促生菌對Cd脅迫下蔞蒿光合特性的影響
2022-03-07 08:05:14周小梅余紅兵譚淑端
草地學報 2022年2期
周小梅, 董 萌, 余紅兵,, 譚淑端, 楊 楠
(1.湖南城市學院建筑與城市規劃學院, 湖南 益陽 413000; 2. 數字化城鄉空間規劃關鍵技術湖南省重點實驗室,湖南 益陽 413000; 3. 湖南省城鄉生態規劃與修復工程技術研究中心, 湖南 益陽 413000; 4.湖南省村鎮飲用水水質安全保障工程技術研究中心, 湖南 益陽 413000)
Cd是一種典型的土壤污染物,環保部和國土資源部2014年聯合發布的《全國土壤污染狀況調查公報》顯示:我國土壤環境中Cd點位超標率為7.0%[1]。Cd毒性極強,非生物體生命活動的必需元素,由于其極強的移動性,極易通過食物鏈危害人體的健康。相比于傳統的物理與化學方法,生物修復技術由于其經濟、環保、無污染等特點,是當前公認的一種環境友好型污染治理技術。
洞庭湖區是湖南省及至全國比較重要的糧食生產基地,其濕地土壤Cd污染較為嚴重[2-3]。蔞蒿(Artemisiaselengensis)是洞庭湖濕地優勢植物,多年生草本,菊科蒿屬,適合刈割、無性繁殖,對Cd具較強耐性,是洞庭湖濕地土壤實施Cd污染修復的理想選擇[4]。
促生菌(Plant growth-promoting bacteria,PGPB)能夠與植物建立良好的協同互惠關系,在重金屬污染土壤的修復過程中可發揮重要作用[5-7]。近年圍繞促生菌在重金屬污染土壤植物修復方面的運用已作了大量的研究,研究主要集中于對植物生長、解毒、生防、對重金屬的生物有效性及其轉運機制等方面的探究[8-9]。光合作用是植物生長的物質基礎,Cd脅迫影響植物葉綠素合成、光合氣體交換、葉綠素熒光特性以及植物對土壤中光合礦質元素的吸收等,從而導致植物代謝紊亂,影響整個生長發育及生命活動[10-11]。促生菌可提高植物的光合作用,促進植物的生長,但由于植物種類和接種菌株的不同,不同植物對促生菌的反映不完全一致。劉美琪等[12研究表明,接種促生菌裸燕麥各時期的凈光合速率(Pn),蒸騰速率(Tr),氣孔導度(Gs),葉片胞間CO2濃度(Ci)均顯著高于對照;袁宗勝等[13]研究顯示,促生菌處理后芳樟葉片光合速率、蒸騰速率和氣孔導度均有所提高,但芳樟葉片胞間CO2濃度降低。促生菌的接種方式可分為單接、混合接。一般認為,單接優于不接,混合接優于單接[14-15]。另外,促生菌對Cd脅迫下植物光合作用的影響研究也已不乏報道[16],但研究多集中于對植物光合色素的探究,缺少對Cd脅迫下植物光合特性的系統性分析,當前關于促生菌對Cd脅迫下植物光合作用的機理尚不甚清楚。
