基于生物信息學(xué)構(gòu)建阿爾茨海默病內(nèi)源競(jìng)爭(zhēng)RNA調(diào)控網(wǎng)絡(luò)
2022-03-11 09:58:32曲浩寧朱文韜何群
中國(guó)醫(yī)科大學(xué)學(xué)報(bào) 2022年2期
曲浩寧,朱文韜,何群
(中國(guó)醫(yī)科大學(xué)生命科學(xué)學(xué)院生物信息學(xué)教研室,沈陽 110122)
我國(guó)阿爾茨海默病(Alzheimer’s disease,AD)患者數(shù)量隨著人口老齡化而不斷增加[1]。AD是一種漸進(jìn)性中樞神經(jīng)退行性疾病,目前認(rèn)為AD的主要致病機(jī)制有β-淀粉樣蛋白(Aβ)生成增加、tau 蛋白沉積、神經(jīng)系統(tǒng)免疫炎癥反應(yīng)、氧化應(yīng)激損傷、糖代謝異常等[2-3]。長(zhǎng)鏈非編碼RNA(long non-coding RNA,lncRNA)在神經(jīng)退行性疾病中有重要作用,并可能通過競(jìng)爭(zhēng)性結(jié)合微RNA(microRNA,miRNA)形成復(fù)雜的內(nèi)源競(jìng)爭(zhēng)RNA(competing endogenous RNA,ceRNA)調(diào)控網(wǎng)絡(luò),從而引發(fā)神經(jīng)退行性疾病[4]。
lncRNA是一組長(zhǎng)度不小于200個(gè)核苷酸、沒有明顯編碼蛋白潛能的非編碼RNA(non-coding RNA,ncRNA)[1]。研究[2-4]表明,lncRNA在轉(zhuǎn)錄和轉(zhuǎn)錄后水平參與基因表達(dá)的調(diào)控,并具有明顯的組織特異性、細(xì)胞特異性、時(shí)空特異性和發(fā)育階段特異性,其變化和發(fā)育也與疾病狀態(tài)有關(guān),故在多種疾病的進(jìn)展中起著關(guān)鍵作用。lncRNA有多種功能,細(xì)胞質(zhì)中的lncRNA可以誘捕miRNA與之結(jié)合,通過ceRNA機(jī)制競(jìng)爭(zhēng)性地結(jié)合miRNA來調(diào)節(jié)基因表達(dá)。[5]
1 材料與方法
1.1 篩選差異表達(dá)靶基因
從基因表達(dá)綜合數(shù)據(jù)庫(gene expression omnibus,GEO)(https://www.ncbi.nlm.nih.gov/geo/)中下載數(shù)據(jù)GSE28146,使用GEO數(shù)據(jù)庫GEO2R以P< 0.05為條件分別篩選AD輕度組、中度組和重度組與正常對(duì)照組比較的差異基因(differentially expressed genes,DEGs),AD輕度組篩選出DEGs 2 612個(gè),AD中度組篩選出DEGs 3 120個(gè),AD重度組篩選出DEGs 3 187個(gè)。