人類糖脂轉運蛋白家族成員在疾病發生發展中的作用
2022-03-02 05:36:34張湘豫盧夢云鄒先瓊
醫學綜述 2022年2期
張湘豫,盧夢云,鄒先瓊
(1.桂林醫學院附屬口腔醫院口腔醫學研究所,廣西 桂林 541004; 2.桂林醫學院基礎醫學院,廣西 桂林 541004)
鞘脂屬于一類脂質,最初被描述為細胞膜結構的主要組成部分,它們普遍存在且高度保守;鞘脂及其代謝產物是多種生理和生物活性的關鍵調節因子,如神經酰胺、鞘氨醇-1-磷酸、神經酰胺-1-磷酸(ceramide-1-phosphate,C1P),能參與細胞增殖、生存、胚胎發育、器官形成和免疫細胞活動調節等重要生理功能,并參與了多種病理過程,特別是一些炎癥反應或相關的疾病[1-2]。此外,鞘脂含量或活性的改變在癌細胞代謝中發揮重要作用。糖脂轉運蛋白(glycolipid transfer protein,GLTP)被認為在葡萄糖神經酰胺代謝中發揮作用,其過表達導致細胞內葡萄糖神經酰胺的合成和轉移顯著增加[3]。而神經酰胺的處理不僅能部分阻斷由于轉錄因子特異性蛋白(specificity protein,Sp)1及Sp3敲除引起的啟動子活性下調,也能改變Sp1/Sp3與GLTP基因啟動子的親和性以及減少Sp3的乙酰化[4]。GLTP家族成員對于維持細胞內鞘脂的穩態極其重要,因此闡明GLTP家族成員的生物學功能對炎癥、腫瘤等相關疾病的防治具有重要意義[5]。已有文獻報道了一些GLTP家族成員的結構和生物學功能[6-8]。但GLTP家族成員在炎癥、腫瘤等疾病發生發展中的作用及其機制尚不清楚。現就人類GLTP家族成員的結構、功能及其在疾病發生發展中的作用予以綜述。
1 GLTP家族成員的結構和功能
為維持細胞內外脂質平衡穩定,脂質在細胞內通過囊泡和非囊泡的機制轉運至生物膜[5]。而脂質膜間運輸的非囊泡機制主要依賴脂質轉運蛋白,脂質轉運蛋白在與特定膜的瞬時相互作用中獲取并傳遞其特定的脂質,脂質轉運蛋白只是暫時黏附在脂質膜上,在轉移過程中可作為脂質分子增溶劑包裹不溶性脂質來發揮作用[5]。研究發現,糖鞘脂(glycosphingolipids,GSLs)轉移蛋白家族成員共享一種獨特的結構折疊,稱為 GLTP折疊[9]。GLTP折疊是一種新穎的外周兩性折疊和結構上獨特的脂質結合基序,GLTP具有獨特的α-螺旋構象,排列在一個兩層“三明治”基序中,這種“三明治”狀結構被認為是真核生物GLTP家族的結構基礎,形成單一的“口袋狀”糖脂結合位點,即GLTP折疊(圖1),具有GLTP折疊的蛋白即GLTP家族成員[5,9-11]。GLTP家族的重要成員包括磷脂酰肌醇-4-磷酸銜接蛋白(phosphatidylinositol-4-phosphate adaptor protein,FAPP)2、人1-磷酸神經酰胺轉移蛋白(ceramide-1-phosphate transfer protein,CPTP)、促細胞死亡蛋白11及GLTP家族的一個新成員——人類糖脂轉運結構域2蛋白(glycolipid transfer protein domain containing protein 2,GLTPD2)[6,12]。