鑒于此,本研究以具促生功能的Bacillussp.T3,PantoeaagglomeransJ2,PseudomonasfluorescensY5的混合菌劑為對象,通過盆栽試驗,系統分析在促生菌作用下蔞蒿光合氣體交換、光合色素、光合礦質元素和葉綠素熒光參數等光合指標對不同濃度Cd脅迫的響應,以揭示促生菌對Cd脅迫下蔞蒿光合特性的作用機理,欲為洞庭湖濕地土壤Cd污染修復建構有效的生物技術提供科學依據。
1 材料與方法
1.1 供試材料
1.1.1促生菌由T3,J2和Y5菌株的菌懸液混合而成,T3菌株為芽孢桿菌(Bacillussp.)分離于蔞蒿的根際土壤,能合成鐵載體、產酸,對Cd2+的耐受濃度為50 mg·L-1[17],J2和Y5菌株分離于蔞蒿的莖葉中,分別為成團泛菌(Pantoeaagglomerans)和熒光假單胞菌(Pseudomonasfluorescens),能分泌吲哚乙酸(IAA),對Cd2+的耐受濃度為90 mg·L-1[18]。供試菌株分別接種于LB液體培養基中,30℃,150 rpm震蕩培養72 h,菌液4℃,8 000 rpm離心 5 min,無菌生理鹽水潤洗2次。為確保各菌株的試驗用量一致,用無菌生理鹽水將各菌液濃度調節至1×108cfu·mL-1,各菌液等量混合制成接種菌劑。
1.1.2基質購自湖南紅星大市場,由草炭、蛭石和珍珠巖混合而成,經測定有機質含量為395 g·kg-1,全P含量為1.15 g·kg-1,全N含量為11.6 g·kg-1,全K含量為10.2 g·kg-1,全Cd含量為0.25 mg·kg-1,pH值為6.85,自然風干且高壓蒸汽滅菌2遍后,添加CdCl2溶液,Cd2+濃度為0 mg·kg-1(Cd0),10 mg·kg-1(Cd10),20 mg·kg-1(Cd20),30 mg·kg-1(Cd30),拌勻平衡備用。
1.1.3蔞蒿取自南洞庭湖管竹山(112°19′33.4″E,28°55′14.9″N),栽種于湖南城市學院苗圃大棚,為保證試驗用苗的一致性,剪取帶3芽的蔞蒿莖段若干進行扦插,正常養護管理,待成活后選擇長勢基本一致的蔞蒿苗用于試驗處理。
1.2 試驗設計
2016年3月10日,將平衡了3周的基質進行裝缽處理。缽大小為:上口徑10.5 cm,底徑7.5 cm,高9.5 cm,稱取基質500 g,與菌劑150 mL充分混勻,然后裝缽,以等量無菌生理鹽水為對照(CK)。選取帶3片完全展開葉且長勢基本一致的蔞蒿扦插苗移栽于營養缽,每處理移栽12缽,澆透水。于苗圃大棚[溫度為(28±2)℃,相對濕度(65±5)%]正常養護管理30天后進行各指標的測定。
1.3 測定方法
1.3.1光合氣體交換參數于2016年4月10日上午9∶00-11∶30進行測定,采用美國Li-Cor公司的Li-6400型光合儀,選取蔞蒿上部第2片完全展開葉進行凈光合速率(Pn),氣孔導度(Gs),蒸騰速率(Tr)和胞間CO2濃度(Ci)的測定,每測定重復3次。測定時光照強度為1 000 μmol·m-2·s-1光量子,氣流速度為50 μmol·s-1,專用內置紅光源,葉室溫度28℃左右。
