外源神經(jīng)遞質(zhì)5-羥色胺(5-HT)和多巴胺(DA)調(diào)控三疣梭子蟹爭(zhēng)斗行為的初步研究*
2022-01-19 06:00:54母昌考徐善良王丹麗
海洋與湖沼 2022年1期
關(guān)鍵詞:神經(jīng)遞質(zhì)
吳 波 張 順 張 成 母昌考 徐善良, 2① 王丹麗①
外源神經(jīng)遞質(zhì)5-羥色胺(5-HT)和多巴胺(DA)調(diào)控三疣梭子蟹爭(zhēng)斗行為的初步研究*
吳 波1張 順1張 成1母昌考1徐善良1, 2①王丹麗1①
(1. 寧波大學(xué)海洋學(xué)院 浙江寧波 315211; 2. 海洋生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室 浙江寧波 315211)
影響甲殼類(lèi)動(dòng)物爭(zhēng)斗行為的因素包括性別差異、體型和生殖狀態(tài)等, 神經(jīng)遞質(zhì)對(duì)甲殼動(dòng)物的爭(zhēng)斗行為也具有調(diào)控作用。為探索三疣梭子蟹在養(yǎng)殖和繁殖過(guò)程中由于爭(zhēng)斗行為受損或死亡的行為機(jī)制, 采用雙抗體夾心法測(cè)定爭(zhēng)斗前后三疣梭子蟹血淋巴中5-HT和DA的含量, 并分析了爭(zhēng)斗前后肌肉、胸神經(jīng)節(jié)、肝胰腺三種組織中5-HT和DA受體基因表達(dá)量變化。同時(shí), 通過(guò)注射外源5-HT和DA, 記錄并分析了三疣梭子蟹爭(zhēng)斗行為差異以及三種組織中5-HT和DA受體基因表達(dá)量變化。結(jié)果表明: (1) 爭(zhēng)斗后三疣梭子蟹血淋巴中5-HT含量顯著升高, DA含量顯著降低(<0.05); (2) 注射10–6mol 5-HT能顯著減少三疣梭子蟹的爭(zhēng)斗行為發(fā)生次數(shù)以及爭(zhēng)斗持續(xù)時(shí)間(<0.05), 而注射10–6mol DA卻能夠顯著增加三疣梭子蟹的爭(zhēng)斗行為發(fā)生次數(shù)和爭(zhēng)斗持續(xù)時(shí)間(<0.05); (3) 爭(zhēng)斗前后5-HT和DA受體在肌肉和胸神經(jīng)節(jié)中表達(dá)量發(fā)生顯著變化(<0.05), 且不同受體在相同組織中未呈現(xiàn)相同的變化趨勢(shì); (4) 注射外源DA能夠調(diào)節(jié)、、受體的表達(dá)水平(<0.05), 外源5-HT也能調(diào)節(jié)和受體的表達(dá)水平(<0.05)。綜上所述, 外源5-HT對(duì)三疣梭子蟹爭(zhēng)斗行為起抑制作用, 而DA對(duì)三疣梭子蟹爭(zhēng)斗行為起促進(jìn)作用。同時(shí), 外源5-HT和DA也會(huì)引起相關(guān)受體表達(dá)量變化, 且它們之間存在相互作用。
三疣梭子蟹; 爭(zhēng)斗行為; 5-HT/DA;受體
“爭(zhēng)斗行為”指的是同種動(dòng)物個(gè)體間為獲得食物、配偶或庇護(hù)所等資源在相遇時(shí)發(fā)生爭(zhēng)斗的行為, 是一種典型的社會(huì)行為(李玉全等, 2013)。甲殼動(dòng)物由于具有強(qiáng)大的螯足, 種群內(nèi)爭(zhēng)斗行為往往表現(xiàn)得尤為突出, 嚴(yán)重影響其養(yǎng)殖過(guò)程中的存活率(李藝等, 2019)。而影響甲殼類(lèi)動(dòng)物爭(zhēng)斗行為的因素有很多, 包括性別差異(Tina, 2018)、體型(Hamasaki, 2018)和生殖狀態(tài)(Costa, 2016)等。此外, 有研究表明神經(jīng)遞質(zhì)對(duì)甲殼動(dòng)物的爭(zhēng)斗行為也具有調(diào)控作用(Zhu, 2018; Yang, 2019)。且爭(zhēng)斗能力不同的甲殼動(dòng)物體內(nèi)神經(jīng)遞質(zhì)含量也是不同的, 而神經(jīng)遞質(zhì)含量的高低變化也會(huì)影響到甲殼動(dòng)物個(gè)體的爭(zhēng)斗能力(Huber, 2001; 趙玉超等, 2017)。而5-羥色胺(5-HT)和多巴胺(DA)作為兩種重要的神經(jīng)遞質(zhì), 與甲殼動(dòng)物爭(zhēng)斗行為的調(diào)控密切相關(guān)(王芳等, 2020)。但是, 目前有關(guān)于5-HT和DA對(duì)三疣梭子蟹()爭(zhēng)斗行為影響的研究較少。
