兩份新株型水稻品系的農藝性狀與遺傳特性分析
2022-03-17 12:09:00梁程向珣朝張歐玲游慧許亮陳永軍
中國水稻科學 2022年2期
梁程 向珣朝 張歐玲 游慧 許亮 陳永軍
兩份新株型水稻品系的農藝性狀與遺傳特性分析
梁程 向珣朝*張歐玲 游慧 許亮 陳永軍
(西南科技大學 植物分子遺傳育種實驗室,四川 綿陽 621010;*通信聯系人, E-mail: xxc@swust.edu.cn)
【】理想株型水稻能提高光合作用效率、經濟產量和抗病能力。通過探究兩份新株型水稻種質的農藝性狀變異和遺傳特性,為水稻高產抗病育種提供優異理想株型種質資源。【】以兩個重組自交系中發現的兩份自然重組的新株型種質08yi和RIL60以及由同一自交系選育出來的常規種質08yc和RILc為材料,進行不同生長時期的株型構成因子和產量性狀的比較分析,同時對4份種質進行理想株型基因測序和糯性鑒定。【】兩份新種質與常規種質相比,在不同發育時期的功能葉性狀、干物質積累量、穗部性狀和產量構成因素上均存在顯著變異,二者發生的有利變異包括功能葉葉角減小、功能葉葉長更合理、株型緊湊、單株有效穗數增加1~2穗,結實率分別提高了7.37%和5.09%;此外,新種質08yi葉綠素含量顯著提高,RIL60的莖稈變粗,二者在逆境下的穗部性狀表現更好,產量降幅更小,抗逆力增強。不利變異包括單穗重降低、穗長變短、每穗著粒數和每穗實粒數減少、枝梗長度和數量下降。基因測序結果顯示與少蘗粳相比,兩新種質并未突變株型變異源自于其他因子。糯性鑒定結果表明RIL60存在糯性變異。【】兩個新種質的一些農藝性狀發生了明確的有利變異,具有較理想的株型結構,可作為優異種質用于理想株型育種和株型研究。RIL60作為理想株型糯稻,由于具有非糯水稻的經濟產量,具有巨大的應用價值。
水稻(L.);理想株型;農藝性狀;遺傳特性;基因
水稻(L.)是我國主要的糧食作物,在經歷了矮化育種、三系雜交稻選育、兩系雜交稻選育、亞種間雜種優勢利用、理想株型育種和綠色超級稻選育等6個重要歷程后,水稻單產已實現巨大突破[1-2]。在育種過程中,通過水稻株型相關基因定位、克隆及有利基因聚合,有效結合分子技術,培育出具有理想株型的水稻品種,是提高水稻產量的重要舉措[3]。水稻株型能極大影響水稻葉片與穗的空間分布、光合作用速率、干物質量積累、抗逆性及生物量;建立理想的水稻株型,提高作物產量,是進行水稻超高產育種和應對現代農業發展挑戰的重要方式[4]。水稻理想株型概念最早由Donald提出,理想株型植株能最大限度地利用光能從而提高經濟系數及作物產量[5]。高產水稻株型模式包括“半矮稈叢生早長”模式、“重穗型”模式、“少蘗大穗”模式、“直立葉、直穗大穗”模式和“超高產水稻株型”模式等[6-7]。這些株型模式提高了水稻在相應生長環境下的產量,同時對株型改良和理想株型育種具有重要的指導意義。