1.3.2光合色素于2016年4月11日上午采摘蔞蒿上部第3葉于冰盒中帶回實驗室,稱取0.5 g,采用80%的丙酮進行浸提,參考李合生[19]的方法進行葉綠素a,葉綠素b和類胡蘿卜素含量的測定。
1.3.3光合礦質元素每處理隨機選取蔞蒿植物6缽,剪取葉片,去離子水沖冼干凈,置于烘箱中,105℃殺青15 min,70℃下烘干至恒重,研磨粉碎,稱取0.2 g樣品,參照梁泰帥等[8]方法進行消解,用流動注射分析儀測定N,P含量,原子吸收分光光度計測定K,Fe,Zn,Ca,Mg,S,Cd含量。
1.3.4葉綠素熒光參數選取蔞蒿上部第2片完全展開葉,采用德國Walz公司的Imaging PAM葉綠素熒光儀進行測定,測定前暗適應20 min,Kinetics窗口檢測各參數的動力學變化曲線,PSⅡ最大光化學效率(Fv/Fm),實際光化學量子產量(Yield),相對電子傳遞速率(rETR),光化學熒光淬滅系數(qP),非光化學熒光淬滅系數(NPQ)等數據從Report窗口導出。
1.4 統計分析
所有數據均為3次重復的平均值,以平均值±標準差表示,Excel 2010進行數據整理并繪圖,SPSS 26.0進行差異顯著性分析。
2 結果與分析
2.1 促生菌對Cd脅迫下蔞蒿光合氣體交換的影響
Cd脅迫使蔞蒿體內的凈光合速率、氣孔導度以及蒸騰速率均顯著下降(圖1A,1B,1C),而胞間CO2濃度呈先降后升的趨勢,當Cd脅迫濃度為Cd20時降幅顯著(圖1D)。促生菌可緩解Cd脅迫對蔞蒿光合作用的抑制效應,促生菌使Cd脅迫下蔞蒿凈光合速率、氣孔導度及蒸騰速率均有不同程度地增加,其中凈光合速率在所有的Cd脅迫水平下均顯著增加,增幅為13.7%~28.6%(圖1A),氣孔導度在Cd脅迫濃度為Cd30時顯著增加44.1%(圖1B),蒸騰速率在Cd脅迫濃度為Cd0,Cd10,Cd30時顯著增加14.3%,21.4%,39.7%(圖1C)。促生菌使Cd脅迫下蔞蒿細胞間CO2濃度下降,在Cd脅迫濃度為Cd0,Cd10,Cd20時顯著下降7.6%,8.3%,10.1%(圖1D)。

圖1 促生菌對Cd脅迫下蔞蒿光合氣體交換的影響Fig.1 Effects of PGPB on photosynthetic gas exchange of Artemisia selengensis under cadmium stress注:不同小寫字母代表不同處理在 0.05 水平差異顯著(P<0.05)。下圖同Note:Different lowercase letters represent significant differences between treatments at 0.05 level (P<0.05). The same as below
2.2促生菌對Cd脅迫下蔞蒿光合色素的影響