5-羥色胺(5-HT)也稱(chēng)血清素(serotonin), 是一種廣泛分布在脊椎動(dòng)物和無(wú)脊椎動(dòng)物體內(nèi)的抑制性神經(jīng)遞質(zhì), 在攝食、睡眠和攻擊性等調(diào)控過(guò)程中起著關(guān)鍵作用(Serretti, 2006)。同時(shí), 5-HT也被認(rèn)為是影響爭(zhēng)斗行為的最主要的神經(jīng)遞質(zhì)(Bortolato, 2013), 其他神經(jīng)遞質(zhì)可能是先通過(guò)影響5-HT的含量變化來(lái)影響爭(zhēng)斗行為(孔雀等, 2006)。趙玉超等(2017)研究發(fā)現(xiàn)通過(guò)注射外源5-HT能夠有效降低日本囊對(duì)蝦()、凡納濱對(duì)蝦()和中國(guó)明對(duì)蝦()平均爭(zhēng)斗次數(shù)。但也有研究表明, 5-HT可能不是直接對(duì)爭(zhēng)斗行為起作用, 而是對(duì)大腦中樞的行為反應(yīng)進(jìn)行調(diào)節(jié), 進(jìn)而影響動(dòng)物的爭(zhēng)斗行為(Bacqué-Cazenave, 2018)。多巴胺(DA)是一種廣泛分布于甲殼類(lèi)動(dòng)物體內(nèi)的兒茶酚胺類(lèi)神經(jīng)遞質(zhì), 也能夠參與甲殼動(dòng)物的運(yùn)動(dòng)能力和爭(zhēng)斗行為的調(diào)節(jié)(Zhu, 2018)。此外, 5-HT和DA需要與突觸前膜、突觸后膜的多種受體進(jìn)行結(jié)合才能發(fā)揮作用(段云峰等, 2013)。目前已經(jīng)在蝦蟹中發(fā)現(xiàn)存在多種5-HT和DA受體, 這些受體大部分屬于G蛋白偶聯(lián)受體家族(Northcutt, 2016)。它們可以通過(guò)激活第二信使來(lái)調(diào)節(jié)5-HT和DA的釋放, 進(jìn)而影響甲殼動(dòng)物的爭(zhēng)斗行為(Lee, 2008; Rodgers, 2013)。
本研究以三疣梭子蟹為實(shí)驗(yàn)對(duì)象, 探究5-HT、DA與爭(zhēng)斗行為間的關(guān)系。旨在了解5-HT和DA對(duì)三疣梭子蟹爭(zhēng)斗行為的響應(yīng)機(jī)制, 為進(jìn)一步了解三疣梭子蟹爭(zhēng)斗行為的分子機(jī)制和減少其養(yǎng)殖過(guò)程中的爭(zhēng)斗傷亡提供一些理論依據(jù)。
1 材料與方法
1.1 實(shí)驗(yàn)三疣梭子蟹暫養(yǎng)
實(shí)驗(yàn)所用三疣梭子蟹()從寧波鄞州區(qū)瞻岐鎮(zhèn)一養(yǎng)殖場(chǎng)獲得, 挑選規(guī)格基本一致、體重為(90±10) g的肢體完整的健康雌雄個(gè)體, 置于水槽(85 cm×40 cm×25 cm)中單養(yǎng)。實(shí)驗(yàn)開(kāi)始前在實(shí)驗(yàn)環(huán)境下適應(yīng)一周, 以減少應(yīng)激。所用海水為經(jīng)沉淀、沙濾后的自然海水, 鹽度為25.5±1.0, 水溫為(26.0±1.0) °C, pH 7.7±0.3。每天18:00投喂縊蟶, 次日7:00清理殘餌及糞便, 日換水1/2, 爭(zhēng)斗當(dāng)天停止投喂。
1.2 實(shí)驗(yàn)設(shè)計(jì)
采用“雄×雄”配對(duì)爭(zhēng)斗, 分為注射5-HT與注射DA的實(shí)驗(yàn)組和對(duì)照組。實(shí)驗(yàn)組三疣梭子蟹5-HT/DA的注射量分別為10–6、10–8和10–10mol/只。另設(shè)三個(gè)對(duì)照組, 即注射等量0.85%無(wú)菌生理鹽水(生理鹽水組)、不注射的空白對(duì)照組以及規(guī)格一致的未爭(zhēng)斗組, 每組9個(gè)重復(fù)。配對(duì)的兩只蟹規(guī)格盡量一致, 分別在頭胸甲上做白色標(biāo)記。如圖1所示, 實(shí)驗(yàn)開(kāi)始啟動(dòng)攝像頭, 先將兩只蟹分別置于實(shí)驗(yàn)水槽兩側(cè), 槽中間用不透明隔板隔開(kāi), 適應(yīng)10 min后撤掉隔板, 記錄兩個(gè)體之間的爭(zhēng)斗行為全過(guò)程, 實(shí)驗(yàn)拍攝時(shí)長(zhǎng)為90 min, 超過(guò)實(shí)際爭(zhēng)斗持續(xù)時(shí)間, 保證能夠記錄完整的爭(zhēng)斗過(guò)程。
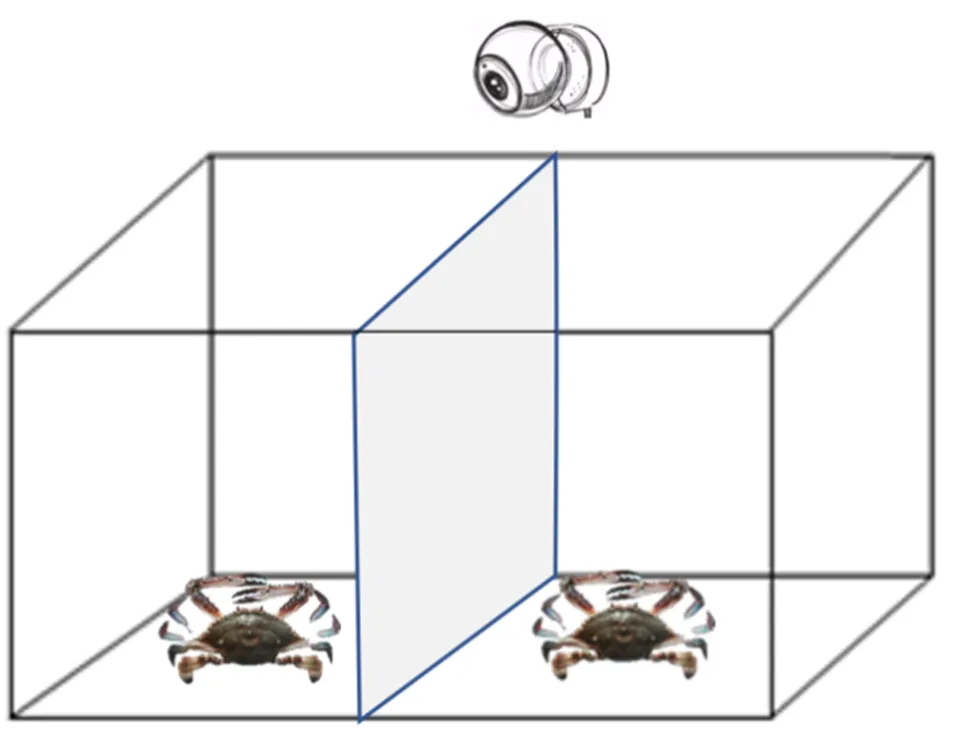
圖1 三疣梭子蟹爭(zhēng)斗實(shí)驗(yàn)裝置圖
注: 中間為不透明PVC板
1.3 5-HT和DA注射試劑準(zhǔn)備
抗凝劑的配制: 應(yīng)根據(jù)標(biāo)本的要求選擇EDTA或檸檬酸鈉作為抗凝劑。