理想株型育種是通過改變株高、葉型、分蘗數量和穗部形態等方式來改良水稻的空間構型,使其能夠最大限度地利用光能,積累更多生物量,提高結實率,從而增加經濟系數[8]。理想株型構成因子主要包括水稻根、莖、葉和穗。其一般特點是根系發達,活力旺盛;莖稈粗壯,株高適中;分蘗能力強,無效分蘗少;葉片厚直、色深,葉夾角小;穗大粒多,結實飽滿[9-10]。利用分子手段聚合株型有利基因和從雜交育種中挖掘新的種質資源是水稻理想株型育種的重要途徑。位于水稻第8染色體上的半顯性基因能調控水稻株型,該基因編碼的啟動子結合蛋白OsSPL14激活了產量相關的基因,包括密集直立穗基因,使植株具有更少的無效分蘗和更多的穗粒,同時在稻瘟病菌侵染時誘導Ser163磷酸化,激活與免疫相關的基因,實現了增產和提高水稻稻瘟病抗性的雙重效用[11-13]。當位于該基因第3外顯子上的第874位點的堿基C突變成A時,釋放miR156和miR529的靶向抑制,導致IPA1 RNA和蛋白水平升高,使水稻具有了理想的株型結構,產量增加了10%~13%[11, 13]。水稻第6染色體上的基因調控胚乳直鏈淀粉含量(amylose content, AC),當其第2外顯子發生23 bp插入突變時不能正常編碼顆粒結合淀粉合成酶(granule binding starch synthase, GBSS),進而無法正常合成直鏈淀粉,水稻發生糯性變異并伴隨粒重降低,產量下降[14-15]。西南科技大學植物分子遺傳育種實驗室在構建的兩個重組自交系中發現兩份自然重組的新種質,其具有理想株型特點。本研究對其與相應常規種質進行不同生長時期的株型構成因子、產量因子性狀考查,同時與含有的少蘗粳一起進行的遺傳差異分析以及糯性鑒定,旨在對新種質做出正確評價,為理想株型育種挖掘優異種質資源和提供理論依據。
1 材料與方法
1.1 供試材料與種植方法
試驗分別于2019年和2020年進行,2019年氣候和開花灌漿期的降雨量正常;2020年氣候異常,開花灌漿期的降雨量太多,從開花期開始連續長時間暴雨,在蠟熟期倒2葉以下被水淹3 d,嚴重影響水稻灌漿。以4份水稻種質08yi、08yc、RIL60和RILc為供試材料,其中08yi與RIL60為西南科技大學植物分子遺傳育種實驗室選育的新株型種質,08yc和RILc分別為同一重組自交系同一株系中分離出的與對應新株型株高和生育期相近而株型差異較大的常規株型種質。試驗材料于每年4月上旬播種,水育秧,秧齡30 d,單株栽插。大田肥料用復合肥,折合純N 150 kg/hm2,P2O575 kg/hm2,K2O 195 kg/hm2,全部作底肥施用。劃廂栽插,廂寬1.66 m,廂溝0.33 m,試驗材料置于田中央,減小試驗誤差。每個材料栽12行,栽插規格按行穴距27 cm × 0.16 cm進行,無重復。按常規方法管理。測試取樣時,在其小區中間進行,避免邊際效應。
