黃龍山林區不同演替時期典型樹種光誘導的氣孔動力學研究
2022-01-15 05:35:40趙霖玉李秧秧
西北植物學報 2021年11期
趙霖玉,李秧秧
(1 西北農林科技大學 林學院, 陜西楊陵 712100;2 黃土高原土壤侵蝕與旱地農業國家重點實驗室, 西北農林科技大學 水土保持研究所, 陜西楊陵 712100)
光是影響氣孔開閉的主要環境因子。自然界中植物由于葉片遮擋、云層覆蓋和太陽傾角的變化,用于光合作用的光量子強度(PPFD)一直在發生變化。在弱光→強光誘導的氣孔開放過程中,氣孔導度(gs)對光強變化的響應通常比光合速率(A)慢一個數量級,從而導致gs直接限制A;而在強光→弱光誘導的氣孔關閉過程中,慢速氣孔關閉導致水分的無效損失,最終影響植物的生產力和水分利用效率[1-3]。因此,加快光誘導過程中gs的響應可能是提高植物生產力和水分利用效率的一條有效途徑。
不同物種[3-5]、同一物種不同基因型[6-9]或同一物種不同生長發育階段[10]光誘導的gs動力學響應過程存在明顯差異。此外,植物生長的環境條件,如土壤水分、光照、大氣CO2濃度和相對濕度等亦影響其光誘導的gs動力學響應過程[7-8,10-11]。目前,關于植物光誘導的gs動力學變化機制并不清楚,一般認為gs響應快慢與氣孔特征(如保衛細胞類型、氣孔密度和大小等)有密切的關系。具有啞鈴型保衛細胞的植物其gs響應速率快于具有腎形保衛細胞的植物[3],斑克木屬植物中具有小氣孔和高氣孔密度的植物種具有更高的gs增加速率,且小氣孔種達到響應幅度50%所需的時間更短[4]。但氣孔特征無法解釋在一系列植物上發現的氣孔關閉速率和氣孔大小無關[5,12]及具有腎形保衛細胞的不同物種間氣孔動力學的差異[3],這表明其他因素,如保衛細胞結構(如細胞骨架和細胞壁彈性)及生物化學因素(如轉運體或離子通道的數量和活性等)等可能在光誘導的gs動力學反應中起著十分重要的作用。
光和水分是影響黃土高原地區植被演替的主要因素,研究表明,植被演替早期種一般具有高的光合速率和水分利用效率(WUE)[13-15],盡管也有相反的報道[16]。這種高光合速率和高WUE有利于演替早期種進行快速生長及適應演替早期開放的光環境。演替早期種高的WUE是否與其快速的光誘導氣孔動力學有關尚不清楚。對熱帶雨林2種演替早期種和9種演替后期種光誘導氣孔動力學的研究表明,2種演替早期樹種的氣孔開放速率和T50%(達到穩態氣孔導度50%所需時間)分別為0.06~0.26 mmol·m-2·s-2和5~14 min,而9種演替后期種的氣孔開放速率和T50%分別為0.06~0.12 mmol·m-2·s-2和5~17 min,不同演替位樹種的氣孔開放速率和T50%存在重疊,并未因演替位的不同而存在差異[17]。在受水分限制的黃土高原地區植被演替中演替早期種和后期種的氣孔動力學是否亦如此尚不清楚。為此,本研究對黃土高原黃龍山林區3種不同演替階段樹種幼苗光誘導的氣孔動力學進行了探討,試圖回答以下問題:1)不同演替階段樹種光誘導的氣孔動力學參數是否存在差異? 2)這種動力學參數差異是否與其氣孔特征有關? 3)不同樹種光誘導的氣孔動力學差異是否可以解釋其水分利用效率的差異? 對這些問題的回答,有助于闡釋黃土高原地區不同演替階段樹種水分利用效率差異的生理基礎,從而為半干旱區樹種選擇和水分調控提供一定的理論依據。