注射液的配制及劑量: 注射前, 將鹽酸5-HT溶于0.85%無(wú)菌生理鹽水配成濃度為5×10–3、5×10–5以及5×10–7mol/L的5-HT溶液, 同理配制濃度為5×10–3、5×10–5和5×10–7mol/L的DA溶液。注釋劑量及方法根據(jù)前人的研究確定(Zhu, 2018; Yang, 2019)。
1.4 注射實(shí)驗(yàn)
兩個(gè)實(shí)驗(yàn)組分別在蟹游泳足的關(guān)節(jié)膜注射0.2 mL 5-HT溶液或DA溶液。5-HT注射量為10–6、10–8和10–10mol/只, DA注射量為10–6、10–8和10–10mol/只, 生理鹽水組則注射等量生理鹽水。有研究表明, 外源5-HT注射后能夠迅速分布整個(gè)血淋巴, 大部分在5min內(nèi)能夠被組織吸收(Tierney, 2001)。注射后的兩只蟹同樣分別置于實(shí)驗(yàn)水槽兩側(cè), 槽中間用不透明隔板隔開(kāi), 適應(yīng)10 min后撤掉隔板。同時(shí), 參考Zhu等(2018)的研究, 外源DA的效果能夠持續(xù)2 h左右, 滿(mǎn)足實(shí)驗(yàn)需要。
1.5 視頻分析與數(shù)據(jù)采集
通過(guò)紅外攝像與視頻逐幀回放技術(shù), 記錄并統(tǒng)計(jì)各組蟹的爭(zhēng)斗行為發(fā)生次數(shù)、持續(xù)時(shí)間, 對(duì)爭(zhēng)斗行為進(jìn)行量化分析。當(dāng)一只螃蟹朝向另一只螃蟹靠近或兩只蟹同時(shí)向?qū)Ψ揭苿?dòng)視為爭(zhēng)斗開(kāi)始, 當(dāng)爭(zhēng)斗中兩只蟹超過(guò)5 min未互動(dòng)視為爭(zhēng)斗結(jié)束, 從爭(zhēng)斗開(kāi)始到爭(zhēng)斗結(jié)束的時(shí)間即為爭(zhēng)斗持續(xù)時(shí)間。統(tǒng)計(jì)爭(zhēng)斗持續(xù)時(shí)間內(nèi)爭(zhēng)斗行為發(fā)生次數(shù), 爭(zhēng)斗行為的定義如表1所示。

表1 三種爭(zhēng)斗行為的描述
1.6 血淋巴和組織的采集
首先, 選取規(guī)格一致的未爭(zhēng)斗個(gè)體直接取樣, 而未注射個(gè)體以及注射個(gè)體待爭(zhēng)斗結(jié)束后立即取樣。取樣時(shí)將蟹置于冰盤(pán)上, 用1 mL注射器預(yù)先吸入預(yù)冷抗凝劑, 再?gòu)挠斡咀慊砍槿?.1 mL血淋巴, 4 000 r/min離心20 min, 取上清; 另取肌肉、肝胰腺、胸神經(jīng)節(jié)3種組織樣品, 放入加有RNA保護(hù)液的1.5 mL無(wú)酶離心管中, 用以提取RNA, 并置于–80 °C保存。
1.7 5-HT、DA含量檢測(cè)
使用上海科興有限公司生產(chǎn)的蟹酶聯(lián)免疫分析(ELISA)試劑盒, 應(yīng)用雙抗體夾心法測(cè)定蟹血淋巴中5-HT和DA水平。
1.8 RT-qPCR分析
利用TRIzol法提取總RNA并合成第一鏈cDNA, 具體操作參考HiFiscript cDNA第一鏈合成試劑盒(康為)。在NCBI上查找與三疣梭子蟹相近物種的、、、、基因序列, 找到凡納濱對(duì)蝦和中華絨螯蟹()的相關(guān)基因并利用DNAMAN進(jìn)行序列對(duì)比, 根據(jù)同源性高的區(qū)域。利用primier5.0設(shè)計(jì)特異性引物后PCR獲得三疣梭子蟹相關(guān)基因核心片段。根據(jù)核心序列設(shè)計(jì)實(shí)時(shí)熒光定量PCR引物, 并檢測(cè)5-HT、DA上述受體基因的表達(dá)水平。目的基因進(jìn)行Real-time qPCR的反應(yīng)條件與內(nèi)參基因一致。數(shù)據(jù)采集在Roche LC480上進(jìn)行, 反應(yīng)結(jié)束后確定Real-time qPCR的擴(kuò)增曲線(xiàn)和熔解曲線(xiàn)。數(shù)據(jù)處理采用2–ΔΔCt法, 且用SPSS 19.0軟件進(jìn)行數(shù)據(jù)統(tǒng)計(jì)分析。實(shí)驗(yàn)所用引物序列見(jiàn)表2。
1.9 數(shù)據(jù)處理與統(tǒng)計(jì)分析
采用SPSS19.0軟件進(jìn)行單因子方差分析(one-way ANOVE), 比較分析各組間的差異, 使用Excel軟件作圖, 以上標(biāo)小寫(xiě)字母不同表示具有顯著性差異(<0.05)。
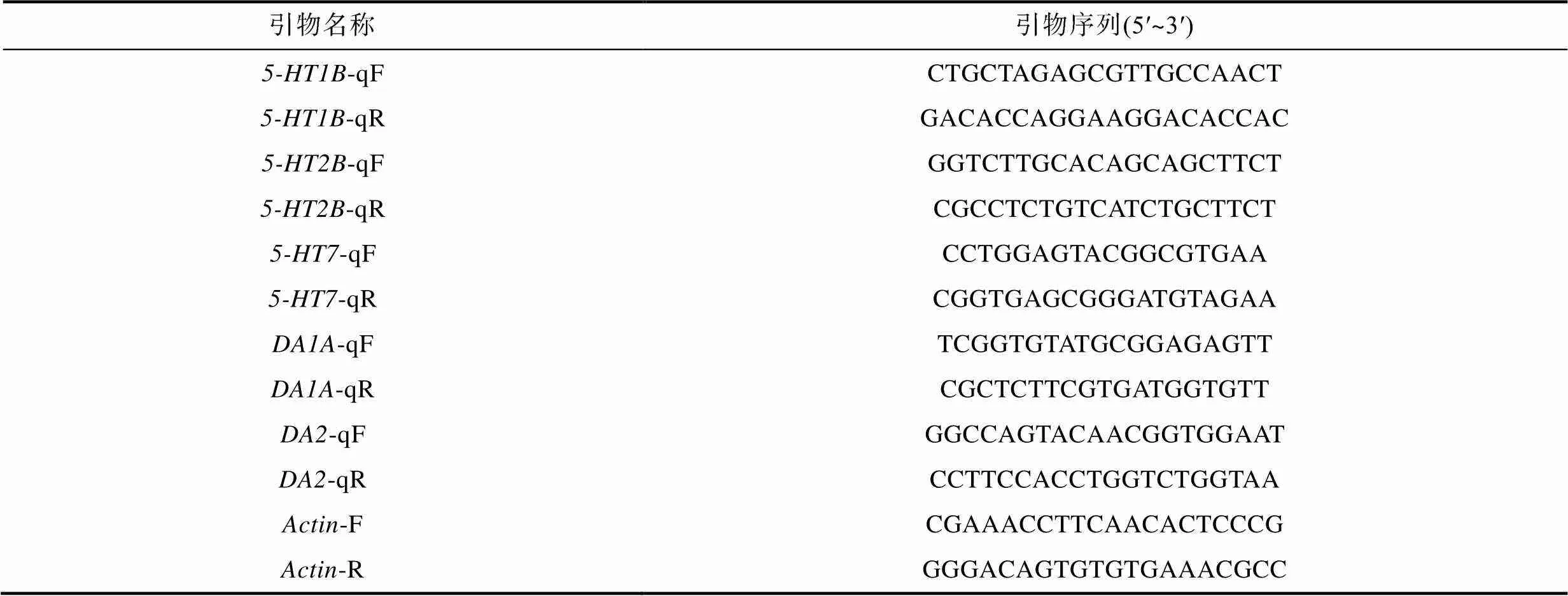
表2 實(shí)驗(yàn)中用到的引物列表