其中,FAPP2不僅可以轉運葡萄糖神經酰胺,還可以轉運半乳糖神經酰胺和乳糖神經酰胺,而CPTP和促細胞死亡蛋白11能選擇性結合或轉移C1P[13]。同時,促細胞死亡蛋白11是自主神經酰胺水平的中間調節蛋白[14]。GLTP家族成員作為可以轉運某些鞘脂功能的蛋白在細胞功能代謝中起著至關重要的作用,因為鞘脂參與了細胞重要的代謝過程。從真菌到人類均存在GLTP家族成員,證明了GLTP家族的重要性[5]。由于GLTP家族成員與腫瘤、炎癥性疾病、神經退行性疾病等密切相關,因此探索GLTP家族成員的功能與疾病的作用關系,對這些疾病的防治具有重要意義。

圖1 糖脂轉運蛋白蛋白空間結構(PDB:1SX6)
2 GLTP家族成員與疾病的關系
有研究表明,GLTP家族成員參與了炎癥性疾病、腫瘤、神經退行性疾病等的發生發展過程[15-17]。因此,闡明GLTP家族成員的生物學功能及其在疾病發生發展中的作用具有重要意義。
2.1GLTP 人類GLTP是一種小的(分子量為24 000)兩性可溶性蛋白,介導各種GSLs的非囊泡狀轉運[6,10,18]。然而,迄今為止決定GLTP選擇性轉運特定GSLs的機制尚不清楚[18]。通過對GLTP的動力學研究表明,GLTP的作用不是GLTP分配于供體囊泡表面,而是通過轉移到膜界面,GLTP及時發現并識別GSLs上的糖基部分,然后形成GLTP-GSLs復合物,進而釋放到受體上并完成GSLs在膜間轉移[6,19](圖2)。

注:GLTP為糖脂轉運蛋白,GSLs為糖鞘脂,GLTP-GSLs Complex in solution為游離的糖脂轉運蛋白-糖鞘脂復合物,Glycolipid為糖脂類,Phosphoglyceride為磷酸甘油酯,Cytoplasm為細胞質圖2 GLTP介導的體外GSLs膜間轉移模型
通常,細胞形狀改變是細胞程序性死亡過程的表型指標,而人類GLTP基因表達的改變,不僅改變了細胞與細胞接觸和表面黏附的某些GSLs的穩態水平,而且還調節了某些人癌細胞株的細胞形狀變化,從而誘導了細胞程序性壞死的發生發展[5]。表達δ聯蛋白可誘導樹突生長,而GLTP基因與δ聯蛋白共表達可加速細胞向圓形形狀的轉變,這些發現是已知的首次由GLTP基因過表達引發的表型變化,并通過與p120聯蛋白家族成員的直接相互作用進行調控[20]。研究發現,在被活化的β聯蛋白轉化的小鼠乳腺上皮細胞中,GLTP基因的表達也會受到影響[21]。同時,通過雙氯醇上調GLTP與丙型肝炎病毒(hepatitis C virus,HCV)非結構5A蛋白(non-structural 5A protein,NS5A)競爭結合宿主囊泡相關膜蛋白相關蛋白A(vesicle-associated membrane protein-associated protein-A,VAP-A),導致VAP-A和HCV NS5A之間的復合物形成中斷,由于VAP-A/NS5A復合物的形成對病毒RNA的復制至關重要,因此GLTP降低了VAP-A/NS5A復合物的水平,從而抑制了HCV的復制[22]。研究證明,GLTP過表達能上調細胞周期蛋白依賴性激酶(cyclin-dependent kinase,CDK)抑制因子1A(p21)及1B(p27),下調CDK2和CDK4、細胞周期蛋白E和細胞周期蛋白D1,進而阻滯細胞周期于G1/S檢查點,誘導細胞程序性壞死,從而抑制人類結直腸癌HT-29細胞的生長[17]。