蕾成熟期較花芽膨大期降低但未達到顯著性水平,在初花期顯著升高,而后在盛花期顯著下降并處于中等水平,其在末花期再次顯著升高(P<0.05)。說明紫葉紫薇經重度修剪后于花芽分化期提高花蕾中IAA水平,而在開花期降低花蕾中IAA水平。
2.1.4 脫落酸(ABA)含量由圖1,④來看,修剪處理和不修剪對照紫葉紫薇花蕾ABA含量在開花過程中均大致表現為“升-降-升”的動態變化趨勢。其中,修剪處理組花蕾中ABA含量變化從花芽膨大期至盛花期均較穩定,無顯著性差異,其在末花期較盛花期顯著增加78.11%(P<0.05)。對照組花蕾中ABA含量在花芽膨大期和花蕾成熟期小幅波動,其在初花期呈最低水平(18.30 μg·g-1),至盛花期又上升至52.48 μg·g-1,但變化并不顯著(P>0.05)。同時,修剪處理的紫葉紫薇花蕾ABA含量在花芽膨大期低于對照組44.38%,在花蕾成熟期及初花期分別比對照提高10.28%和39.80%,至末花期高于對照組49.55%,但整個開花過程中修剪處理組與對照組之間的ABA含量差異不顯著(P>0.05)。以上結果說明重度修剪對紫葉紫薇花蕾中ABA含量的影響不顯著。
2.2 修剪處理對紫葉紫薇花蕾非結構性碳水化合物含量的影響
2.2.1 可溶性糖含量圖2,①顯示,紫葉紫薇植株修剪處理組花蕾中可溶性糖含量隨花發育進程逐漸升高,且花蕾成熟期及初花期比花芽膨大期分別顯著上升了20.39%、62.48%,其在初花期與盛花期間無顯著差異且處于較高水平(12.3~12.8 mg·g-1),至末花期比盛花期顯著降低38.31%(P<0.05)。紫葉紫薇對照組花蕾中可溶性糖含量隨花發育進程持續上升,各發育時期比花芽膨大期含量顯著上升了23.92%~106.81%(P<0.05)。修剪處理組花蕾中可溶性糖含量在花芽膨大期至盛花期比對照組高6.94%~17.46%,在花蕾成熟期與初花期比對照組顯著增加,其卻在末花期比對照組顯著降低43.06%(P<0.05)。說明紫葉紫薇經重度修剪后花芽分化期及開花前期的花蕾中可溶性糖含量增加,花期末期的花蕾中可溶性糖含量降低。
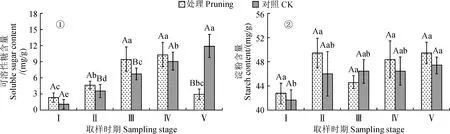
圖2 修剪處理對紫葉紫薇花蕾可溶性糖和淀粉含量的影響Fig.2 Effect of pruning treatment on soluble sugar and starch contents in flower buds of purple-leaf L. indica
2.2.2 淀粉含量從圖2,②可知,紫葉紫薇修剪處理組花蕾中淀粉含量從花芽膨大期至花蕾成熟期上升,在初花期下降,在盛花期與末花期再次逐漸升高;修剪處理組花蕾淀粉含量在花發育進程中處于42.75~49.46 mg·g-1之間,但其在各發育時期間均未表現顯著性差異(P>0.05)。對照組花蕾淀粉含量在花發育進程中呈現逐漸上升趨勢,其從花芽膨大期的41.64 mg·g-1上升到末花期的47.40 mg·g-1,顯著上升了13.83%(P<0.05)。同時,修剪處理花蕾中的淀粉含量在初花期比不修剪對照降低3.96%,在其他時期增加2.66%~7.49%,整體花發育進程中處理組與對照組間差異均并不顯著(P>0.05)。說明紫葉紫薇經重度修剪后花蕾中淀粉含量小幅度增加但受到影響并不顯著。