2 結(jié)果
2.1 爭(zhēng)斗前后血淋巴中5-HT、DA含量變化
爭(zhēng)斗前后三疣梭子蟹血淋巴中5-HT和DA含量均存在顯著性差異。爭(zhēng)斗后三疣梭子蟹血淋巴中5-HT含量呈顯著上升趨勢(shì)(<0.05), 比爭(zhēng)斗之前的上升19.5%; DA含量則呈現(xiàn)顯著下降的趨勢(shì)(<0.05), 比爭(zhēng)斗之前下降11.3%, 如圖2所示。
2.2 5-HT和DA對(duì)爭(zhēng)斗行為的影響
注射外源5-HT和DA對(duì)三疣梭子蟹爭(zhēng)斗行為發(fā)生次數(shù)的影響見(jiàn)表3。與生理鹽水組或?qū)φ战M相比, 注射10–6mol/只5-HT組三疣梭子蟹的夾擊、擊打和推搡發(fā)生次數(shù)都顯著減少(<0.05)。而與生理鹽水組相比, 注射10–8mol/只5-HT和10–10mol/只5-HT組三疣梭子蟹僅擊打和推搡發(fā)生次數(shù)顯著降低(<0.05), 而夾擊次數(shù)無(wú)明顯變化(>0.05)。同時(shí), 與生理鹽水組或?qū)φ战M相比, 注射10–6mol/只 DA組三疣梭子蟹的夾擊, 擊打和推搡發(fā)生次數(shù)都顯著增多(<0.05), 而注射10–8mol/只DA組和10–10mol/只DA組三疣梭子蟹的爭(zhēng)斗行為發(fā)生次數(shù)則無(wú)明顯變化(>0.05)。
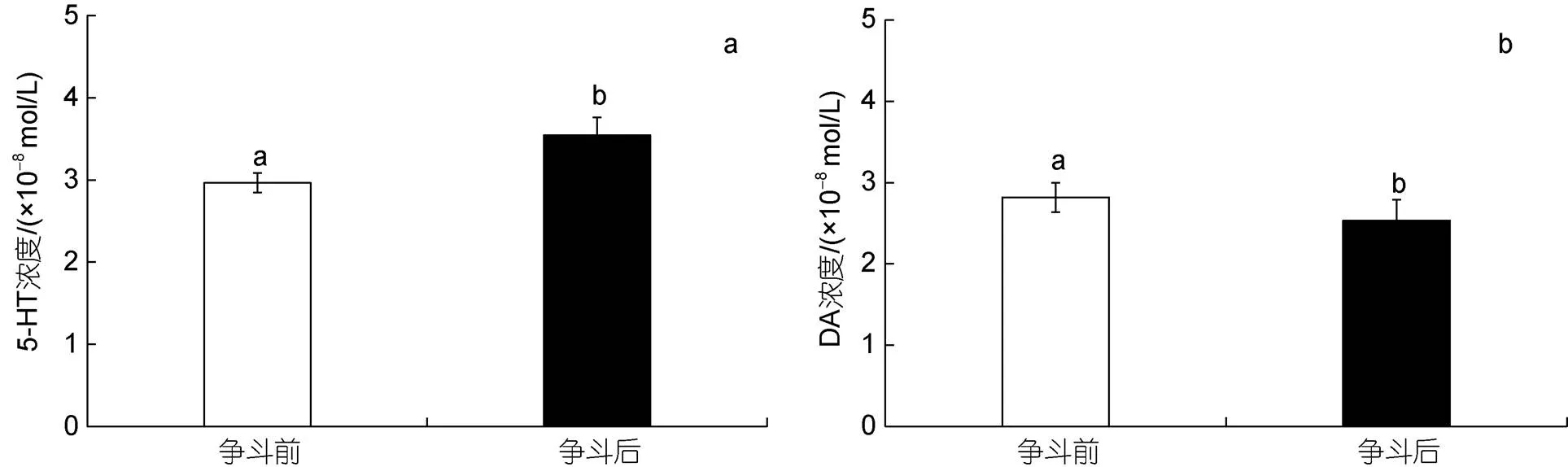
圖2 爭(zhēng)斗前后血淋巴中5-HT (a)和DA (b)含量變化
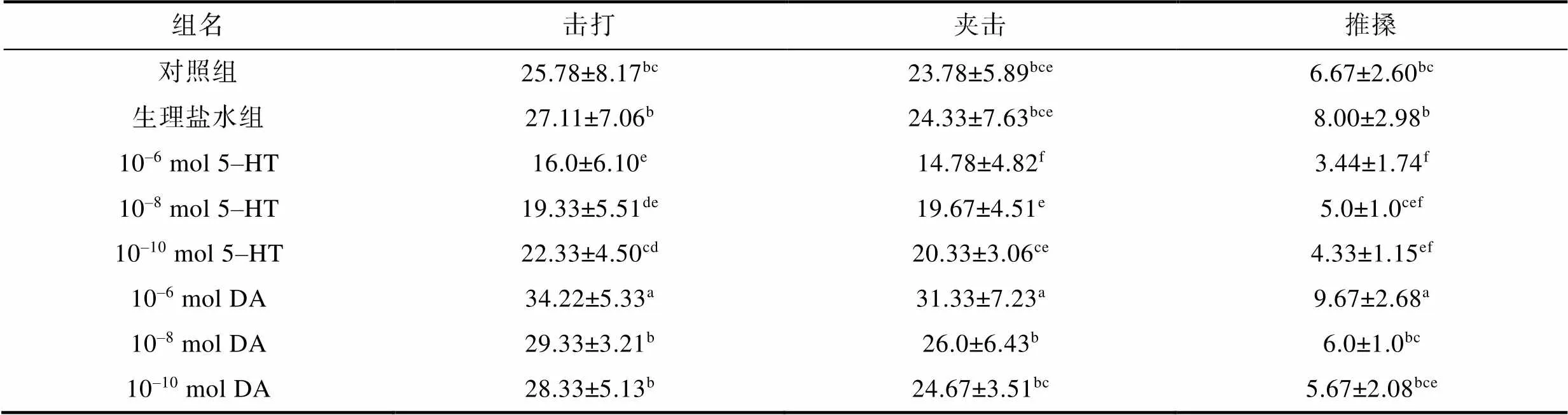
表3 注射5-HT和DA對(duì)爭(zhēng)斗行為發(fā)生次數(shù)的影響
注: 表中同列不同行上標(biāo)字母不同表示存在顯著差異(<0.05)
此外, 如圖3所示, 注射10–6mol/只5-HT組相較于注射生理鹽水組三疣梭子蟹的爭(zhēng)斗時(shí)間減少了約26%, 僅持續(xù)約38 min, 存在顯著性差異(<0.05), 相較于空白對(duì)照組三疣梭子蟹的爭(zhēng)斗時(shí)間縮短了約24%, 存在顯著性差異(<0.05), 而注射10–8mol/只5-HT組和10–10mol/只5-HT組三疣梭子蟹的爭(zhēng)斗時(shí)間則無(wú)明顯變化(>0.05); 注射10–6mol/只DA組相較于注射生理鹽水組三疣梭子蟹的爭(zhēng)斗時(shí)間分別增加了約20%, 超過(guò)60 min, 存在顯著性差異(<0.05)。而注射10–8mol/只DA組和10–10mol/只DA組的爭(zhēng)斗時(shí)間, 相較于生理鹽水組有所增加, 但無(wú)顯著差異(>0.05)。
2.3 爭(zhēng)斗前后5-HT和DA受體基因表達(dá)分析
利用實(shí)時(shí)熒光定量PCR分別檢測(cè)未注射的三疣梭子蟹爭(zhēng)斗前后5-HT和DA相關(guān)受體基因在肌肉、肝胰腺和胸神經(jīng)節(jié)中的表達(dá)分析。結(jié)果如圖4所示, 爭(zhēng)斗后mRNA表達(dá)量在僅肌肉中顯著增強(qiáng)(<0.05, 圖4a), 在胸神經(jīng)節(jié)中有降低趨勢(shì), 但無(wú)顯著變化(>0.05, 圖4a)。mRNA表達(dá)量在肌肉和胸神經(jīng)節(jié)中均顯著降低(<0.05, 圖4b)。而mRNA表達(dá)量?jī)H在胸神經(jīng)節(jié)中顯著增強(qiáng)(<0.05, 圖4c), 在肌肉中無(wú)明顯變化(>0.05, 圖4c)。爭(zhēng)斗后,和mRNA表達(dá)量在肌肉中顯著增強(qiáng)(<0.05, 圖4d, 4e), 在胸神經(jīng)節(jié)中顯著降低(<0.05, 圖4d, 4e)。
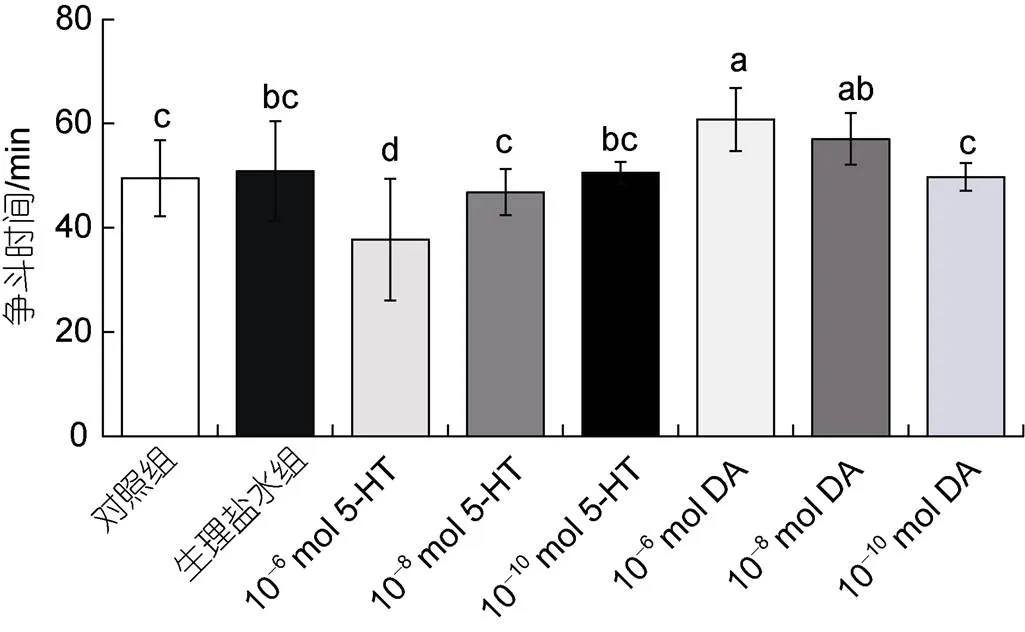
圖3 注射5-HT和DA對(duì)爭(zhēng)斗時(shí)間的影響
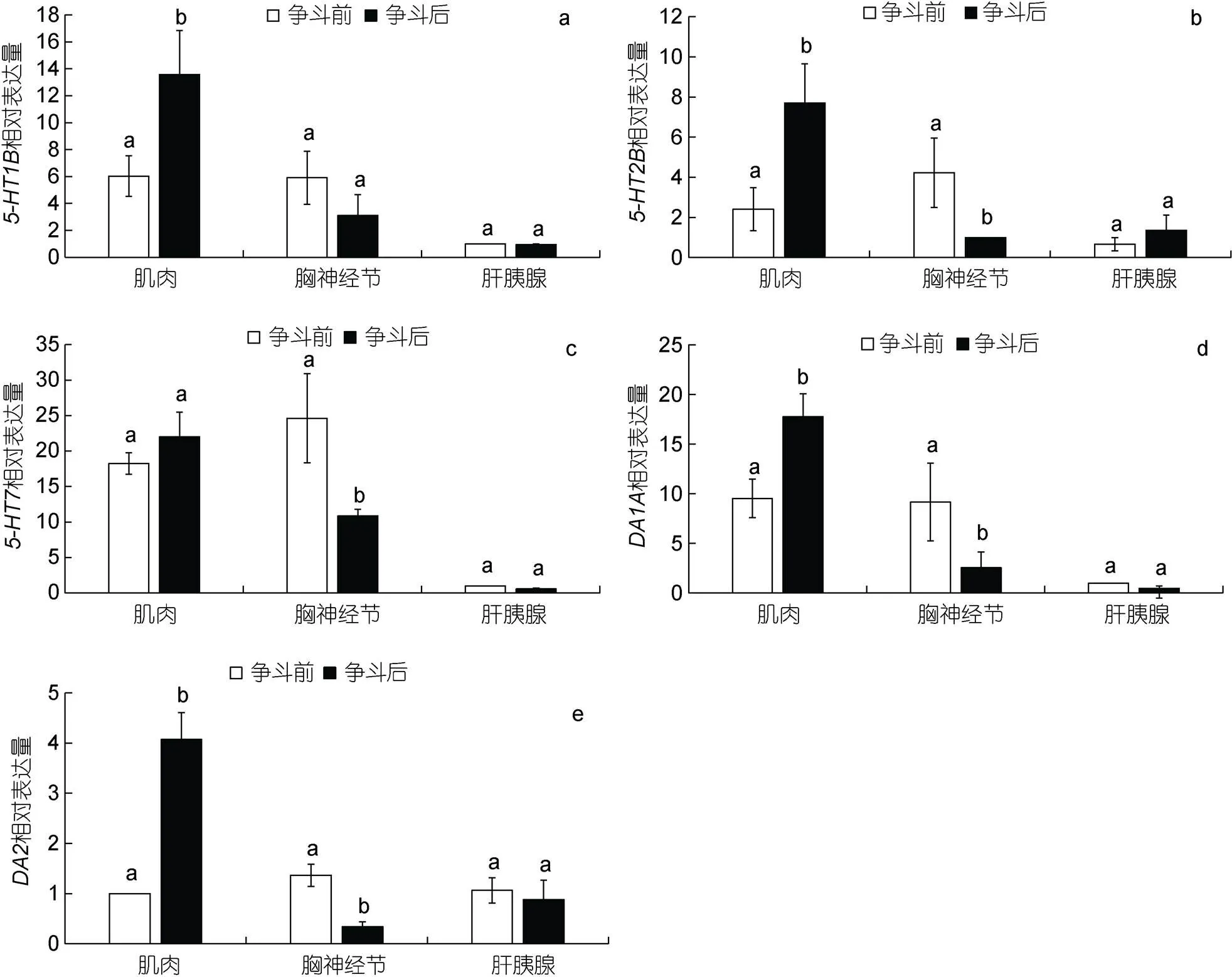
圖4 爭(zhēng)斗前后5-HT和DA受體基因在不同組織中的表達(dá)分析
注: a:, b:, c:, d:, e:
2.4 注射后5-HT和DA相關(guān)受體基因的表達(dá)分析