1 材料和方法
1.1 試驗材料
試驗用樹種為黃龍山林區演替早期種山楊(Populusdavidiana)和白樺(Betulaplatyphylla)以及演替后期種遼東櫟(Quercusliaotungensis)的實生苗,采用盆栽試驗,在陜西楊陵西北農林科技大學水土保持研究所盆栽試驗場進行。2021年3月下旬在陜西黃龍縣蔡家川林場挖取3種樹種的實生苗(1~3年生),每種樹種大約15株,帶回到西北農林科技大學水土保持研究所。挑選生長均一的實生苗栽植于塑料盆中,試驗用土為蔡家川林場林地表層土壤,其土壤容重、有機質、全N、堿解N和Olsen-P含量分別為1.26 g·cm-3、12.56 g·kg-1、1.05 g·kg-1、78.26 mg·kg-1和30.5 mg·kg-1。土壤經風干、粉碎過篩后裝入塑料盆(上底內徑28.5 cm、下底內徑21.0 cm、高24.5 cm),每盆裝土14 kg,栽植樹苗1株,每個樹種栽植10盆。3種樹種幼苗于2021年3月25日栽種,然后澆水至田間最大持水量。山楊和白樺4月初開始萌發生長,遼東櫟4月中旬開始萌發生長。在生長過程中,土壤含水量一直維持在田間持水量的75%~85%,每隔2周,用Hoagland營養液進行養分補充,每次補充500 mL完全Hoagland營養液。于2021年6月20日(生長約2個多月后)每個樹種挑選無病蟲害的健康株8株,對冠層上部完全展開的成熟葉進行氣孔導度動力學、氣孔特征、光合能力和碳同位素分辨率(δ13C)等的測定。
1.2 測定項目及方法
1.2.1 氣孔導度動力學參數使用LI-6400便攜式光合作用系統(Li-COR, Lincoln, USA)測定最上部完全展開葉的氣孔導度動力學參數。植物在室內恒定環境下提前適應一晚,整個測量過程按照McAusland等[3]的光誘導方法,涉及從弱光→強光誘導的氣孔開放過程和從強光→弱光誘導的氣孔關閉過程。即葉片先在100 μmol·m-2·s-1PPFD下平衡至A和gs達到穩定狀態(一般持續0.5 h),然后將光強增加到1 200 μmol·m-2·s-1持續測定1 h后,再返回至100 μmol·m-2·s-1直至新的穩定狀態(0.5 h左右)。測定時葉室條件設置為:葉溫25 ℃,CO2濃度400 μmol·mol-1,相對濕度60%。整個測定過程在生長箱中進行,使用自動打點每隔30 s記錄一次數據。考慮到植株晝夜生理節律的影響,所有測量均在8:00~15:00之間進行。每個處理重復4次。
1.2.2 氣孔解剖特征測量完葉片氣體交換參數后的葉片用來進行氣孔解剖特征的測定,采用指甲油印跡法制成臨時裝片保存備用,每一片葉下表面2個裝片。用顯微數碼相機進行圖片拍攝,每個裝片觀察4個視野,用Image J軟件測量;氣孔密度(SD)和表皮細胞密度的觀測在100×鏡下進行,氣孔長(保衛細胞長度,SL)、氣孔寬(垂直于氣孔長的保衛細胞最大寬度,SW)和氣孔面積(SA)的觀測在400×鏡下進行,每片葉下表面氣孔形態特征的觀測不少于60個氣孔。依據測定數據計算氣孔指數[SI,氣孔密度/(氣孔密度+表皮細胞密度)×100%][7],表征氣孔數占葉表面細胞總數的百分比。