2.3 修剪處理對紫葉紫薇花蕾全氮含量的影響
修剪處理組和對照組紫葉紫薇花蕾中全氮含量隨著花芽分化和開花時期均逐漸降低(圖3)。其中,修剪處理組花蕾中全氮含量從花芽膨大期至花蕾成熟期顯著降低6.18%(P<0.05),而其在初花期至末花期未表現出顯著性變化。對照組花蕾中全氮含量從花芽膨大期至末花期下降率在1.08%~2.99%,均不表現顯著性差異(P>0.05)。同時,修剪處理組花蕾中全氮含量在花發育各階段均高于對照組,比對照組分別增加了5.76%、1.54%、0.37%、1.91%和1.67%,但兩者間未表現顯著性差異(P>0.05)。說明紫葉紫薇經重度修剪后能夠稍微提高花蕾中全氮含量,但增幅并不顯著。
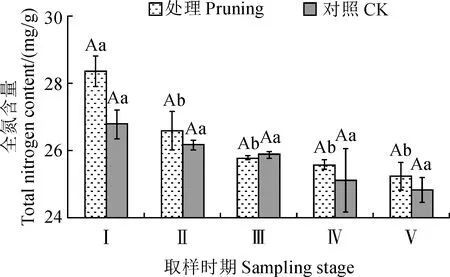
圖3 修剪處理對紫葉紫薇花蕾全氮含量的影響Fig.3 Effect of pruning treatment on total nitrogen content in flower buds of purple-leaf L. indica
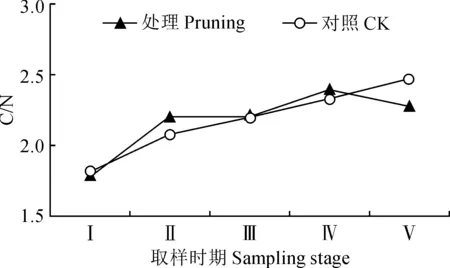
圖4 修剪處理對紫葉紫薇花蕾C/N的影響Fig.4 Effect of pruning treatment on C/N in flower buds of purple-leaf L. indica
2.4 修剪處理對紫葉紫薇花蕾碳氮代謝的影響
修剪處理及未修剪的紫葉紫薇植株的花蕾中碳氮比總體上均呈現上升趨勢(圖4)。其中,修剪處理組的花蕾碳氮比值從花芽膨大期至花蕾成熟期上升較快,初花期比值波動小,然后在盛花期逐漸增大,最終在末花期逐漸下降至2.27。對照組的花蕾碳氮比值從花芽膨大期至末花期持續小幅度上升。同時,修剪處理的紫葉紫薇花蕾中碳氮比值在花芽分化期至盛花期高于不修剪對照,但其間碳氮比值差值小(0.030~0.126);其在末花期低于不修剪對照,碳氮比值相差約0.198。可見,紫葉紫薇經重度修剪會小幅度提高花芽分化期及開花前期花蕾中的碳氮比值。
3 結論與討論
夏季修剪通過剪除部分老枝老葉及殘花敗果,可阻礙夏季高溫多雨引發的葉部疾病,刺激腋芽抽發新枝及新生葉片的營養生長,促進內源激素重新分配及激素平衡調節,增強碳水化合物積累,為花發育進程提供能量。紫薇常采用夏季修剪的方式達到再度開花、延長花期的目的。從花的分化形成到再次開花的整個過程,始終發生著內源激素相互協調、碳水化合物代謝等各種生理生化反應。前人研究發現:巴拉多葡萄(Vitisvinifera‘Balado’)經花穗整穗修剪后測得花穗在盛花期及落花期細胞分裂素(CTK)含量提高,在葡萄幼果期ABA含量顯著升高,盛花期和幼果初期IAA含量降低,盛花期至幼果期GA含量均降低。表明花穗整形可以減少巴拉多葡萄落花落果,增強座果率[13]。本研究發現,紫葉紫薇經過重度修剪后花蕾中ZR含量顯著提高,并且自初花期至末花期GA3含量顯著增加,說明紫葉紫薇修剪后使得植株再次成花、花期延長可能與ZR及GA3含量上升有關。李建軍等[14]也發現忍冬(LonicerajaponicaThunb.)大白期時間延長與花蕾ZR水平升高有關。楊曉紅等[15]表示植物組織處于衰老狀態時,內源細胞分裂素水平會下降以減少水分的消耗,壽命長的花比壽命短的花具有更高的ZR水平。因此高含量ZR可能有利于水分的快速吸收,使植株花期延長。