Cd脅迫下蔞蒿葉綠素a,葉綠素b,總葉綠素以及類胡蘿卜素含量均呈先升后降的趨勢,在Cd脅迫濃度為Cd10時達最大值(圖2A,2B,2C,2E),葉綠素a/b和類胡蘿卜素/總葉綠素隨Cd脅迫濃度的增加而增大,在Cd脅迫濃度為Cd30時顯著增加14.6%和26.9%(圖2D,2F)。促生菌有利于Cd脅迫下蔞蒿葉綠素a,葉綠素b,總葉綠素和類胡蘿卜素含量的增加,葉綠素a含量在Cd脅迫濃度為Cd30時顯著增加23.9%(圖2A),葉綠素b含量在Cd脅迫濃度為Cd10~ Cd30時顯著增加,增幅為16.2%~24.6%(圖2B),總葉綠素含量在Cd脅迫濃度為Cd20,Cd30時顯著增加9.6%,23.9%(圖2C),類胡蘿卜素含量在Cd脅迫濃度為Cd30時顯著增加20.0%(圖2E)。促生菌降低了Cd脅迫下蔞蒿葉綠素a/b的值,并呈先降后升的趨勢,在Cd脅迫濃度為Cd20時顯著降低15.7%(圖2D)。促生菌對Cd脅迫下蔞蒿體內類胡蘿卜素/總葉綠素無顯著影響(圖2F)。

圖2 促生菌對Cd脅迫下蔞蒿光合色素的影響Fig.2 Effects of PGPB on photosynthetic pigments of Artemisia selengensis under cadmium stress
2.3 促生菌對Cd脅迫下蔞蒿葉綠素熒光參數的影響
Cd脅迫使蔞蒿體內的最大光化學效率、實際光化學量子產量、相對電子傳遞速率、光化學淬滅系數均呈下降趨勢(圖3A,3B,3C,3D),非光化學淬滅系數隨Cd脅迫濃度的增加而上升(圖3E)。促生菌使Cd脅迫下蔞蒿體內的最大光化學效率、實際光化學量子產量、相對電子傳遞速率、光化學淬滅系系數均顯著增加,非光化學淬滅系數顯著降低,最大光化學效率在Cd脅迫濃度為Cd20,Cd30時顯著增加12.9%,18.0%(圖3A),實際光化學量子產量在Cd脅迫濃度為Cd0,Cd10,Cd30時顯著增加12.5%,18.4%,25.8%(圖3B),相對電子傳遞速率在Cd脅迫濃度為Cd10~ Cd30時顯著增加8.4%~14.6%(圖3C),光化學淬滅系系數在Cd脅迫濃度為Cd10~ Cd30時顯著增加14.3%~20.0%(圖3D),非光化學淬滅系數在在Cd脅迫濃度為Cd10~ Cd30時顯著減少11.1%~53.8%(圖3E)。
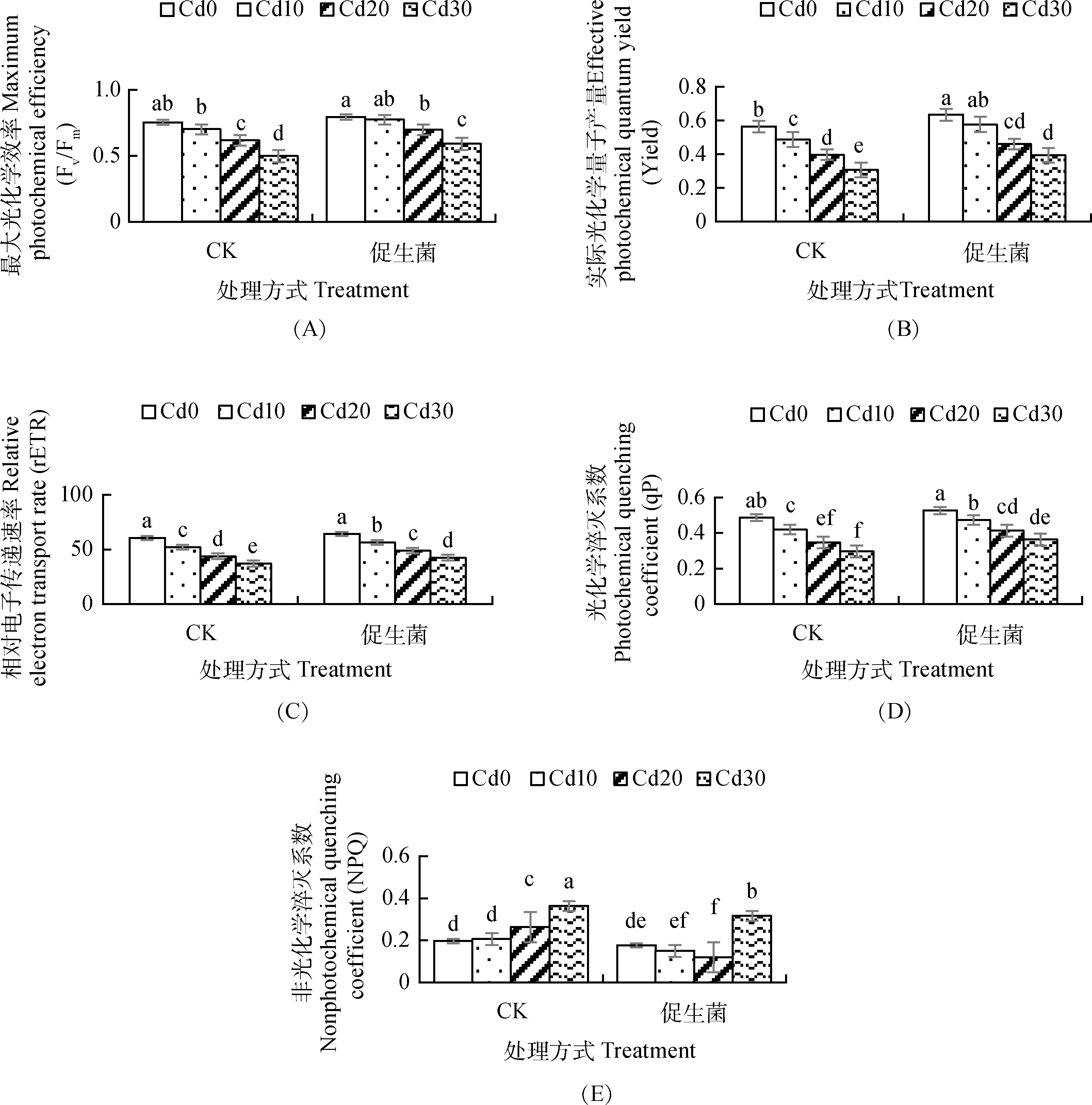
圖3 促生菌對Cd脅迫下蔞蒿葉綠素熒光參數的影響Fig.3 Effects of PGPB on chlorophyll fluorescence parameters of Artemisia selengensis under cadmium stress
2.4 促生菌對Cd脅迫下蔞蒿光合礦質元素的影響