應(yīng)用Venny 2.1.0 [Venny 2.1.0(csic.es)],取輕、中、重度組DEGs的交集為候選差異基因(candidate differentially expressed genes,pDEGs),在pDEGs中 篩選差異表達(dá)的lncRNA進(jìn)行后續(xù)分析。
1.2 lncRNA靶基因的預(yù)測(cè)及富集分析
利用在線分析工具RegRNA(http://regrna2.mbc.nctu.edu.tw/)預(yù)測(cè)lncRNA的靶基因。RegRNA是專門用來進(jìn)行RNA功能性的motif預(yù)測(cè)網(wǎng)站,該網(wǎng)站預(yù)測(cè)內(nèi)容包含轉(zhuǎn)錄motif、mRNA降解原件、RNA-RNA結(jié)合、翻譯預(yù)測(cè)等功能。
1.3 預(yù)測(cè)lncRNA的亞細(xì)胞定位
應(yīng) 用lnclocater(http://www.csbio.sjtu.edu.cn/bioinf/lncLocator/)和iLoc-mRNA(http://lin-group.cn/server/iLoc-mRNA/predictor.php)對(duì)lncRNA的亞細(xì)胞定位進(jìn)行預(yù)測(cè),找到lncRNA具體存在的細(xì)胞位置。
1.4 miRNA靶基因的預(yù)測(cè)
應(yīng)用miRDB(https:/www.mirdb.org/)、TargetScan(https://www.targetscan.org/)和TargetMiner(https://www.isical.ac.in/~bioinfo_miu/final_html_targetminer/hsamiR-1254.html)3個(gè)在線數(shù)據(jù)庫預(yù)測(cè)mRNA的靶基因,并用Venny 2.1.0繪制韋恩圖,將此3個(gè)數(shù)據(jù)庫得到的靶基因與GSE28146數(shù)據(jù)集中輕、中、重度3組共有的表達(dá)上調(diào)的pDEGs取交集作為最后miRNA靶基因集合。將靶向基因集提交DAVID(https://david.ncifcrf.gov/tools.jsp)網(wǎng)站以P< 0.05為條件進(jìn)行基因本體(gene ontology,GO)分析和PATHWAY分析。
1.5 ceRNA網(wǎng)絡(luò)的構(gòu)建及其富集分析