關于GA誘導開花的理論認為GA濃度越高,表現出越大的促進作用,不存在超最適濃度的抑制作用[16],因此可能修剪處理后的紫葉紫薇花蕾GA3含量在開花期升高促進了植株花期延長。有學者發現短截后從根部運輸而來的養分使枝條剪口下部的芽GA3含量大幅度提高[7],大量研究表明GA可以向上或向下進行雙向運輸[17],本研究中GA3水平的上升可能是因為枝條中上部被剪除后GA通過韌皮部積累于側芽中,根部合成的GA通過木質部不斷輸送,使二次花GA3累積量增加。孟雯雯[18]對高羊茅(FestucaarundinaceaSchreb.)進行不同修剪高度處理后發現其根系赤霉素含量與對照組相比顯著增加。其次,本研究發現,紫葉紫薇經過重度修剪處理后IAA含量在花芽分化期顯著高于對照,在初花期至末花期較對照降低,賈玥等[13]也發現葡萄花穗經整穗修剪后IAA含量在開花中后期顯著低于對照,其他時期IAA含量較穩定。Nito等[19]在報道中認為IAA含量高低在同一花穗中的分布有所不同,枝條上部IAA水平更高,中部及下部IAA水平較低,因此紫葉紫薇開花期IAA含量降低可能是IAA含量更高的部分被剪除而導致。另外,在本研究中,花芽分化期修剪組ABA含量低于對照組但無顯著影響,據黃叢林等[20]研究發現葡萄果實中的ABA大部分來源于葉片,因此葉片可能是紫葉紫薇花蕾中ABA的主要來源,修剪處理使早期花芽內ABA來源不充足,ABA含量低于對照。重度修剪處理對紫葉紫薇花蕾中ABA含量整體情況的影響不顯著,與劉仁道等[21]為促進甜櫻桃(Prunus.aviumL.)生殖生長對直立生長的新梢進行拿枝處理的報道中情況相似,結果發現拿枝后ABA含量與對照差異不顯著。在本研究中,對照組與修剪組在盛花期和末花期ABA含量均處于較高水平,ABA含量的增加可能與ABA具有增強代謝的作用有關,盛花期至末花期紫葉紫薇開始產生果實,需要更多的營養,ABA含量上升促進了代謝庫活性[22]。有學者認為ABA在成花的不同階段可能有不同作用[23],本研究的紫葉紫薇修剪處理組ABA含量在花芽分化期低于對照組而在末花期高于對照組,推測高水平ABA可能與紫葉紫薇植株修剪后花期延長相關,但修剪后ABA含量在末花期升高對紫葉紫薇花期延長的影響效應有待進一步探究。
在花的形成與開放過程中,碳水化合物和氮代謝起著重要的作用。孫穎等[24]認為油茶在簡化整形修剪模式下很大程度地提高了葉碳水化合物的累積,從而提高油茶產量與品質。本研究發現,紫葉紫薇花蕾中可溶性糖含量隨開花進程逐漸上升,但變化量不大,這與高小俊等[25]對芒果(MangiferaindicalL.)短截后花芽內可溶性糖含量變化的研究結果類似。可溶性糖作為關鍵的能量來源和結構物質,其含量的增加可促進細胞液濃度,是花發育進程的保障。修剪處理組可溶性糖含量在花芽分化期高于對照組,可能是修剪處理解除了頂端優勢,可溶性糖大多集中于附近側芽,促進腋芽發育成花芽[26]。前人研究中關于番紅花(CrocussativusL.)[27]、棗(ZiziphusjujubaMill.)[28]等在花芽分化期其可溶性糖含量增加,我們可以認為修剪處理使紫葉紫薇可溶性糖含量提高與紫葉紫薇再次成花相關,高可溶性糖含量有助于紫葉紫薇花芽分化。