Cd脅迫對蔞蒿N,K的吸收無明顯影響,對S,Cd的影響隨Cd脅迫濃度的增加而增加,對蔞蒿P,Fe,Zn,Ca,Mg的影響隨Cd脅迫濃度的增加呈下降趨勢(表1)。促生菌對Cd脅迫下蔞蒿N,Zn的吸收無顯著影響,促生菌有利于Cd脅迫下蔞蒿對Cd,P,K,Fe,Ca,Mg,S的吸收,P,K分別顯著增加2.2%~4.1%,0.5%~0.7%,Cd在脅迫濃度為Cd10,Cd20,Cd30時顯著增加21.0%,99.3%,79.7%,Fe在Cd脅迫濃度為Cd0,Cd10,Cd20時顯著增加2.8%,3.7%,2.7%,Ca在Cd脅迫濃度為Cd30時顯著增加11.9%,Mg在Cd脅迫濃度為Cd20,Cd30時顯著增加2.7%,5.2%,S在Cd脅迫濃度為Cd30時顯著增加2.5%(表1)。

表1 促生菌對Cd脅迫下蔞蒿光合礦質元素的影響Table1 Effects of PGPB on photosynthetic mineral elements of Artemisia selengensis under cadmium stress 單位:mg·kg-1DW
3 討論
3.1 促生菌對Cd脅迫下蔞蒿光合氣體交換的影響
光合作用是綠色植物最重要的生理活動,為地球上生命的生存和繁衍提供物質和能量[20]。Cd是一種光合抑制劑,Cd脅迫會影響植物體內水分平衡,阻礙氣孔的打開,導致黃瓜凈光合速率和蒸騰速率的下降[21]。無論接菌與否,Cd脅迫下蔞蒿的凈光合速率、蒸騰速率和氣孔導度均顯著低于非Cd脅迫,由此可見,Cd脅迫對蔞蒿光合產生抑制。相關研究表明,接種促生菌可明顯改善裸燕麥的氣孔限制,促進其凈光合速率和蒸騰速率的增加[12]。本研究發現,接種促生菌,蔞蒿的凈光合速率、蒸騰速率和氣孔導度明顯高于單一Cd脅迫,證明接種促生菌緩解了Cd脅迫對蔞蒿光合的限制,這與文獻報道結果一致。但也有不同的報道,Arkhipova等[22]發現接種能分泌細胞分裂素的Bacillussp.IB-22,萵苣的氣孔導度并沒有提高。產生差異的原因可能是促生菌的生理特性不一樣,也可能是試驗材料的不同而異。Cd脅迫導致植物光合傷害的機理可分為氣孔因子和非氣孔因子,非氣孔因子主要表現為葉肉細胞光合活性的降低,胞間CO2濃度的變化可作為氣孔因子和非氣孔因子的評判依據[23]。一般認為,植物凈光合速率、氣孔導度與胞間CO2濃度均下降,則為氣孔因子的限制,如果植物凈光合速率和氣孔導度的下降伴隨胞間CO2濃度的上升,則表現為非氣孔因子的傷害。本研究結果顯示,蔞蒿凈光合速率和氣孔導度隨Cd脅迫濃度的增加而下降,而胞間CO2濃度表現為先降后升,表明Cd脅迫對蔞蒿氣孔的傷害與Cd脅迫濃度有關,濃度較低時主要表現為氣孔限制,而濃度較高時則主要表現為非氣孔限制,這與Cd脅迫龍葵的結果相似[24]。Cd脅迫下蔞蒿葉綠素含量的變化趨勢也證明了這一點(圖2A,2B,2C)。接種促生菌明顯增加蔞蒿凈光合速率和氣孔導度,顯著降低胞間CO2濃度,分析原因可能是由于促生菌的運用增強了蔞蒿葉肉細胞的光合活性,進一步調節了Cd脅迫對蔞蒿氣孔因子與非氣孔因子的影響。