將靶向基因集提交DAVID(https://david.ncifcrf.gov/tools.jsp)網(wǎng)站以P< 0.05為條件進(jìn)行GO富集分析分析和PATHWAY分析。使用Cytoscape(3.8.0版)繪制ceRNA網(wǎng)絡(luò),構(gòu)建GSE28146數(shù)據(jù)集中pDEGs得蛋白質(zhì)相互作用(protein-protein interaction,PPI)網(wǎng)絡(luò)并用Cytoscape找到其關(guān)鍵基因,與miRNA靶基因集合做交集,以確定ceRNA網(wǎng)絡(luò)中的lncRNA在疾病中是否有關(guān)鍵作用,調(diào)控關(guān)鍵蛋白。
2 結(jié)果
2.1 共有差異lncRNA的篩選結(jié)果
AD輕度組篩選出DEGs 2 612個(gè),中度組篩選出DEGs 3 120個(gè),重度組篩選出DEGs 3 187個(gè)。為了篩選在AD中其關(guān)鍵作用的基因,取早中晚3組與正常相比異常的252個(gè)DEGs,然后從數(shù)據(jù)集GSE28146中早中晚差異表達(dá)的pDEGs中篩選出LINC02047、LINC01124、LINC00582和LINC02478,均為表達(dá)上調(diào)基因。
2.2 lncRNA的亞細(xì)胞定位
為了分析這些差異lncRNA是否參與ceRNA競(jìng)爭(zhēng)機(jī)制,利用lnclocater和iLoc-mRNA網(wǎng)站對(duì)其亞細(xì)胞定位進(jìn)行分析,結(jié)果顯示,LINC02047、LINC01124和LINC02478定位于細(xì)胞質(zhì),見表1。

表1 亞細(xì)胞定位Tab.1 Data of subcellular localization
2.3 lncRNA靶基因的預(yù)測(cè)結(jié)果
RegRNA預(yù)測(cè)LINC02047的靶基因?yàn)閔sa-miR-132-3p,LINC011124的靶基因有4個(gè),分別為hsa-miR-1254、hsa-miR-4640-5p、hsa-miR-4690-3p和hsa-miR-4786-3P,LINC02478的靶基因?yàn)閔sa-miR-3612。
2.4 miRNA靶基因的預(yù)測(cè)結(jié)果
使用數(shù)據(jù)庫miRDB、TargetMiner和TargetScan分別預(yù)測(cè)hsa-mir-132-3p、hsa-mir-1254、hsa-mir-3612、hsa-mir-4640-5p、hsa-mir-4690-3p、hsa-mir-4786-3p的靶基因。其中hsa-mir-132-3p用在線韋恩圖工具Venny 2.1.0可得到靶基因交集126個(gè),占總數(shù)4.4%。hsa-mir-1254靶基因交集722個(gè),占總數(shù)11.6%。