同時,GLTP參與了一種罕見的神經退行性疾病中的中樞神經系統的髓鞘增生,該增生是C型尼曼-皮克病最明顯的病理特征之一[23]。有學者通過微陣列分析發現,GLTP基因是6 000個經篩選的乳腺組織互補DNA克隆中的12個基因之一,這些克隆在經過部分或完全轉分化為鱗狀化生的轉化小鼠乳腺上皮中轉錄上調[6]。在不影響結直腸癌HT-29細胞神經酰胺水平的情況下,GLTP基因上調可導致鞘氨醇-1-磷酸水平降低,而鞘氨醇-1-磷酸是調節細胞生長和抑制細胞凋亡的信號分子[4]。以上研究表明,GLTP與細胞周期及細胞程序性壞死密切相關,在丙型肝炎、神經退行性疾病、結直腸癌等疾病的發生發展中發揮重要作用。
2.2FAPP2 人類FAPP2是FAPP成員,具有Pleckstrin同源結構域,其基因位于染色體7p21,由519個氨基酸組成[16]。FAPP2定位于高爾基體反式面,可以將葡萄糖神經酰胺從高爾基體轉運到內質網,在復雜GSLs的合成中發揮關鍵作用[24]。同時,FAPP2-GLTPH結構揭示了一種控制GLTP家族特異性的元素,稱為ID環(individual or identification loop)。不同的ID環有不同的識別中心,可在家庭成員中產生不同的特異性[11,25]。
FAPP2在GSLs的代謝中發揮了重要作用,該蛋白與程序性細胞死亡過程的調節密切相關[5]。細胞凋亡是細胞死亡的一種特殊形式,它發生在正常和病理過程中,可由多種刺激引起[23]。通過干擾小RNA沉默FAPP2基因增強了抗Fas腫瘤細胞對Fas/Fas配體誘導的凋亡的敏感,其中Fas和Fas配體是腫瘤壞死因子死亡受體/配體家族成員,Fas配體與表達Fas的細胞結合可觸發其凋亡[26-27](圖3)。FAPP2基因已初步被血清學鑒定為潛在的乳腺癌抗原,很可能與乳腺癌密切相關[27-29]。FAPP2基因在結腸癌腫瘤組織中的表達增加,而FAPP2基因的表達下調抑制了結腸癌細胞的生長;同時研究發現,Wnt/β聯蛋白信號通路參與了FAPP2調控的腫瘤細胞生長,提示FAPP2可能作為癌基因調控腫瘤生長[30]。研究證明,利用CRISPR/Cas9(clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9)基因編輯技術敲除FAPP2基因能夠通過下調Wnt/β聯蛋白信號通路抑制肝癌細胞的增殖和侵襲,因此FAPP2基因被認為是一種與腫瘤發生密切相關的腫瘤相關調節因子[16]。研究發現,在HCV感染致病過程中,HCV NS5A會激活磷脂酰肌醇-4-激酶,導致磷脂酰肌醇-4-磷酸(phosphatidylinositol-4-phosphate,PI4P)脂質的大量生產和再分配到復制復合物中,PI4P招募宿主效應子來調節HCV基因組復制或病毒粒子的產生,而FAPP2具有對PI4P和糖脂的特異性功能脂質結合域,因此PI4P結合FAPP2的表達可抑制HCV復制,同時糖脂結合FAPP2蛋白導致復制效率顯著下降,表明這兩個結構域對FAPP2在HCV基因組復制中的作用均至關重要[31]。以上研究表明,FAPP2在細胞程序性死亡過程中具有重要的生物學功能,與丙型肝炎、肝癌、結腸癌等疾病的發生發展密切相關。