1.2 農藝性狀測定
在4份種質的3個主要生育期(乳熟期、蠟熟期和完熟期)分別在小區中間隨機取樣3株,進行農藝性狀的測定。08yi和08yc生育期相同,為135 d,RIL60與RILc生育期相同,為145 d。
1.2.1 功能葉性狀測定
分別在乳熟期、蠟熟期和完熟期取樣,測定每株所有有效分蘗功能葉的葉長、葉寬和葉角。分別計算每份種質各功能葉在3個時期的葉面積():=葉長×葉寬×0.75[16]。
1.2.2 葉綠素含量測定
在乳熟期和蠟熟期,分別對4份種質取功能葉中倒2葉和倒3葉去掉葉脈后全部剪碎混勻,然后稱取100 mg,加少量石英砂和少量碳酸鈣粉末,7 mL 95%乙醇研磨呈勻漿狀,倒入帶塞試管中,置于60℃~65℃的溫箱中保溫浸提,葉色變白后取出過濾至棕色瓶,將濾紙洗凈,在暗處貯存或稀釋,以95%乙醇為空白,分別在665、649和470 nm下測定吸光度(optical density, OD)值,計算葉綠素含量[17],生物學重復3次。
1.2.3 干物質量測定
于主要生育期(乳熟期、蠟熟期和完熟期)分別對4份種質取樣3株,對每株的莖、葉、穗進行分割整理,并存放在帶有標簽的紙袋內,105℃下殺青1 h,60℃下烘干1周至恒重。冷卻到室溫后用天平分別對莖、葉和穗稱重。
1.2.4 穗部性狀測定
在完熟期分別對4份種質取樣3株,對每株有效穗、單穗重、穗長、一次枝梗數、一次枝梗總長和二次枝梗數等性狀進行測定。
1.2.5 產量及其構成因子測定
在完熟期分別對4份種質取樣3株,進行株高、著粒數、實粒數和千粒重的測定。并結合穗部性狀數據計算出單株產量、結實率和著粒密度。考種性狀統計標準參照《水稻品種實驗田間記載及室內考種項目與標準》[18]。
1.3 遺傳差異分析
1.3.1基因測序分析
在水稻分蘗期分別取4份種質的新鮮葉片,用微量十二烷基硫酸鈉(sodium dodecyl sulfate, SDS) 法提取基因組DNA[19],以擴增引物[11]進行PCR擴增,反應體系為50 μL,以2%瓊脂糖凝膠電泳鑒定擴增條帶,用純化試劑盒進行PCR產物純化,用Oligo6設計測序引物(表1),一并送往華大基因進行測序。
1.3.2 糯性遺傳特性鑒定
利用田志喜等[20]開發的分子標記Wx M1對4份種質進行糯性遺傳分析。
1.4 數據分析和統計
采用Office 2016進行數據統計,采用Excel繪制圖表,采用IBM SPSS statistics軟件對數據進行差異顯著性分析及多重比較。用DNAMAN軟件進行DNA測序結果比對。