末花期修剪處理組可溶性糖含量顯著低于對照組,田璐[29]同樣發現忍冬花蕾可溶性糖含量隨花芽分化進行逐漸上升,至花盛放時達最高值,開放后由于能量的大量釋放糖含量開始降低,因此可以認為紫葉紫薇再度開放后末花期有較多果實產生,消耗了植株大部分養分導致糖含量快速下降。本試驗中修剪組紫葉紫薇花蕾中淀粉含量先升高后下降,然后再次升高,與環剝處理下毛棉杜鵑(RhododendronmoulmainenseHook.)花芽分化過程葉淀粉含量變化規律相似[30]。紫葉紫薇初花期淀粉含量的降低趨勢可能是因為淀粉在植株內作為能量補充的角色,并不直接被利用,因此淀粉水解為可溶性糖為生長代謝提供能量,淀粉累積量較低[25]。淀粉作為貯藏性營養物質,在后期成為主要的花發育能量提供者,因此開花中后期淀粉含量繼續升高。
本研究中,紫葉紫薇修剪處理組與對照組的全氮含量表現為逐漸降低趨勢,且修剪組全氮含量略高于對照組,與羅麗[31]短截處理川香核桃(Juglansregia‘Chuanxiang’)全氮含量變化類似。而張翔[9]的研究認為薄殼山核桃在不同程度短截處理后其全氮含量均低于對照,推測這與植物的試驗地區、當地氣候條件及植物品種關系密切。本試驗中紫葉紫薇修剪組的全氮含量較對照組增幅并不顯著,在夏季修剪對紅花玉蘭生理影響的研究中同樣發現全氮及可溶性蛋白含量僅在一定程度上增加[8]。修剪處理使得全氮含量在花發育進程提高的響應特征有待進一步探究。
碳氮代謝是植株最基本的代謝活動,碳和氮的比例適當,營養充足,有利于花芽分化,開花也多。碳氮比協調對紫葉紫薇花發育生理過程起到推動作用。本試驗結果表明,紫葉紫薇花蕾碳氮比值隨花發育時間呈逐漸上升趨勢,修剪組碳氮比值在花芽分化期及開花盛期小幅度高于對照組。碳氮比學說認為高碳氮比狀態有利于花芽分化,反之則抑制花芽的形成。前人對短截處理后的薄殼山核桃的碳氮代謝研究也發現碳氮比提高,從而說明適度修剪處理可以促進薄殼山核桃成花[9]。夏黑葡萄(Vitisvinifera×Vitislabrusca‘Summer Black’)采用噴藥處理促進二花的試驗中,其可溶性糖含量增加,使得碳氮比增加,表明二次成花與碳氮比提高有密切關系[32]。因此,紫葉紫薇經重度修剪后碳氮比值高于對照可能與紫葉紫薇再次成花相關。在本研究中,紫葉紫薇修剪組碳氮比值在末花期下降,且比值低于對照組,原因可能是末花期氣溫降低及光照強度減弱,葉片逐漸衰老,植株光合能力受到影響進而非結構性碳水化合物積累下降;也可能是花果期即將結束,非結構性碳水化合物向其他儲存器官轉移為休眠期做準備[33]。
綜上所述,內源激素的作用方式及碳氮代謝的協調程度與紫葉紫薇整個花發育進程的關系極其復雜,紫葉紫薇經過重度修剪處理后,花蕾中ZR、GA3含量顯著升高,IAA含量先上升再下降,ABA含量變化較穩定;可溶性糖、淀粉、全氮含量增加;碳氮比值小幅度提高,然后在末花期比值降低。多種因素共同調控紫葉紫薇的二次成花進程。本試驗只研究了紫葉紫薇花發育過程中的生理變化(內源激素和碳氮含量),尚未探討內含物質變化的內因,有關影響紫葉紫薇花發育過程中這一系列變化的基因調節機制有待深入研究。另外,本研究為小區域試驗,且品種單一,研究結果具有一定的局限性,還需要增加品種數量及植株數量進一步探究與充實。