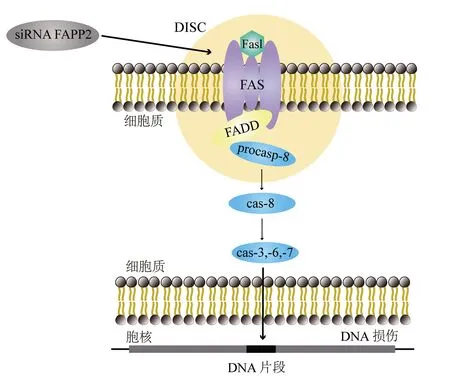
注:siRNA為干擾小RNA,FAPP2為磷脂酰肌醇-4-磷酸銜接蛋白,DISC為死亡誘導信號復合體,FasL為Fas配體,FADD為Fas相關死亡結構域蛋白,procasp-8為無活性的胱天蛋白酶8,cas為胱天蛋白酶圖3 FAPP2基因在腫瘤細胞中對Fas誘導的凋亡的模式圖
2.3CPTP 人類CPTP是由GLTPD1編碼的一種蛋白質,已被證明能形成GLTP折疊,其特征是修飾的
GLTP折疊層選擇性地結合C1P,而不是GSLs、神經酰胺或其他具有磷酸頭基的脂類[10,32]。C1P是由神經酰胺經位于反式高爾基體的神經酰胺激酶磷酸化而成的一類重要鞘脂分子,其通過與細胞膜上的Gi蛋白結合介導多條信號通路,從而產生誘導細胞增殖、干細胞動員、巨噬細胞遷移、誘導自噬介導的促炎細胞因子以及激活IVA胞質型磷脂酶A2α(cytosolic phospholipase A2α,cPLA2α)產生二十烷類化合物等生物活性作用[33-34]。IVA cPLA2α和二十烷類化合物被認為在炎癥和細胞損傷中發揮重要作用,C1P積累和IVA cPLA2α的激活導致花生四烯酸和促炎癥類花生酸的產生[34]。
CPTP是早期自噬體組裝和炎癥小體驅動的促炎細胞因子生成和釋放的內源性調節因子,但不是細胞凋亡的內源性調節因子,同時宮頸癌細胞系HeLa、人腎上皮細胞系HEK-293T以及單核巨噬細胞均表現出CPTP誘導的自噬,在單核巨噬細胞中,炎癥小體組裝和胱天蛋白酶1的強烈激活伴隨兩種促炎細胞因子(白細胞介素-1B和白細胞介素-18)的釋放顯著增加[1,33,35-36](圖4)。研究發現,下調CPTP基因可導致人肺癌A549細胞中促炎癥花生酸的產生[32]。在重癥胰腺炎中,CPTP基因可能通過下調IVA cPLA2α的表達影響緊密連接相關蛋白(即閉合小環蛋白1)的表達,保護腸上皮屏障;緊密連接是調節腸道通透性的結構基礎,可能在細菌易位中發揮重要作用[37]。近年研究表明,在結直腸癌SW480細胞、乳腺癌中,CPTP是腫瘤抑制因子微RNA(microRNA,miRNA/miR)-328的直接作用靶點[38]。且已有文獻報道,miR-328在許多腫瘤中低表達,提示CPTP可能通過靶向miR-328作用于腫瘤細胞而發揮抗腫瘤作用[39]。以上研究表明,CPTP在自噬體組裝和炎癥小體的形成中發揮了重要的生物學功能,且與炎癥性疾病、胰腺癌、乳腺癌等疾病的發生發展密切相關。
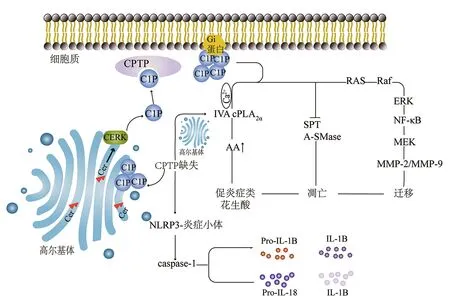
注:CPTP為人1-磷酸神經酰胺轉移蛋白,C1P為神經酰胺-1-磷酸,CERK為神經酰胺激酶,Cer為神經酰胺,NLRP3為核苷酸結合寡聚化結構域樣受體蛋白3,caspase-1為胱天蛋白酶1,IVA cPLA2α為IVA胞質型磷脂酶A2α,AA為花生四烯酸,Pro為前體,IL為白細胞介素,SPT為絲氨酸棕櫚酰轉移酶,A-SMase為酸性鞘磷脂酶,ERK為胞外信號調節激酶,NF-κB為核因子κB,MEK為促分裂原活化的蛋白激酶激酶,MMP為基質金屬蛋白酶圖4 CPTP缺失誘導炎癥小體的激活及C1P在哺乳動物細胞中的生物學功能