對(duì)5-HT和DA相關(guān)受體基因在對(duì)照組、生理鹽水組、10–6mol/只5-HT和10–6mol/只DA組的肌肉和胸神經(jīng)節(jié)中的相對(duì)表達(dá)量進(jìn)行分析分析。結(jié)果表明, 10–6mol/只5-HT注射后, 肌肉和胸神經(jīng)節(jié)中和mRNA表達(dá)量均顯著降低(<0.05, 圖5a, 5c), 而mRNA表達(dá)量在肌肉和胸神經(jīng)節(jié)中均無(wú)顯著變化(>0.05, 圖5b)。同時(shí), 注射5-HT后,mRNA表達(dá)量在肌肉中顯著降低(<0.05, 圖5d), 在胸神經(jīng)節(jié)中無(wú)顯著變化(>0.05, 圖5d)。而mRNA表達(dá)量在肌肉中顯著降低(<0.05, 圖5e), 在胸神經(jīng)節(jié)中顯著升高(<0.05, 圖5e)。在10–6mol/只DA注射后,mRNA表達(dá)量在肌肉中顯著降低(<0.05, 圖5a), 但在胸神經(jīng)節(jié)中無(wú)顯著變化(>0.05, 圖5a)。而和mRNA表達(dá)量在肌肉中均顯著降低(<0.05, 圖5b, 5c), 在胸神經(jīng)節(jié)中顯著升高(<0.05, 圖5b, 5c)。在注射DA后,mRNA表達(dá)量在肌肉中顯著降低(<0.05, 圖5d), 在胸神經(jīng)節(jié)中無(wú)明顯變化(>0.05, 圖5d)。而mRNA表達(dá)量在胸神經(jīng)節(jié)中顯著升高(<0.05, 圖5e), 在肌肉則顯著降低(<0.05, 圖5e)。以上結(jié)果表明注射5-HT和DA會(huì)引起相應(yīng)受體表達(dá)量的變化, 進(jìn)而參與三疣梭子蟹爭(zhēng)斗行為的調(diào)節(jié)。
3 討論
3.1 5-HT和DA對(duì)爭(zhēng)斗行為的影響
5-HT和DA作為神經(jīng)調(diào)節(jié)劑能夠參與甲殼動(dòng)物爭(zhēng)斗行為的調(diào)控。有研究表明, 5-HT和DA能夠通過(guò)G-蛋白偶聯(lián)受體調(diào)節(jié)環(huán)磷酸腺苷(cAMP)的釋放, 進(jìn)而影響個(gè)體的爭(zhēng)斗行為(Momohara, 2013), 而兩者對(duì)爭(zhēng)斗行為的影響也不相同。Pang等(2019)對(duì)中華絨螯蟹的研究發(fā)現(xiàn), 5-HT和DA之間存在一定的拮抗作用。同樣, 本實(shí)驗(yàn)結(jié)果發(fā)現(xiàn), 三疣梭子蟹爭(zhēng)斗后血淋巴中5-HT的含量顯著上升, 而DA的含量則顯著降低, 存在相反的變化趨勢(shì)。以上結(jié)果均表明5-HT和DA之間可能存在拮抗作用, 但兩者對(duì)爭(zhēng)斗行為的確切影響還需要進(jìn)一步研究。因此, 本研究通過(guò)注射不同濃度的5-HT和DA來(lái)進(jìn)一步探究?jī)烧邔?duì)三疣梭子蟹爭(zhēng)斗行為的影響。我們發(fā)現(xiàn)注射10–6mol/只5-HT能夠顯著減少三疣梭子蟹的爭(zhēng)斗時(shí)間和爭(zhēng)斗行為的發(fā)生次數(shù), 而注射10–6mol/只DA后三疣梭子蟹的爭(zhēng)斗時(shí)間和爭(zhēng)斗行為發(fā)生次數(shù)則都顯著減少。這些結(jié)果表明5-HT對(duì)三疣梭子蟹爭(zhēng)斗行為起抑制作用, 而DA對(duì)三疣梭子蟹爭(zhēng)斗行為起促進(jìn)作用。

圖5 注射5-HT和DA后相關(guān)受體基因在不同組織中的表達(dá)分析
注. a:; b.; c.; d:; e.
因此, 從本實(shí)驗(yàn)結(jié)果來(lái)看, 5-HT和DA作為神經(jīng)遞質(zhì)參與了三疣梭子蟹的爭(zhēng)斗行為, 并分別對(duì)三疣梭子蟹爭(zhēng)斗行為起到了抑制和促進(jìn)作用。如果在實(shí)際養(yǎng)殖過(guò)程中, 通過(guò)提高三疣梭子蟹體內(nèi)5-HT含量或降低DA含量, 從而減少其爭(zhēng)斗現(xiàn)象的發(fā)生, 以達(dá)到提高產(chǎn)量的目的。例如已有研究學(xué)者通過(guò)給擬穴青蟹()飼喂色氨酸(TRP)來(lái)增加血淋巴中5-HT的含量, 從而抑制爭(zhēng)斗行為的發(fā)生, 提高個(gè)體的存活率(Laranja, 2010)。
3.2 5-HT和DA相關(guān)受體基因參與三疣梭子蟹爭(zhēng)斗行為的調(diào)控