2 結果與分析
2.1 葉部性狀
2019年,4份水稻新種質的葉部性狀考查數據及差異統計分析(-檢驗)見表2和圖1。在考查的3個時期,08yc的劍葉、倒2葉和倒3葉葉長均顯著長于08yi,其中劍葉長變幅最顯著(<0.05),08yi在3個時期的劍葉長較08yc分別降低了28.59%、44.14%和66.03%。RIL60和RILc的功能葉葉長在乳熟期無顯著差異,在蠟熟期和完熟期,RILc功能葉的葉長均較RIL60長。同一時期新種質08yi和RIL60的葉寬與其常規種質無顯著性差異。就葉角度而言,08yc和08yi在3個時期的劍葉葉角度無顯著性差異,08yi在乳熟期的倒2葉葉角度顯著大于08yc(<0.05),但在蠟熟期和完熟期其倒2葉和倒3葉葉角均小于08yc。在乳熟期,RIL60的劍葉葉角較RILc增大了49.63%,但倒3葉葉角卻顯著降低了24.02%。在蠟熟期和完熟期,RILc劍葉的葉角均顯著大于RIL60(<0.05),其差異分別為44.52%和41.48%,但兩者間的倒2葉和倒3葉葉角并無顯著性差異。從乳熟期到完熟期各種質的功能葉葉角的動態變化看,08yi倒2葉不斷下降,而劍葉和倒3葉變化不大;RIL60劍葉不斷減小,倒2葉不斷增加,倒3葉變化不大;08yc倒2葉和倒3葉不斷增加,而劍葉變化不大;RILc劍葉和倒2葉不斷增加,而倒3葉不斷減小。08yi功能葉葉面積在3個時期均較08yc顯著減少(<0.05),其單葉平均面積分別比08yc減少了17.22、14.12和26.52 cm2;RIL60在3個時期的單葉平均面積比RILc分別降低了11.21%、10.62%和13.07%(圖1),RILc功能葉葉面積總體水平上顯著大于RIL60。
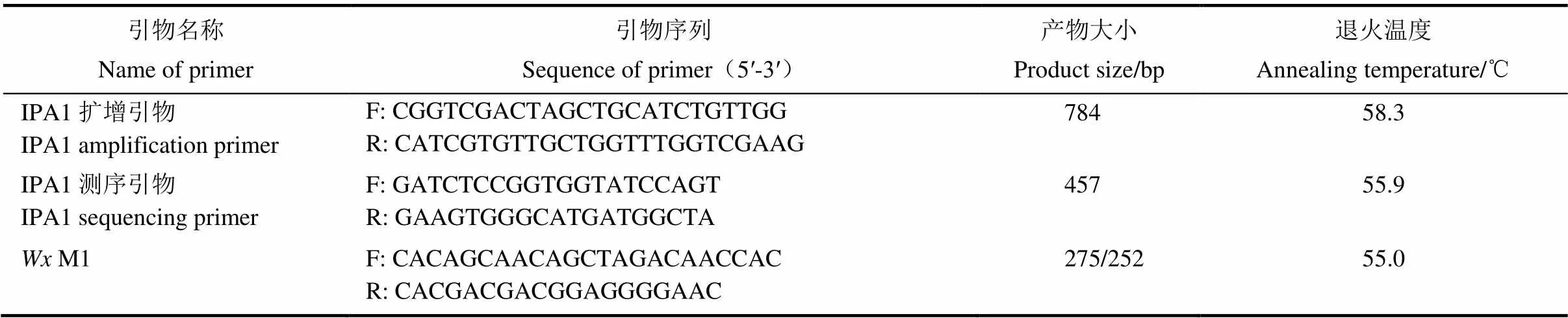
表1 引物信息

A?乳熟期;B?蠟熟期;C?完熟期。均值±標準差,不同小寫字母表示不同種質差異顯著(P<0.05, n=4)(F檢驗)。
Fig. 1. Comparison of functional leaves area for different germplasms at different developmental stages.
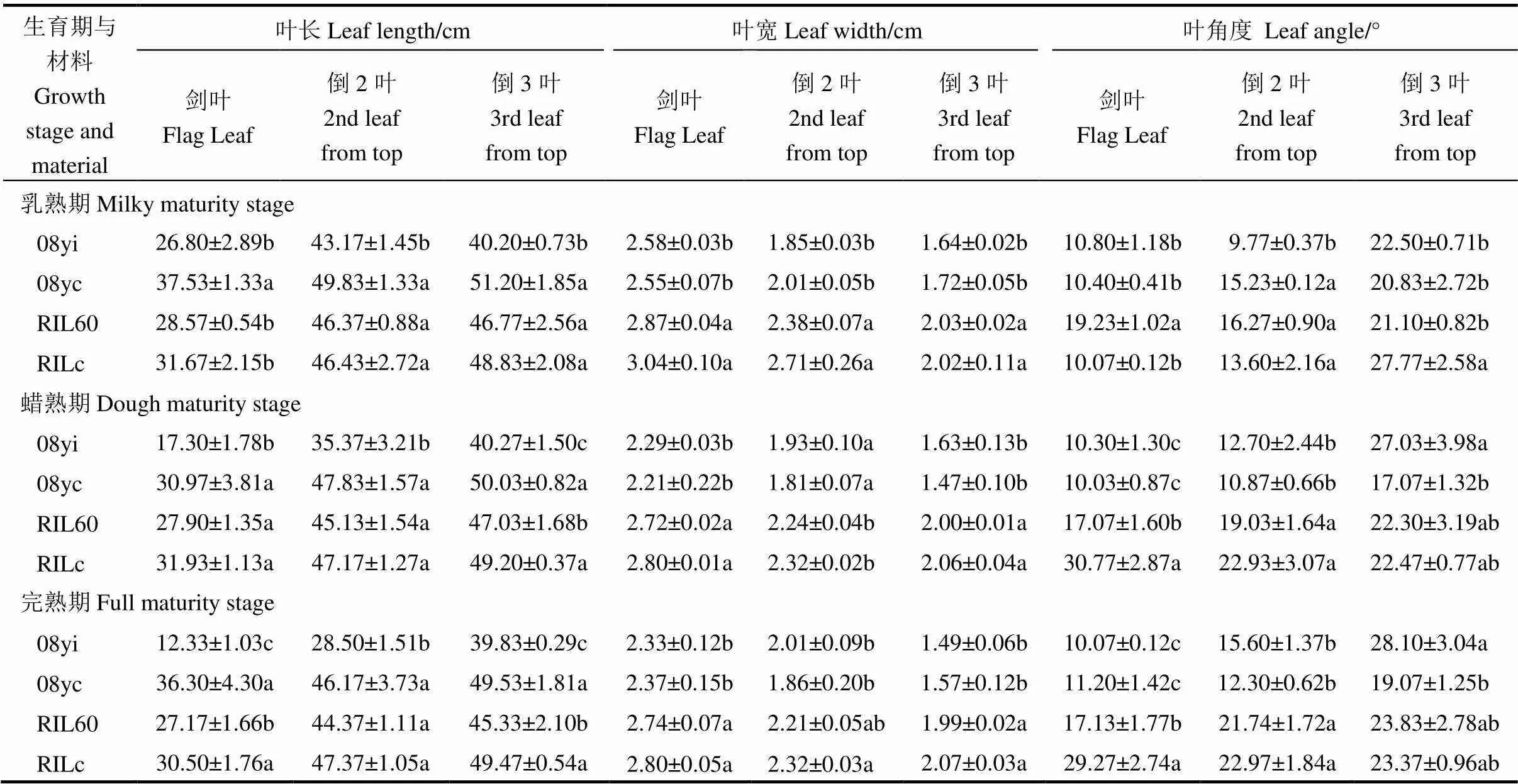
表2 不同種質的功能葉葉長、葉寬和葉角比較
均值±標準差(=4),同一列同一時期中不同小寫字母表示不同種質差異顯著(<0.05,-檢驗)。
Mean±(=4).Different letters in the same column and in the same period are significantly different at<0.05 by-test.