3.2 促生菌對Cd脅迫下蔞蒿光合色素的影響
光合色素可分為葉綠素和類胡蘿卜素,葉綠素是植物光合光能捕獲的關鍵物質,其含量高低在某種程度上可體現植物光合及抗逆性的強弱[25-26]。Cd脅迫通常會導致葉綠素含量的下降,與Cd影響氨基-γ-戊酮酸的合成和抑制原葉綠素酯氧還酶的活性有關,也與Cd破壞葉綠體的結構有關[27]。而本研究中,葉綠素a,葉綠素b,總葉綠素含量呈先升后降的趨勢,表明Cd脅迫對蔞蒿葉綠素含量的影響存在劑量效應,低濃度Cd具有一定的毒物興奮作用。相關研究表明,接種促生菌可提高甘藍型油菜和番茄的抗逆性,保障其葉綠素酶的催化能力,提高其葉綠素含量[28-29]。本研究結果顯示,蔞蒿葉綠素a,葉綠素b和總葉綠素含量在促生菌的作用下明顯增加,表明接種促生菌提高了蔞蒿的抗逆性并改善Cd脅迫對蔞蒿葉綠素酶活性和葉綠體結構的影響,這與文獻報道結果相似;蔞蒿葉綠素a/b值顯著降低,說明葉綠素b比葉綠素a增加得更多,葉綠素b主要存在于光系統的天線組分中,表明接種促生菌蔞蒿葉片吸收和傳遞到光系統反應中心的光能增多;類胡蘿卜素屬于天線色素兼具光保護的功能,可吸收多余光能并以熱能的形式釋放,促生菌作用下蔞蒿類胡蘿卜素含量增加,說明接種促生菌后蔞蒿葉片在吸收較多光能的同時開啟了自保護機制,避免其葉綠體在強光下受到傷害,這與前人的相關研究結論一致[25];但蔞蒿葉綠素a/b值在Cd脅迫濃度為Cd20時顯著減少,而類胡蘿卜素含量是在Cd脅迫濃度為Cd30時顯著增加,兩者變化不完全同步,究其原因可能與促生菌對光能的利用有關,也可能是蔞蒿適應Cd脅迫的一種自我保護。
3.3 促生菌對Cd脅迫下蔞蒿葉綠素熒光參數的影響
葉綠素熒光參數反映植物光合過程中光能的吸收、傳遞、耗散和分配情況[21]。最大光化學效率對環境脅迫比較敏感,可作為植物PSⅡ受傷害程度的判斷指標[27]。正常情況下植物葉片最大光化學效率為0.75~0.85[30]。單一Cd脅迫蔞蒿最大光化學效率均低于0.75,接菌后蔞蒿最大光化學效率有所提高,表明促生菌降低了Cd脅迫對蔞蒿PSⅡ的傷害程度。實際光化學量子產量和相對電子傳遞速率用于反映環境脅迫下植物PSⅡ的光化學活性[31-32]。Cd脅迫下蔞蒿的實際光化學量子產量和相對電子傳遞速率均下降,Cd2+一旦進入葉綠體便會與類囊體膜上的巰基結合導致蛋白變性,會替代放氧復合體中Ca/Mn聚集體上的Ca2+[23],從而降低蔞蒿的PSⅡ的光化學活性。促生菌顯著提高Cd脅迫下蔞蒿的實際光化學量子產量和相對電子傳遞速率,緩解Cd脅迫對蔞蒿PSⅡ光化學的抑制。光化學淬滅系數表示天線色素所吸收光能的光合電子傳遞的情況而非光化學淬滅系數表示光能的熱耗散部分[23,33]。Cd脅迫下蔞蒿光化學淬滅系數下降而非光化學淬滅系數隨Cd脅迫濃度的增加而增加,說明蔞蒿在Cd脅迫下啟動了光保護機制,減少光化學途徑能量的傳遞,避免光氧化脅迫對光合機制的進一步傷害,這與Cd脅迫結縷草的結論相似[27]。接種促生菌,蔞蒿的光化學淬滅系數增加,非光化學淬滅系數減少,表明促生菌提高了蔞蒿對所吸收光能的轉化,更多用于驅動光化學反應,提高蔞蒿對光能的利用效率。