hsamir-3612靶基因交集290個(gè),占總數(shù)3.3%。hsa-mir-4640-5p靶基因交集231個(gè),占總數(shù)的3.3%(圖1)。hsa-mir-4690-3p靶基因交集126個(gè),占總數(shù)2.2%。hsa-mir-4786-3p靶基因交集234個(gè),占總數(shù)的3.4%。再將3個(gè)數(shù)據(jù)庫預(yù)測(cè)到的靶基因交集在與早中晚3組共有表達(dá)上調(diào)的pDEGs取交集,見表2。GO富集分析及京都基因與基因組百科全書(Kyoto encyclopedia of genes,KEGG)通路分析結(jié)果見表3、4。
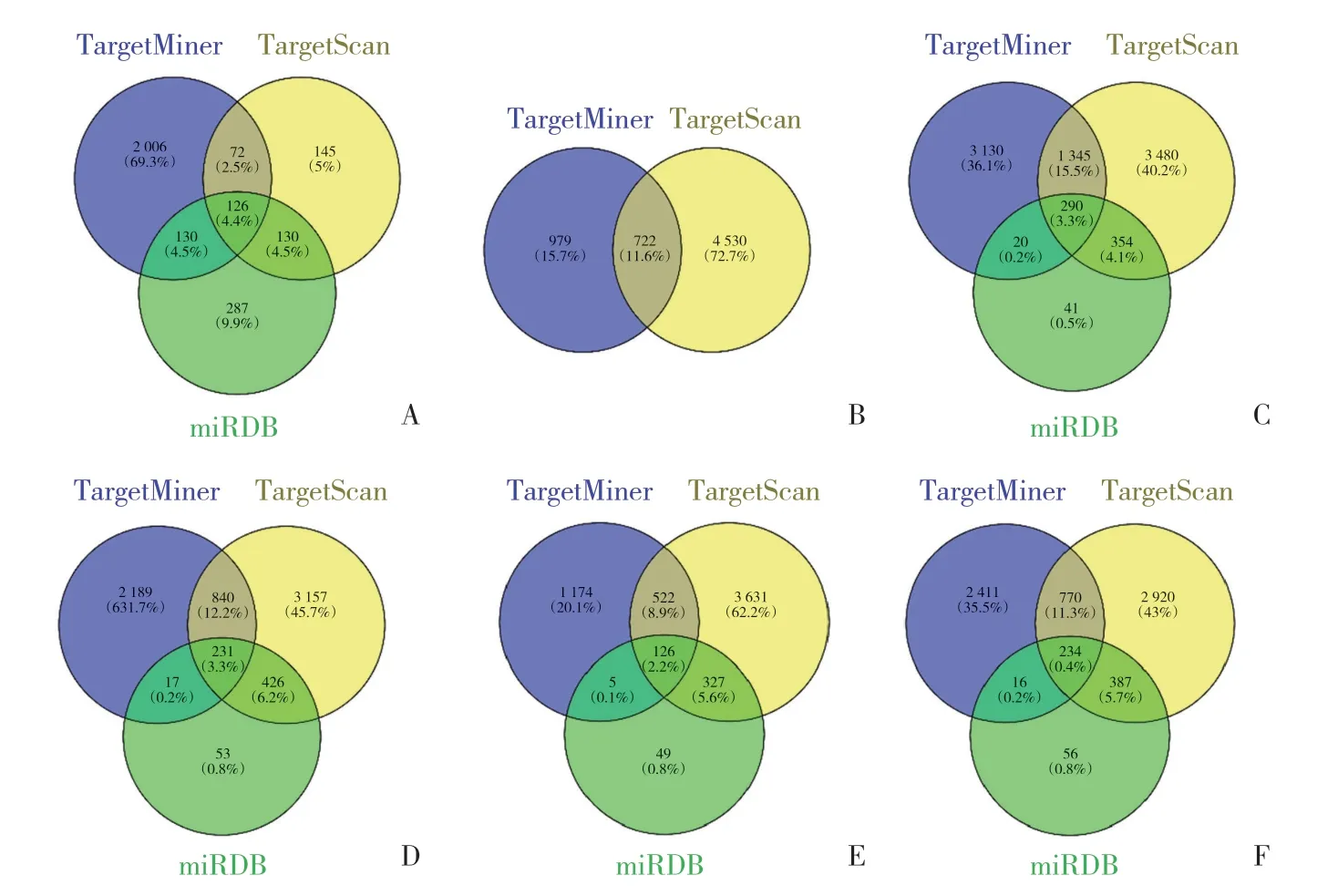
圖1 miRNA靶基因預(yù)測(cè)結(jié)果Fig.1 Prediction results of miRNA target genes

表2 miRNA靶基因預(yù)測(cè)結(jié)果與早中晚3組pDEGs的交集Tab.2 Intersection of prediction results of miRNA target genes and early,middle,and late differentially expressed genes

表3 miRNA靶基因GO富集分析結(jié)果Tab.3 Gene ontology enrichment analysis results of miRNA target genes

表4 miRNA靶基因KEGG通路分析結(jié)果Tab.4 KEGG pathway analysis results of miRNA target genes
2.5 構(gòu)建ceRNA網(wǎng)絡(luò)
將所得lncRNA、miRNA與mRNA在Cytoscape3.8.0中繪制出ceRNA網(wǎng)絡(luò),共21條通路,30個(gè)節(jié)點(diǎn),27條邊。見圖2。構(gòu)建PPI網(wǎng)絡(luò)后經(jīng)Cytoscape hubba分析得分較高的基因與miRNA交集后得到了4個(gè)得分較高的關(guān)鍵基因,分別為NMNAT2、DLGAP2、SVOP和UBE3A,得分分別為20 541、17 936、12 362和95。
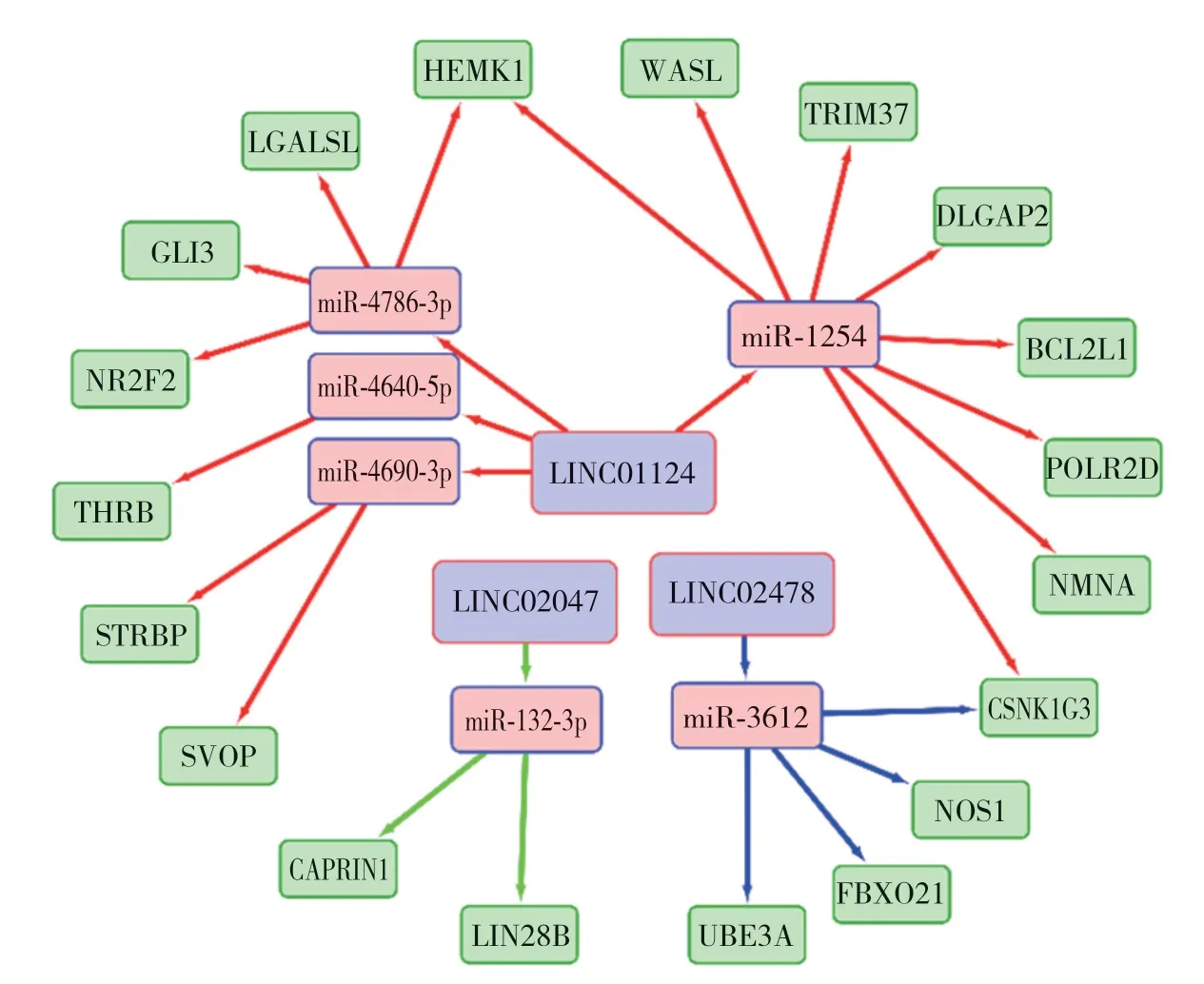
圖2 lncRNA、miRNA與mRNA 組成的ceRNA網(wǎng)絡(luò)Fig.2 Competing endogenous RNA network composed of lncRNA,miRNA,and mRNA
3 討論