因此,在兩份新株型種質灌漿過程中,從乳熟期到完熟期,劍葉和倒2葉葉角最顯著的特點是均表現減小的趨勢,而常規種質則呈現出不斷增加的趨勢。此外,這兩份新種質較常規種質均存在葉長變短,葉寬無顯著變化,葉角總體減小,葉面積減小的現象。
2.2 葉綠素含量
2019年在乳熟期和蠟熟期,新種質08yi的葉綠素a、葉綠素b和葉綠素總量均顯著高于08yc;在乳熟期(圖2-A),RILc與RIL60的葉綠素b含量無顯著差異,但其葉綠素a含量和葉綠素總含量均顯著高于RIL60。在蠟熟期(圖2-B),RILc的葉綠素a、b及總量均顯著高于RIL60。新株型08yi的葉綠素含量較08yc在乳熟期和蠟熟期分別增加了29.47%和23.05%,但新株型RIL60較RILc在這兩個時期則分別降低了26.36%和21.37%。
2.3 干物質量
在2019年的乳熟期、蠟熟期和完熟期,4份種質的干物質量分布和差異顯著性分析(-檢驗)如圖3所示。08yc在3個時期的葉和莖干質量都顯著高于08yi;08yc的穗干質量在乳熟期和蠟熟期均顯著高于08yi(圖3-A~B),但在完熟期時反而下降,而08yi穗干質量穩定上升(圖3-C),最終兩株型的穗干質量差異并不顯著。RIL60與RILc在乳熟期的穗和葉干質量并無顯著性差異(圖3-A),而RIL60莖干質量比RILc增加了56.56%;在蠟熟期(圖3-B),RILc穗干質量顯著高于RIL60,兩種質間的葉干質量無顯著差異,RIL60莖干質量仍顯著高于RILc(<0.05)。在完熟期時(圖3-C),RILc在穗、葉和莖干質量均顯著高于RIL60。
說明新種質08yi的干物質積累能力優勢主要表現在生育期后期,而RIL60主要集中在前中期,在完熟期比RILc表現差。
2.4 產量及其構成因素
2.4.1 穗部性狀
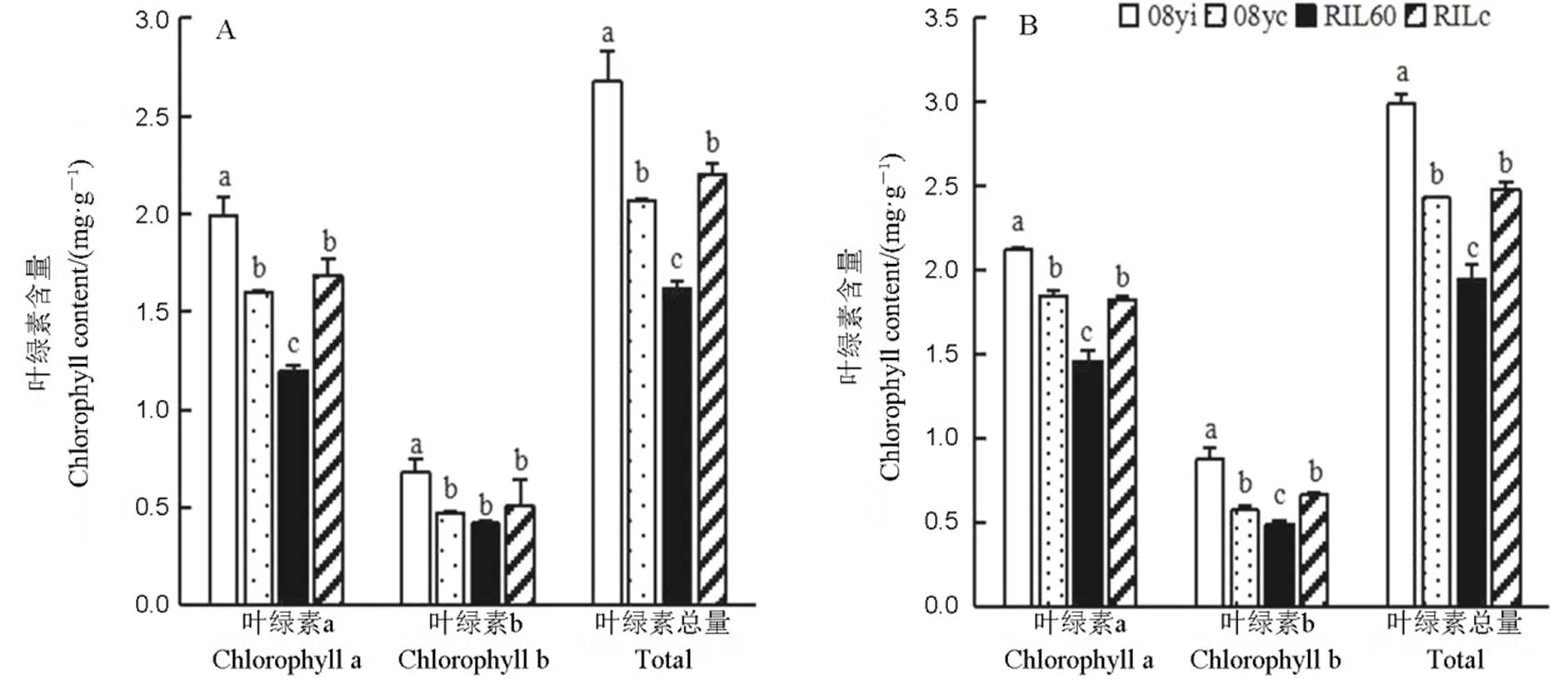
A?乳熟期;B?蠟熟期。均值±標準差,不同小寫字母表示不同種質差異顯著(P<0.05, n=4)(F檢驗)。
Fig. 2. Comparison of chlorophyll contents for different germplasms at different developmental stages.
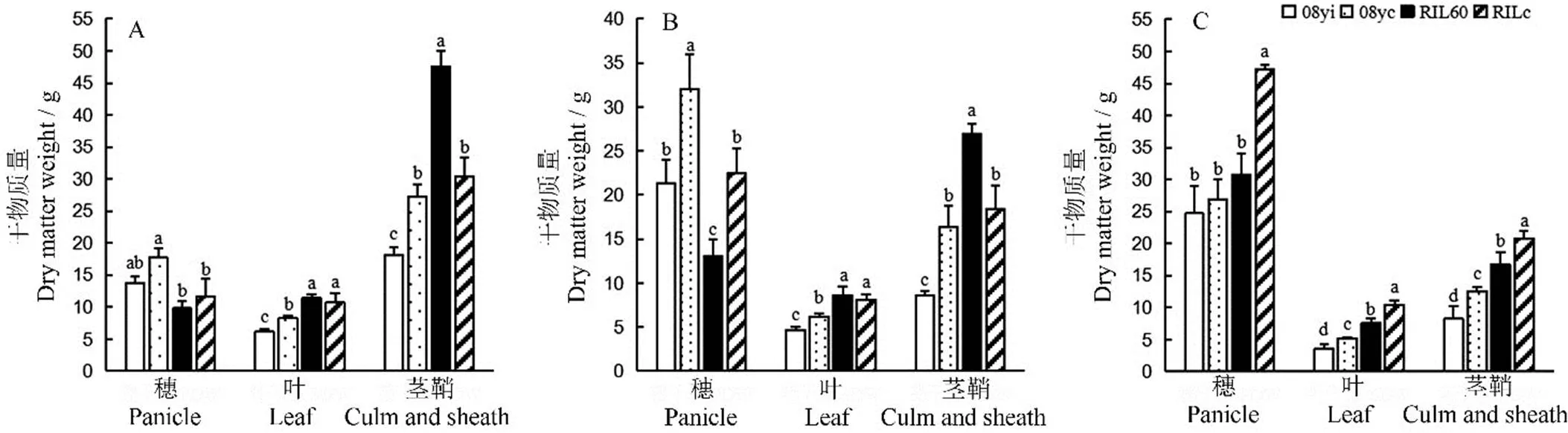
A?乳熟期;B?蠟熟期;C?完熟期。均值±標準差,不同小寫字母表示不同種質差異顯著(P<0.05, n=4)(F檢驗)。
Fig. 3. Comparison of dry matter content of different germplasms at different developmental stages.