2.4GLTPD2 人類GLTPD2是GLTP家族的一個新成員,GLTPD2基因最初是由計算機生成的人類基因組注釋預測出來的[5]。有研究通過聚合酶鏈反應分析發現,GLTPD2信使RNA在肝臟組織中高表達[5,12]。GLTPD2基因是一種蛋白質編碼基因,與該基因有關的基因本體論注釋包括糖脂結合和GLTP活性,其重要旁系同源物為CPTP[5]。人類GLTPD2的GLTP結構域位于102~252氨基酸殘基,1~36氨基酸殘基為N端信號肽,13~32氨基酸殘基為跨膜區,人類GLTPD2分子量為31 600,等電點為10.19[6]。有研究表明,人類GLTPD2與肝、腸、腎等器官的功能及疾病的發生發展密切相關[12]。GLTPD2基因是新生小鼠壞死性小腸結腸炎中上調程度最高的15個基因之一,也是單聚體而非五聚體C反應蛋白處理內皮祖細胞時上調程度最高的12個基因之一,五聚C反應蛋白與單體C反應蛋白的分離被認為參與了動脈粥樣硬化斑塊發生部位的局部促炎癥反應[40-41]。在一項全基因關聯研究中,GLTPD2基因被確定為35個脂類相關基因位點中與心血管疾病風險相關的10個基因變體之一[41]。以上研究表明,GLTPD2與小腸結腸炎、心血管疾病等疾病的發生發展密切相關。
3 小結與展望
GLTP家族成員的鞘脂代謝“傳感器”功能極其重要,因為鞘脂參與了細胞的生死過程,即細胞增殖、分化、發育、凋亡、自噬、炎癥、焦亡、壞死等重要的生物學功能[5]。進化修飾導致GLTP折疊對C1P具有特異性(如CPTP)或對GSLs具有可變選擇性(如FAPP2和GLTP),而這些GLTP家族成員由不同人類染色體上具有不同內含子、外顯子組織的單拷貝基因編碼[10]。雖然關于GLTP及其相關的GLTP同源蛋白的結構、功能和基因組方面已有了較多研究,但這些胞質兩性鞘脂轉運蛋白在體內的功能仍不清楚[10,18,20,34]。
由于還有許多新的GSLs轉運蛋白成員尚未被發現,因此對鞘脂信號調控癌細胞信號轉導和代謝機制的進一步了解將有助于改進癌癥的治療方式[42]。了解鞘脂信號在與腫瘤微環境(包括基質細胞、內皮細胞、破骨細胞或血小板)中相關的腫瘤和宿主細胞之間通訊調控中的作用,將有助于開發新的治療策略來抑制癌癥的生長、增殖和轉移[2,6]。此外,還需要更多研究來探討如何改變鞘脂信號,以增加T細胞的抗腫瘤功能,并降低骨髓源性抑制細胞和腫瘤相關巨噬細胞的抑制功能,提高免疫治療的抗癌效果[42-43]。因此,尋找新的GSLs轉運蛋白對未來相關疾病的預防、診斷及治療非常重要。
已知GLTP家族成員在細胞周期、細胞程序性壞死、自噬體組裝和炎癥小體形成等生物學過程中發揮了重要作用,并與丙型肝炎、神經退行性疾病、結直腸癌、肝癌、乳腺癌、小腸結腸炎、心血管疾病等的發生發展密切相關。這些研究為今后涉及人類健康和疾病中的GLTP的跨學科、轉化醫學研究開辟了新途徑,如有針對性地調控特定的GLTP家族成員的表達水平,進而影響細胞內特異鞘脂的水平,作為相關疾病防治的有效手段。相信隨著研究的不斷深入,更多GLTP蛋白家族成員將會被發現,而闡明其生物學功能及其在疾病發生發展中的作用將為人類最終戰勝這些疾病做出重要貢獻。