遺傳因素也是影響甲殼動(dòng)物爭(zhēng)斗行為的關(guān)鍵。5-HT和DA都是脊椎動(dòng)物和無(wú)脊椎動(dòng)物中高度保守的神經(jīng)遞質(zhì), 5-HT和DA相關(guān)受體在甲殼類(lèi)動(dòng)物中也高度保守。已有研究表明蝦蟹中存在5-HT和DA受體, 且能夠參與爭(zhēng)斗行為的調(diào)節(jié)(Yang, 2019)。徐澤文等(2015)克隆了中華絨螯蟹()和受體基因片段。而De Boer等(2005)將5-HT受體家族中與爭(zhēng)斗行為密切相關(guān)的和受體基因敲除時(shí), 發(fā)現(xiàn)褐家鼠()的攻擊行為顯著增加。5-HT和DA受體包含多種類(lèi)型, 且不同的5-HT和DA受體具有不同的表達(dá)機(jī)制, 其調(diào)控機(jī)理尚不清晰(Briffa, 2007)。本實(shí)驗(yàn)結(jié)果顯示,、、、和受體在不同組織中的表達(dá)量存在明顯差異(圖3)。在胸神經(jīng)和肌肉組織中的表達(dá)量較高, 在肝胰腺中表達(dá)量較低且無(wú)明顯變化。爭(zhēng)斗前后5-HT和DA受體在肌肉和胸神經(jīng)節(jié)中表達(dá)量發(fā)生顯著變化, 表明其可能主要通過(guò)這兩種組織發(fā)揮作用。5-HT和DA多個(gè)受體在相同組織中未呈現(xiàn)相同的變化規(guī)律, 表明各個(gè)受體在爭(zhēng)斗行為中的作用可能是不同的。爭(zhēng)斗前后5-HT和DA受體基因在肌肉和胸神經(jīng)中變化明顯, 這可能與爭(zhēng)斗行為受神經(jīng)系統(tǒng)調(diào)控, 而肌肉則是直接參與爭(zhēng)斗行為的發(fā)生有關(guān)。不同受體在爭(zhēng)斗過(guò)程中發(fā)揮作用不同且在不同的組織中的作用也不同, 這方面還有待于進(jìn)一步研究。
5-HT和DA通過(guò)與鳥(niǎo)嘌呤核苷酸結(jié)合和G蛋白偶聯(lián)受體蛋白相互作用介導(dǎo)許多社會(huì)行為(Weitekamp, 2017)。本研究也證實(shí)了注射5-HT和DA可以調(diào)節(jié)肌肉和胸神經(jīng)節(jié)中、、、和受體的表達(dá)水平(圖5)。表明5-HT和DA可能通過(guò)其相應(yīng)受體來(lái)參與爭(zhēng)斗行為的調(diào)節(jié)。同時(shí), 我們也發(fā)現(xiàn)DA能夠調(diào)節(jié)、、受體的表達(dá)水平(圖5a, 5b, 5c),和受體的表達(dá)水平也受到5-HT的調(diào)節(jié)(圖5d, 5e)。這也證實(shí)了5-HT和DA之間存在相互作用。為進(jìn)一步驗(yàn)證5-HT和DA相關(guān)受體參與爭(zhēng)斗行為的調(diào)節(jié), Pang等(2019)通過(guò)向中華絨螯蟹體內(nèi)注射抑制劑(ketanserin)和受體激動(dòng)劑[R(?)-TNPA]來(lái)驗(yàn)證它們對(duì)爭(zhēng)斗行為的影響, 發(fā)現(xiàn)抑制的活性, 能夠減少中華絨螯蟹的爭(zhēng)斗行為, 而激活受體能夠顯著降低中華絨螯蟹的爭(zhēng)斗行為。相反, Johnson等(2009)對(duì)黑腹果蠅()爭(zhēng)斗行為的研究發(fā)現(xiàn), 激活受體同樣也會(huì)降低個(gè)體的爭(zhēng)斗行為。Guo等(2015)也發(fā)現(xiàn)注射拮抗劑或iRNA敲除受體, 會(huì)增加?xùn)|亞飛蝗()的運(yùn)動(dòng)能力。以上結(jié)果進(jìn)一步證實(shí)和受體參與甲殼動(dòng)物爭(zhēng)斗行為的調(diào)節(jié), 但對(duì)不同甲殼動(dòng)物的調(diào)節(jié)存在種間差異性。這也與我們的研究發(fā)現(xiàn)相一致, 但更清晰的關(guān)系還需要進(jìn)一步探索。
4 結(jié)論
本研究結(jié)果表明, 外源5-HT對(duì)三疣梭子蟹爭(zhēng)斗行為起抑制作用, 而DA對(duì)三疣梭子蟹爭(zhēng)斗行為起促進(jìn)作用。同時(shí), 外源5-HT和DA會(huì)引起相關(guān)受體表達(dá)量變化, 進(jìn)而參與三疣梭子蟹爭(zhēng)斗行為的調(diào)控, 且它們之間存在相互作用。
王芳, 劉晶晶, 劉大鵬, 2020. 甲殼動(dòng)物爭(zhēng)斗行為研究進(jìn)展[J]. 中國(guó)海洋大學(xué)學(xué)報(bào), 50(2): 31-36.
孔雀, 邰發(fā)道, 2006. 攻擊行為神經(jīng)機(jī)制的研究進(jìn)展[J]. 現(xiàn)代生物醫(yī)學(xué)進(jìn)展, 6(8): 55-58.
李藝, 李曉東, 2019. 甲殼動(dòng)物攻擊行為的研究進(jìn)展[J]. 水產(chǎn)科學(xué), 38(3): 428-434.