1.2.3 葉片光合能力及δ13C值在剩余4株中與葉氣體動力學參數測定相同位置選擇葉片,進行CO2響應曲線測定。CO2由LI-6400光合儀攜帶的小氣瓶提供,其設置梯度為400、300、200、100、50、200、400、600、800、1 000、1 200、1 500和1 800 mol·mol-1,測定前先在飽和光強下誘導30 min,測定過程中維持光強1 200 μmol·m-2·s-1,葉溫大約25 ℃,相對濕度50%~60%。利用Sharkey等的方法[18]求得最大羧化速率(Vcmax)和最大電子傳遞速率(Jmax),連同大氣CO2濃度下(400 μmol·mol-1)的光合速率(Aair)來表征植物葉片的光合能力。測定結束后采集葉片掃描測量葉面積,然后在75 ℃下烘干至恒重,稱其干重,葉干重與葉面積之比即為比葉重(LMA)。烘干樣粉碎過篩后,用MAT-251型質譜儀(美國Finnigan公司)測定葉片的13C/12C比,并以PDB(Pee Dee Belemnite)為標準計算葉片δ13C,質譜儀分析精度為±0.2‰。所有測定重復4次。質譜儀測定的葉片δ13C可間接反映C3植物水分利用效率的大小,該值越大,葉長期水分利用效率越大。
1.3 數據分析
1.3.1 氣孔導度動力學模型及參數采用常用的指數曲線對氣孔的開放與關閉過程進行模擬[18]:
氣孔開放過程:gs=gsmax+(gsmin-gsmax)e-t/Ki
(1)
氣孔關閉過程:gs=gsmin+(gsmax-gsmin)e-t/Kd
(2)
其中:gs為擬合的氣孔導度,gsmin與gsmax分別為最小和最大穩態gs值,Ki和Kd分別為氣孔開放和氣孔關閉過程中的時間常數(即達到gs變異幅度63%所需時間),gsmax-gsmin為氣孔開放和氣孔關閉過程中gs的變化幅度(Δgsi和Δgsd),e為歐拉常數。用Vialet-Chabrand等提供的Excel菜單[19]對氣孔動力學過程進行模擬。
1.3.2 統計分析用SPSS 20.0軟件進行統計分析。處理之間gs動力學參數、氣孔解剖結構及葉光合能力、葉δ13C值之間的差異顯著性用單因素方差分析,若單因素方差分析達到顯著后,用Tukey法進行處理之間差異顯著性的多重比較(α=0.05)。
2 結果與分析
2.1 不同演替時期樹種光誘導的氣孔動力學
圖1顯示,3種樹種光誘導的氣孔導度動力學曲線變化趨勢較相似;氣孔開放過程中山楊和白樺氣孔導度增加速率明顯超過遼東櫟,且達到最大值后存在一定程度的下降,而遼東櫟這種過飽和現象較弱;遼東櫟氣孔關閉最先達到穩態,其次是白樺,而山楊最晚達到穩態。
進一步對圖1氣孔導度動力學過程進行模擬獲得相應參數(圖2)。其中,在弱光→強光誘導的氣孔開放過程中,3個樹種的時間常數(Ki)和氣孔導度響應幅度(Δgsi)均存在顯著差異,具體表現為:山楊和白樺的Ki顯著低于遼東櫟,但Δgsi顯著大于遼東櫟,而山楊和白樺間的Ki及Δgsi均無顯著差異。在強光→弱光誘導的氣孔關閉過程中,山楊和白樺氣孔關閉的時間常數(Kd)均顯著大于遼東櫟,且山楊和白樺間無顯著差異;山楊的氣孔導度響應幅度(Δgsd)顯著大于白樺和遼東櫟,后兩者無顯著差異。山楊和白樺的Kd/Ki均接近1,并顯著高于遼東櫟,表明它們的氣孔開放與關閉的時間相當,呈現出近似對稱的響應;而遼東櫟Kd/Ki遠小于1,表明氣孔關閉時間遠快于氣孔開放時間,呈現出明顯的不對稱響應。