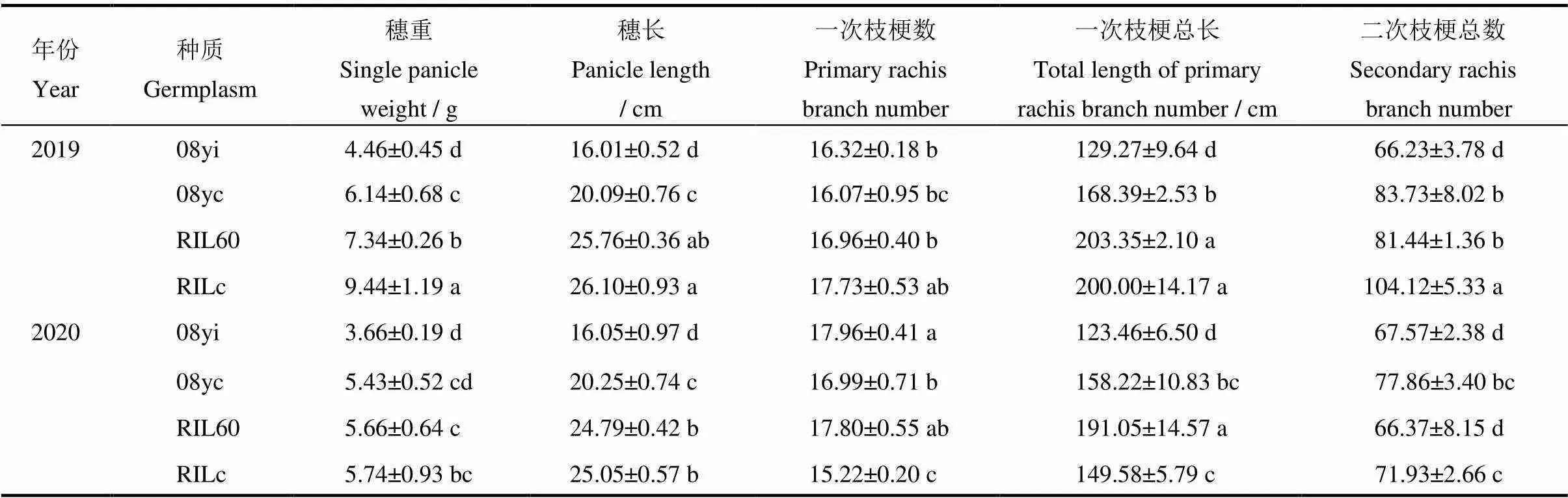
表3 不同種質的穗部性狀比較
均值±標準差。同一列中不同小寫字母表示不同種質以及同一種質不同年份間差異顯著(<0.05,=8)(-檢驗)。
Mean±SD.Within a column, different lowercase letters mean significantdifference at<0.05by-test(=8).
對2019年4份種質的穗部性狀進行差異顯著性分析(-檢驗)及多重比較(表3),結果表明新種質08yi的一次枝梗數與08yc無顯著性差異,平均每穗達到16個,但其單穗重較08yc降低了27.36%,穗長降低了20.31%,一次枝梗總長減少了23.23%,二次枝梗總數減少了20.90%。2020年由于氣候異常影響,兩種質的部分穗特征值與2019年比差異較大(表3),08yi穗重降低了17.94%,一次枝梗數增加了10.05%;08yc穗重降低了11.56%,一次枝梗總長減少了6.04%,二次枝梗數減少了7.01%。2019年,RIL60的穗重比RILc降低了22.25%,穗長減少了20.74%,一次枝梗數減少了14.49%,二次枝梗總數減少了21.78%。2020年各指標與2019年比,RIL60的穗重降低了22.89%,二次枝梗數減少了18.50%;RILc穗重降低了39.19%,穗長減少了4.23%,一次枝梗數減少了14.16%,一次枝梗總長減少了25.21%,二次枝梗總數減少了30.92%。
說明兩份新種質雖然在穗部性狀上的表現不如常規種質,但2020年灌漿期間的連續降雨嚴重影響光合作用,在如此逆境中RIL60的穗特征值降幅較小,說明其抗逆性可能較強。
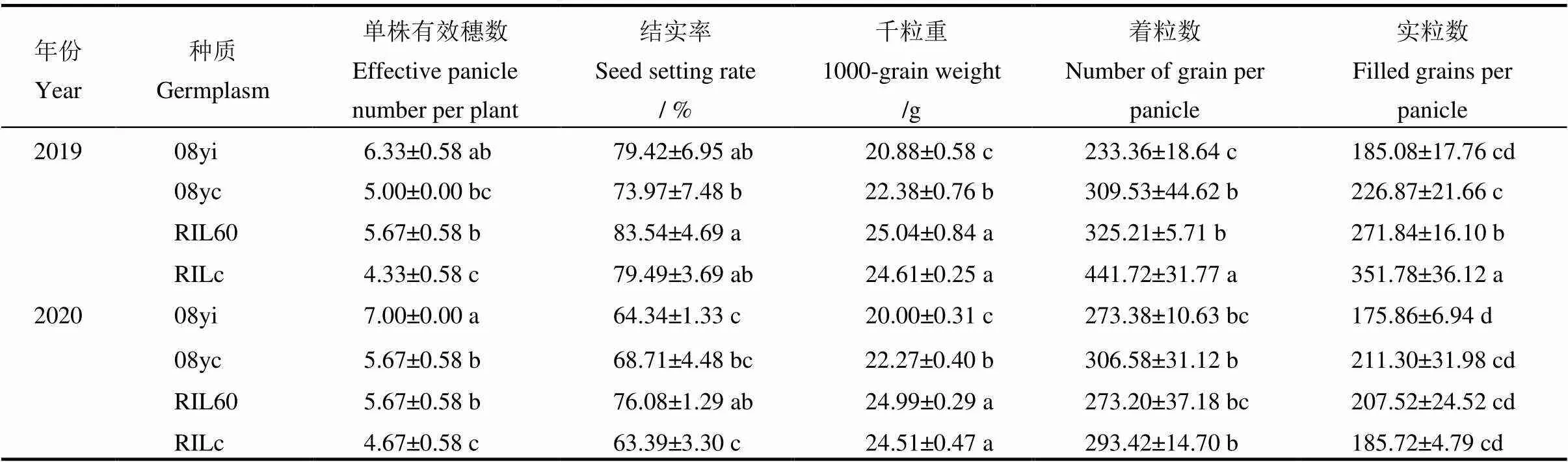
表4 不同種質單株產量性狀比較
均值±標準差。同一列中不同小寫字母表示不同種質以及同一種質不同年份間差異顯著(<0.05,=8)(-檢驗)。
Mean±SD. Different lowercase letters in the same column are significantly different at<0.05 among different germplasm and between different years of the same germplasm by-test(=8).