3.4 促生菌對Cd脅迫下蔞蒿光合礦質元素的影響
礦質元素直接或間接影響光合作用,Cd脅迫影響植物對礦質元素的吸收和改變植物體內的化學物質,致使光合的下降。研究表明Cd2+通過取代1,5-二磷酸核酮糖羧化酶(Rubisco)活性中心的Mg2+,改變Rubisco結構,導致其活性的下降,影響CO2的固定[34]。植物體內聚集的Cd2+會替代葉綠素卟啉環中的Mg2+,形成了“Cd替代葉綠素”,影響電子釋放[35]。Cd2+和Ca2+的化學性質極其相似,Cd2+會與Ca2+競爭離子通道和載體蛋白,阻礙Ca2+信號傳遞,影響葉綠素的合成[36]。Cd2+會替代PSII放氧復合體中Ca/Mn聚集體上的Ca2+,影響水的光解和電子釋放[37-38]。Cd2+和Zn2+是一對競爭性陽離子,會競爭相似的活性位點,引起植物的Cd中毒[39]。本研究發現,Cd脅迫下蔞蒿體內Cd含量顯著增加,并且Cd脅迫對蔞蒿Zn,Ca,Mg的吸收產生拮抗作用,這與Cd脅迫黑麥草的試驗結果一致[40]。Cd脅迫與Zn,Ca,Mg產生競爭,可能會形成“Cd替代葉綠素”,可能會影響Rubisco活性,也可能會取代Ca/Mn聚集體上的Ca2+,影響水的光解或與Zn競爭活性位點等,導致蔞蒿光合的下降,但下降的具體原因有待進一步的驗證。Fe參與光合電子傳遞,Cd脅迫導致蔞蒿Fe含量下降,缺Fe會影響光合電子傳遞。但Root等[41]認為,Cd脅迫引起Fe含量上升,且Fe/Zn值增加是Cd脅迫下玉米葉片黃化的主要原因。缺P會影響植物光合同化力和中間磷酸化合物的形成,缺K影響植物光合產物的運輸,S是植物葉綠體的重要組成成分[42]。接種促生菌后蔞蒿體內Ca,Mg,Fe,S,P,K的含量顯著增加,促生菌改善了Cd脅迫下蔞蒿對礦質元素的吸收,緩解了Cd脅迫對蔞蒿光合的抑制,這與促生菌對Cd脅迫下鷹嘴豆礦質元素吸收的影響相似[43]。促生菌之所以能促進Cd脅迫下蔞蒿對礦質元素的吸收,可能是因為①促生菌中的J2,Y5菌株能分泌IAA,促進蔞蒿根系對土壤中礦質元素的吸收;②促生菌中的T3菌株能產生鐵載體且能分泌有機酸,促進蔞蒿對鐵的吸收并降低土壤pH值,改善根際礦質元素的生物有效性。
4 結論
相比單一Cd脅迫,接種促生菌能緩解Cd脅迫對蔞蒿氣孔和非氣孔因子的限制,緩解高濃度Cd脅迫對蔞蒿光合色素的破壞,促進蔞蒿對土壤中光合礦質元素的吸收,減輕Cd脅迫對蔞蒿PSⅡ的傷害,提高蔞蒿對所吸收光能的轉化和利用,保障其光合作用。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