AD是全球癡呆最常見的類型,全球約有4 000萬人患有AD,并且患病率逐年上升[7-8]。lncRNA的DEGs已被證實(shí)與神經(jīng)退行性疾病的發(fā)生發(fā)展有密切關(guān)系[9],但尚無在AD調(diào)控網(wǎng)絡(luò)作用機(jī)制中針對(duì)lncRNA的差異表達(dá)研究。
為了尋找與AD相關(guān)的關(guān)鍵lncRNA,本研究選取了GEO數(shù)據(jù)庫GSE28146,分析了其正常組織與各個(gè)時(shí)期AD病例組織中DEGs,篩選差異表達(dá)的lncRNA。其中,LINC02047、LINC01124和LINC02478在AD早期、中期和晚期的病理組織中均存在差異。為進(jìn)一步探討上述3種lncRNA在AD發(fā)病機(jī)制中起到的關(guān)鍵作用,本研究通過亞細(xì)胞定位確定了這3個(gè)lncRNA在細(xì)胞質(zhì)中的表達(dá),表明上述lncRNA可通過ceRNA機(jī)制調(diào)控基因表達(dá)參與AD發(fā)病。
RegRNA預(yù)測(cè)結(jié)果顯示,lncRNA通過調(diào)控miRNA而影響神經(jīng)退行性病變中的基因表達(dá)。LINC01124具有抑制細(xì)胞增殖、遷移和侵襲的能力[10];miR-132被證實(shí)是AD神經(jīng)元中最顯著下調(diào)的miRNA,miR-132可促進(jìn)神經(jīng)突起的延長(zhǎng)和分支,減少神經(jīng)元死亡[11];miR-1254通過與靶區(qū)的非結(jié)構(gòu)位點(diǎn)結(jié)合也可以起到阻遏作用[12],以上結(jié)果揭示了ceRNA的部分調(diào)控關(guān)系,然而對(duì)于LINC01124,LINC02478,miR-4060,miR-4090,miR-4786,miR-3612的分子機(jī)制及其如何作用于神經(jīng)退行性病變,迄今尚無研究闡明。
基于lncRNA-miRNA-mRNA調(diào)控關(guān)系,本研究對(duì)各項(xiàng)功能節(jié)點(diǎn)的可信度進(jìn)行了篩選,并對(duì)ceRNA的調(diào)控過程構(gòu)建了可視化網(wǎng)絡(luò)。基于David對(duì)ceRNA網(wǎng)絡(luò)進(jìn)行分析的結(jié)果,基因組整體展現(xiàn)了抑制細(xì)胞增殖、遷移和侵襲與類固醇激素受體的功能。