2.2 不同演替時期樹種的葉氣孔解剖特征
3種樹種葉片均只有下表面具有氣孔。從表1可知,山楊和白樺的氣孔密度顯著低于遼東櫟,兩者的氣孔密度僅分別為遼東櫟的19.7%和32.7%;3種樹種氣孔長表現為山楊>白樺>遼東櫟,且三者之間均存在顯著差異;3種樹種氣孔寬度和氣孔面積均表現為山楊和白樺顯著大于遼東櫟,且山楊和白樺之間無顯著差異;3種樹種氣孔指數則表現為山楊<白樺<遼東櫟,且三者之間均存在顯著差異。以上結果說明演替后期種遼東櫟表現出氣孔密度大、氣孔小和氣孔指數大的特點。
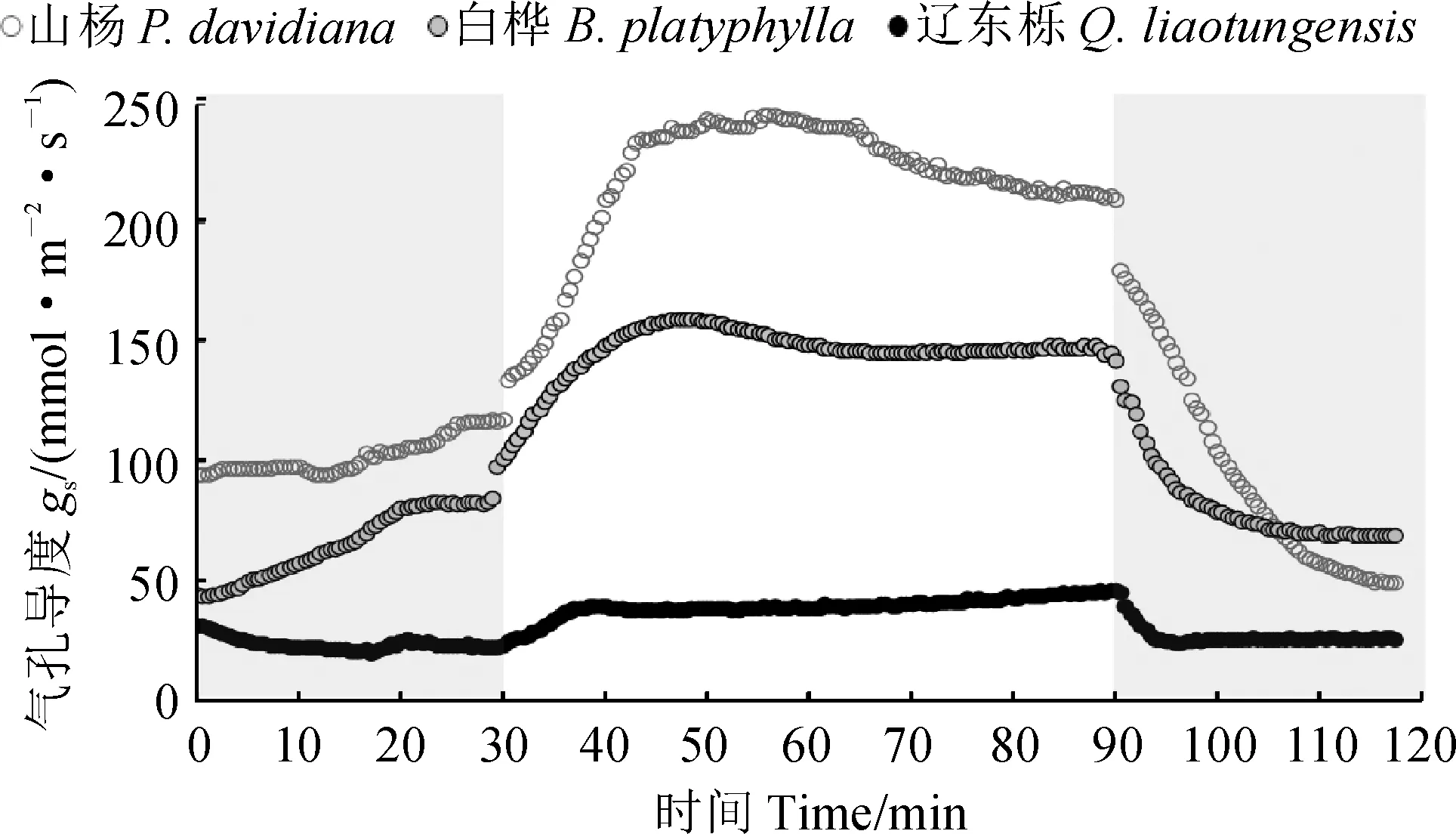
灰色部分為100 mol·m-2·s-1的弱光,白色部分為1 200 mol·m-2·s-1的強光圖1 3種樹種光誘導的氣孔導度動力學曲線Grey and white color correspond to 100 and 1 200 mol·m-2·s-1 PPFD, respectivelyFig.1 Light-induced stomatal conductance dynamics in three typical species
2.3 不同演替時期樹種的葉光合能力與長期水分利用效率
表2顯示,3種樹種的葉面積并無顯著差異;遼東櫟的比葉質量最高并顯著高于白樺,但與山楊之間無顯著差異,而山楊的比葉質量與白樺之間也無顯著差異。演替早期種山楊和白樺的光合速率、最大羧化效率和最大電子傳遞速率顯著高于演替后期種遼東櫟,且山楊的最大光合速率與最大電子傳遞速率顯著高于白樺。3種樹種的葉δ13C值之間均存在顯著性差異,表現為山楊>白樺>遼東櫟。以上結果提示相對于演替后期種遼東櫟,演替早期種山楊和白樺具有更高的光合能力和長期水分利用效率。

不同小寫字母代表處理間在0.05水平下差異顯著,下同圖2 3種樹種氣孔開放和氣孔關閉過程中光誘導的氣孔導度動力學參數The different normal letters indicate significant difference among treatments at 0.05 level. The same as below.Fig.2 Dynamic parameters of stomatal conductance during light-induced stomatal opening and closing in three species

表1 3種樹種葉的氣孔解剖特征

表2 3種樹種葉光合能力與δ13C值
3 討 論
3.1 不同演替時期樹種光誘導的氣孔導度動力學參數間差異
本研究結果表明:在光誘導的氣孔開放過程中,演替早期種山楊和白樺氣孔開放明顯快于演替后期種遼東櫟,而氣孔關閉過程則相反,演替后期種遼東櫟的氣孔關閉更快。一般認為,氣孔的快速開放有助于葉快速達到最大光合速率,從而減輕氣孔開放過程中氣孔導度對光合作用的限制,而氣孔關閉過程中快速的氣孔關閉則有助于保持水分,從而減輕碳攝取降低時水分的無效消耗[3,6,10]。本研究中,演替早期種山楊和白樺一般生長在山坡底部,土壤水分條件較好且具有高的光合速率,因而快速的氣孔開放反應有助于減輕氣孔開放過程中的氣孔限制,同時一定程度上補償了演替早期種高蒸騰導致的水力梯度增大的缺陷[4],反映了演替早期種對演替早期變化的光環境的適應性。演替后期種遼東櫟一般生長在水分限制的山坡上,因而快速的氣孔關閉有助于維持水分。