2.4.2 產量及相關農藝性狀
對兩年的農藝性狀進行差異顯著性分析(-檢驗)和多重比較,結果顯示兩新種質的單株有效穗數均顯著多于對應的常規種質(<0.05)(表4)。2019年,新株型08yi的結實率比08yc提高了7.37%,但千粒重減少了6.70%,其每穗著粒數和實粒數比08yc均減少,但是僅每穗著粒數差異達到顯著水平(<0.05)。兩株型間的單株產量和著粒密度并沒有顯著差異(圖4-A、B),但新株型08yi的株高比08yc矮11.39 cm(<0.05)(圖4-C)。2020年,08yi的每穗著粒數比2019年增加了17.15%,著粒密度增加了16.56%。2020年,兩新種質的單株產量、結實率、每穗實粒數和株高等性狀比2019年均減少,但僅株高差異達到顯著水平(<0.05)。
2019年,新株型RIL60的千粒重和結實率等單株產量性狀比RILc均增加,但差異不顯著。而RIL60的每穗著粒數和實粒數則顯著低于RILc(表4)。由圖4-A可知,兩株型間的單株產量并無顯著性差異,而RIL60的株高比RILc增加了4.26 cm(圖4-C),但其著粒密度卻降低了25.95%(圖4-B)。2020年的各性狀與2019年相比,兩種質在單株產量、結實率、每穗著粒數、每穗實粒數、著粒密度和株高等性狀上均顯著下降,但RILc比RIL60下降幅度大。說明在逆境條件下新種質RIL60比RILc有更好的適應能力。
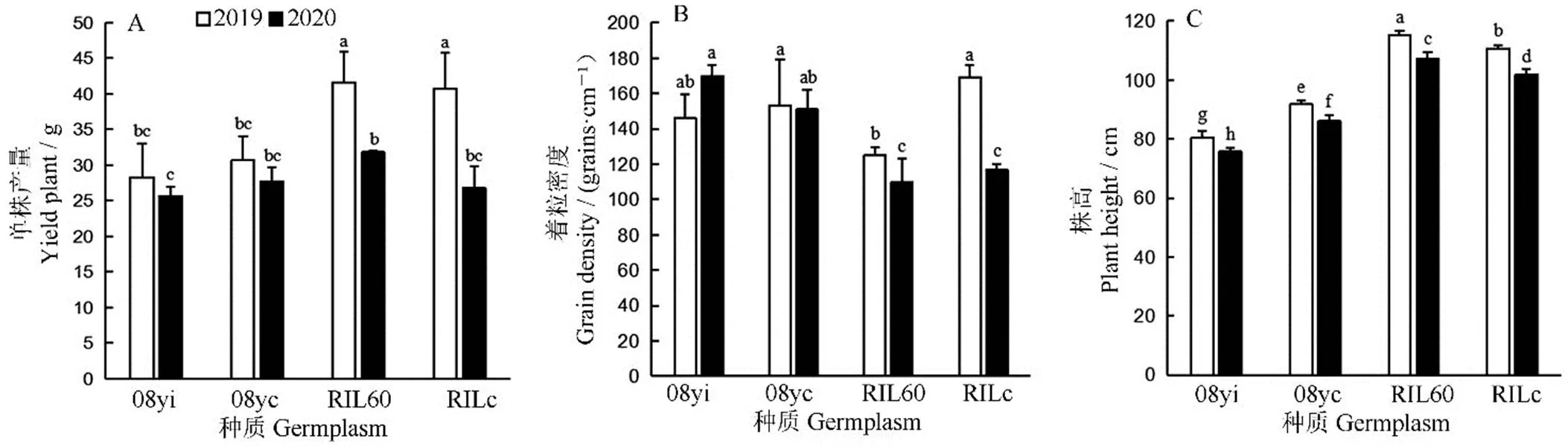
均值±標準差。不同小寫字母表示不同種質間以及同一種質不同年份間差異顯著(P<0.05, n=8, F-檢驗)。
Fig. 4. Comparison of yield per plant and its components for different germplasms.