KEGG通路分析提示其參與Hedgehog信號(hào)通路。Hedgehog信號(hào)通路在人胚胎干細(xì)胞神經(jīng)分化的d6~d14持續(xù)活化,而Sonic Hedgehog作為Hedgehog信號(hào)通路家族成員之一,對(duì)基底前腦膽堿能神經(jīng)元(basal forebrain cholinergic neurons,BFCNs)的發(fā)育分化起到至關(guān)重要的作用[13]。BFCNs投射系統(tǒng)的退行性病變是導(dǎo)致AD患者出現(xiàn)空間認(rèn)知功能障礙的主要原因[14]。樞紐基因NMNAT2的缺失可能會(huì)導(dǎo)致自發(fā)性的軸突變性,使軸突運(yùn)輸受阻,進(jìn)而引發(fā)AD[15-17];Dlgap2和AD表型在變異、基因和蛋白質(zhì)表達(dá)以及甲基化水平上存在關(guān)聯(lián)[18];Ube3A的缺失觸發(fā)Arc和Ephexin-5的積累,分別驅(qū)動(dòng)GluR1的內(nèi)化和RhoA的激活,最終導(dǎo)致突觸的剪斷被Ube3A的恢復(fù)所阻斷[19];神經(jīng)元表達(dá)轉(zhuǎn)運(yùn)蛋白SV2相關(guān)蛋白(SVOP和SVOPL)在調(diào)節(jié)胞吐中起到關(guān)鍵作用[20]。
本研究利用生物信息學(xué)軟件挖掘了各時(shí)期AD中共差異表達(dá)的lncRNA,并繪制了其與靶向miRNA、mRNA的ceRNA網(wǎng)絡(luò)圖。結(jié)果驗(yàn)證了部分已被闡明的AD作用機(jī)制,同時(shí)也發(fā)現(xiàn)了尚未得到充分研究且可能成為潛在藥物靶點(diǎn)的重要lncRNA與miRNA。本研究基于臨床樣本數(shù)據(jù),探尋各期AD存在的共同差異,構(gòu)建了完整的ceRNA調(diào)控網(wǎng)絡(luò),為AD分子機(jī)制的深入研究提供了嚴(yán)謹(jǐn)?shù)臄?shù)據(jù)支持。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級(jí)語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
電子制作(2018年18期)2018-11-14 01:48:24
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
財(cái)經(jīng)(2017年2期)2017-03-10 14:35:35
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
財(cái)經(jīng)(2016年15期)2016-06-03 07:38:02
財(cái)經(jīng)(2016年3期)2016-03-07 07:44:46
財(cái)經(jīng)(2016年6期)2016-02-24 07:41:51