李玉全, 孫霞, 2013. 水生動(dòng)物的爭(zhēng)勝行為[J]. 動(dòng)物學(xué)研究, 34(3): 214-220.
趙玉超, 秦浩, 李娜, 等, 2017. 神經(jīng)遞質(zhì)多巴胺(DA)和5-羥色胺(5-HT)對(duì)三種主要養(yǎng)殖對(duì)蝦爭(zhēng)勝行為的影響[J]. 海洋與湖沼, 48(3): 628-633.
段云峰, 吳曉麗, 王濤, 等, 2013. 五羥色胺和多巴胺與攻擊行為的關(guān)聯(lián)研究進(jìn)展[J]. 生命科學(xué), 25(10): 1027-1035.
徐澤文, 楊筱珍, 黃堅(jiān), 等, 2015. CODEHOP法設(shè)計(jì)引物克隆中華絨螯蟹5-ht和5-ht受體基因片段及序列分析[J]. 生物學(xué)雜志, 32(1): 1-5.
BACQUé-CAZENAVE J, CATTAERT D, DELBECQUE J P,, 2018. Serotonin has opposite effects on the aggressiveness of crayfish, facing either a smaller or a larger rival: alteration of size perception [J]. Journal of Experimental Biology, 221(12): jeb177840.
BORTOLATO M, PIVAC N, SELER D M,, 2013. The role of the serotonergic system at the interface of aggression and suicide [J]. Neuroscience, 236: 160-185.
BRIFFA M, ELWOOD R W, 2007. Monoamines and decision making during contests in the hermit crab[J]. Animal Behaviour, 73(4): 605-612.
COSTA J R, DALOSTO M M, PALAORO A V,, 2016. Contest duration and dynamics are affected by body size in a potentially subsocial crayfish (Crustacea: Decapoda) [J]. Ethology, 122(6): 502-512.
DE BOER S F, KOOLHAAS J M, 2005. 5-HT1Aand 5-HT1Breceptor agonists and aggression: a pharmacological challenge of the serotonin deficiency hypothesis [J]. European Journal of Pharmacology, 526(1/3): 125-139.
GUO X J, MA Z Y, KANG L, 2015. Two dopamine receptors play different roles in phase change of the migratory locust [J]. Frontiers in Behavioral Neuroscience, 9: 80.
HAMASAKI K, SUGIYAMA A, JINBO T,, 2018. The influence of male size on competitive mating success in the Japanese spiny lobster(von Siebold, 1824) (Decapoda: Palinuridae): implications for broodstock management techniques [J]. Journal of Crustacean Biology, 38(4): 393-400.
HUBER R, PANKSEPP J B, YUE Z,, 2001. Dynamic interactions of behavior and amine neurochemistry in acquisition and maintenance of social rank in crayfish [J]. Brain, Behavior and Evolution, 57(5): 271-282.
JOHNSON O, BECNEL J, NICHOLS C D, 2009. Serotonin 5-HT2and 5-HT1A-like receptors differentially modulate aggressive behaviors in[J]. Neuroscience, 158(4): 1292-1300.