植物氣孔開放和氣孔關閉的速率不同,一些植物氣孔關閉快于氣孔開放,如在McAusland等研究[3]的15種植物中,有7種的Kd顯著小于Ki,在禾本科植物水稻[6]和小麥[10]也發現氣孔關閉比氣孔開放快,這種不對稱性可能與這些植物水分保持優先于碳同化的策略有關。演替早期種山楊和白樺Kd/Ki值接近于1,氣孔開放與氣孔關閉快慢相近,可能與其生境中水分相對充足,植物優先碳同化來維持高生長速率有關,遼東櫟氣孔低的Kd/Ki值表明其在受水分限制的植被演替過程中,選擇了保水優先于CO2攝取的策略,這種策略有助于增加其對長期水分虧缺的適應。
3.2 不同演替時期樹種氣孔動力學參數和氣孔解剖特征之間的關系
由于研究的物種少(n=3),我們無法建立物種間氣孔動力學參數與氣孔解剖特征間的相關關系。前人研究表明:氣孔密度大、氣孔小及氣孔指數高的物種其氣孔反應更快[4,17]。從本研究看,演替后期種遼東櫟氣孔密度大、氣孔小且氣孔指數高,但其氣孔開放最慢。其次,山楊和白樺的氣孔長和氣孔指數也具有一定差異,但二者氣孔開放和氣孔關閉快慢并無顯著差異。因而我們認為,氣孔特征之間的差異可能不是影響3種物種間氣孔動力學快慢的主要因素,其他因素如副衛細胞數量和大小、葉生物化學、離子通道相關基因表達等可能影響更大[2,20]。
3.3 不同演替時期樹種氣孔動力學參數與水分利用效率間關系
氣孔動力學研究顯示:演替早期種山楊和白樺的氣孔開放快于遼東櫟,而氣孔關閉過程中遼東櫟的氣孔關閉更快。演替早期種山楊和白樺同時具有更高的光合能力。快速的氣孔開放有助于植物利用光斑,從而最大化光合作用[21],但在小麥上的研究表明穩態光合能力與氣孔開放和關閉的時間常數呈正相關,即穩態光合能力越強,氣孔開放越慢,穩態光合能力與氣孔開閉快慢之間存在一種妥協關系[10]。本研究中演替早期種山楊和白樺快速的氣孔開放可能有助于其取得更高的光合能力,但其慢速的氣孔關閉則增加了氣孔關閉過程中的蒸騰耗水,不利于水分利用效率的提高。葉片長期水分利用效率與氣孔開放與關閉過程中光合速率和蒸騰速率變化均有關,盡管山楊和白樺氣孔關閉過程中失水增加,但氣孔開放過程中光合增加,導致其具有更高的水分利用效率。一些研究認為:快速的氣孔關閉有助于保持水分,從而提高葉器官水平水分利用效率[3,6,10],但從本研究看,遼東櫟快速的氣孔關閉反應并未增加其水分利用效率,相反其長期水分利用效率低于山楊和白樺,原因可能與遼東櫟慢速的氣孔開放限制了其光合速率有關。
植物葉長期水分利用效率不僅與動態光強下氣孔的反應有關,與穩態光下氣孔的反應亦有關。在火炬松 (Pinustaeda)上的研究表明:動態光下慢速的氣孔導度響應對總的日蒸騰速率、碳同化及水分利用效率影響不大[22],說明穩態光下氣孔的反應可能更重要。此外,葉水分利用效率也受氣孔對其他因素,如土壤水分、溫度、蒸汽壓虧缺和風等敏感性的影響。因此,今后需要通過氣孔對環境因子的綜合響應研究,以揭示3種植物氣孔調節和葉水分利用效率差異的生理機制。