LARANJA J L Q, QUINITIO E T, CATACUTAN M R,, 2010. Effects of dietaryL-tryptophan on the agonistic behavior, growth and survival of juvenile mud crab[J]. Aquaculture, 310(1/2): 84-90.
LEE S H C, TAYLOR K, KRASNE F B, 2008. Reciprocal stimulation of decay between serotonergic facilitation and depression of synaptic transmission [J]. Journal of Neurophysiology, 100(2): 1113-1126.
MOMOHARA Y, KANAI A, NAGAYAMA T, 2013. Aminergic control of social status in crayfish agonistic encounters [J]. PLoS One, 8(9): e74489.
NORTHCUTT A J, LETT K M, GARCIA V B,, 2016. Deep sequencing of transcriptomes from the nervous systems of two decapod crustaceans to characterize genes important for neural circuit function and modulation [J]. BMC Genomics, 17(1): 868.
PANG Y Y, SONG Y M, ZHANG L,, 2019. 5-HT2B, 5-HT7, and DA2 receptors mediate the effects of 5-HT and DA on agonistic behavior of the Chinese mitten crab () [J]. ACS Chemical Neuroscience, 10(11): 4502-4510.
RODGERS E W, KRENZ W D, JIANG X Y,, 2013. Dopaminergic tone regulates transient potassium current maximal conductance through a translational mechanism requiring D1Rs, cAMP/PKA, Erk and mTOR [J]. BMC Neuroscience, 14(1): 143.
SERRETTI A, CALATI R, MANDELLI L,, 2006. Serotonin transporter gene variants and behavior: a comprehensive review [J]. Current Drug Targets, 7(12): 1659-1669.
TIERNEY A, MANGIAMELE L, 2001. Effects of serotonin and serotonin analogs on posture and agonistic behavior in crayfish [J]. Journal of Comparative Physiology A, 187(10): 757-767.
TINA F W, JAROENSUTASINEE M, KEERATIPATTARAKARN K,, 2018. Sex and burrow/chimney ownership affecting time allocation for surface activities in(Tweedie, 1937) (Brachyura, Ocypodidae) [J]. Crustaceana, 91(1): 51-62.
WEITEKAMP C A, NGUYEN J, HOFMANN H A, 2017. Social context affects behavior, preoptic area gene expression, and response to D2 receptor manipulation during territorial defense in a cichlid fish [J]. Genes, Brain and Behavior, 16(6): 601-611.
YANG X Z, PANG Y Y, HUANG G Y,, 2019. The serotonin or dopamine by cyclic adenosine monophosphate-protein kinase A pathway involved in the agonistic behaviour of Chinese mitten crab,[J]. Physiology & Behavior, 209: 112621.
ZHU F, FU Y Y, MU C K,, 2018. Molecular cloning, characterization and effects of catechol--methyltransferase (comt) mRNA and protein on aggressive behavior in the swimming crab[J]. Aquaculture, 495: 693-702.
PRELIMINARY STUDY ON THE REGULATION OF AGONISTIC BEHAVIOR OFBY EXOGENOUS 5-HT AND DA
WU Bo1, ZHANG Shun1, ZHANG Cheng1, MU Chang-Kao1, XU Shan-Liang1, 2, WANG Dan-Li1
(1. School of Marine Science, Ningbo University, Ningbo 315211, China; 2. Key Laboratory of Applied Marine Biotechnology, Ministry of Education, Ningbo University, Ningbo 315211, China)
To explore the behavioral mechanism of damage or death ofdue to agonistic behavior during culture and reproduction, contents of 5-HT and DA in hemolymph ofbefore and after fight were determined by double antibody sandwich method, and the changes of 5-HT and DA receptor gene expression in muscle, thoracic ganglion, and hepatopancreas before and after fight were analyzed. Meanwhile, the difference of agonistic behavior and the changes of 5-HT and DA receptor gene expression in the three tissues were recorded and analyzed by injecting exogenous 5-HT and DA. Results show that, after fighting, the content of 5-HT in hemolymph ofincreased significantly, while the content of DA decreased significantly (<0.05). Injection of 10–6-mol 5-HT could significantly reduce the frequency and duration of agonistic behavior (<0.05), while injection of 10–6-mol DA could significantly increase the frequency and duration of agonistic behavior of(<0.05). The expression of 5-HT and DA receptor in muscle and thoracic ganglion changed significantly before and after fight, and different receptors did not show the same trend in the same tissue. Exogenous 5-HT could also regulate the expression ofandreceptors, and exogenous DA could regulate the expression of,, andreceptors. Therefore, 5-HT can inhibit the agonistic behavior of, while DA can promote the agonistic behavior. In addition, 5-HT and DA receptor genes can participate in the regulation of agonistic behavior ofthrough 5-HT/DA, interactively.
; agonistic behavior; 5-HT/DA;receptor
S967; Q958.12
10.11693/hyhz20210600132
*浙江省重大科技專(zhuān)項(xiàng), 2019C02057號(hào); 國(guó)家自然科學(xué)基金項(xiàng)目, 42076087號(hào)。吳 波, 碩士研究生, E-mail: 1035757132@qq.com
徐善良, E-mail: xushanliang@nbu.edu.cn; 王丹麗, E-mail: wangdanli@nbu.edu.cn
2021-06-08,
2021-08-13
猜你喜歡
健康之家(2025年11期)2025-08-27 00:00:00
大自然探索(2025年7期)2025-08-03 00:00:00
中國(guó)圖書(shū)評(píng)論(2022年4期)2022-11-22 08:59:08
世界科學(xué)技術(shù)-中醫(yī)藥現(xiàn)代化(2022年3期)2022-08-22 00:27:56
中國(guó)獸醫(yī)雜志(2022年5期)2022-08-06 08:19:34
右江醫(yī)學(xué)(2022年3期)2022-04-17 11:42:53
大眾健康(2021年2期)2021-03-09 13:32:23
中國(guó)中藥雜志(2016年23期)2017-04-07 23:03:47
飼料博覽(2017年3期)2017-04-04 11:25:24
湖南中醫(yī)藥大學(xué)學(xué)報(bào)(2015年1期)2016-01-06 01:06:40

- 海洋與湖沼的其它文章
- 兩種混養(yǎng)方式對(duì)異枝江蘺(Gracilaria bailinae)生長(zhǎng)性能、表面附生細(xì)菌群落及抗生素抗性基因的影響*
- 基于無(wú)人機(jī)高光譜遙感技術(shù)對(duì)內(nèi)陸?zhàn)B殖池塘水質(zhì)監(jiān)測(cè)的研究*
- 仿生態(tài)條件下星洲紅魚(yú)(Singapore red tilapia)和奧尼羅非魚(yú)(Oreochromis niloticus)生長(zhǎng)和肌肉營(yíng)養(yǎng)特性的比較研究*
- 印度洋赤道潛流年際變化特征及其與印度洋偶極子的聯(lián)系*
- 太平洋內(nèi)部副熱帶-熱帶經(jīng)向翻轉(zhuǎn)環(huán)流的季節(jié)變化特征*
- 凡納濱對(duì)蝦(Litopenaeus vannamei)不同品系生長(zhǎng)與耐高鹽性狀配合力及雜種優(yōu)勢(shì)分析*