2.5 新種質遺傳差異
2.5.1 理想株型基因測序
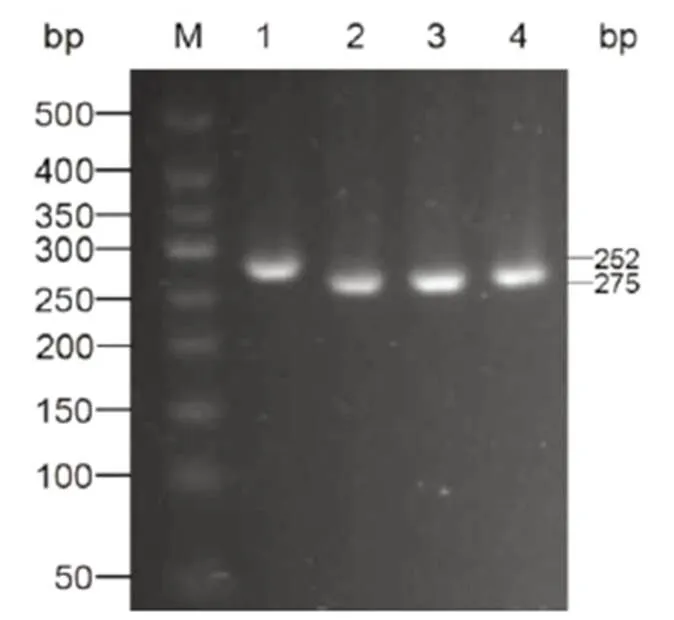
根據測序結果可知(圖5),與少蘗粳的相比,兩份新種質在基因的第874位點上并未發生單堿基突變(C→A),說明新種質株型差異并非基因在該位點突變所致。
2.5.2 稻米糯性變異
利用分子標記,鑒定兩份新種質與相應常規種質的糯性基因型,結果表明新種質RIL60發生糯性變異,新種質08yi未發生糯性變異(圖6)。
3 討論
水稻葉片形態結構是衡量株型是否理想的重要指標,它與光能利用率密切相關,發掘功能葉性狀理想的新種質對理想株型育種有重要意義。葉片卷曲度、披垂度、葉片角以及葉長和葉寬是株型育種的主要研究方面,其中葉角能夠決定葉片的空間伸展姿態,對水稻群體的受光面積有重要影響[21];葉片長寬決定水稻葉面積,是衡量光能利用的重要指標[16]。就葉長而言,倒2葉>倒3葉>劍葉,倒2葉和倒3葉都比劍葉長近40%時能達到高產[22]。本研究中,兩份新種質的功能葉長度比例更接近該理想狀態。高產水稻功能葉具有狹長、直立、窄厚等特點,葉片直立,葉角小是已有研究表明的理想葉型特點,直立葉能夠兩面受光,又能減少葉片相互遮蔽,使得植株整體受光面積最大化,有效提升群體光合效率[23]。葉傾角小不但能夠增大植株下部葉片的受光面積,提高整體光能利用率,并且不利于害蟲的著落,從而有效防治病蟲害[24]。本研究中發現的兩份新種質功能葉的葉角總體上小于常規種質,且更符合理想株型上部三葉長勢特點:劍葉葉傾角最小、倒2、倒3葉的葉傾角依次增大使植株呈塔形,能獲得最大程度的陽光照射,提高光能利用率[25]。新種質株型優勢為通過合理密植提高單位面積產量提供了資源選擇。

圖5 四份種質IPA1 基因測序結果
Fig. 5. Sequencing ofgene of the four germplasms.
1, RIL60; 2, RILc; 3, 08yi; 4, 08yc.
Fig. 6. Result of identification ongene for the four germplasms.
葉片形態、葉面積和葉綠素含量是影響植物光合作用的主要因素,而干物質積累又是光合作用的最終形式,與產量密切相關[26]。本研究中雖然新種質的葉形態更理想,但其葉面積顯著減少,RIL60的葉綠素含量顯著降低,導致新種質在不同生育時期部分器官的干物質量比常規種質少。水稻抽穗至成熟期的干物質積累量對產量影響最大,干物質積累與產量成正相關[27];在此階段,新種質RIL60與RILc相比,出現葉面積降低和葉綠素減少的現象,但在完熟期二者的單株產量并無顯著性差異,這表明理想株型種質有更強的光合能力和干物質積累能力。另外,劉巧泉等[15]研究表明,當品種的直鏈淀粉含量降低時,其粒重會下降,糯性位點對稻米粒重具有負效應。因此,這也是RIL60與RILc相比沒有產量優勢的原因之一。但RIL60的理想株型結構為糯稻通過合理密植提高光合效率和干物質積累量進而提高單產指明了選育方向。
水稻穗是理想株型育種和高產育種的重要考查性狀,包括有效穗數、穗形、穗長、穗重、千粒重和每穗粒數等多個因子,其中有效穗、千粒重和實粒數對單株產量貢獻最大[28]。單株有效穗與分蘗能力直接相關,單株有效穗數多,無效分蘗少,能最大限度地提高單株產量,但其同時又制約著每穗粒數的增加,單株有效穗與每穗著粒數呈負相關[29]。本研究中兩份新種質的單株有效穗數均比常規種質增加1~2穗,但其每穗著粒數和每穗實粒數均下降,導致其單株產量與常規種質比并無顯著差異。穗型改良過程中,穗形、穗長、穗重是主要改良因子,與之相應的“直立穗型”、“大穗型”和“重穗型”等高產水稻的成功培育[30],使穗部性狀的變異為水稻超高產育種提供了豐富的種質資源。本研究中發現的兩份新種質在穗長、穗重、一次枝梗、二次枝梗等性狀上均比常規種質有較大的變異幅度,有利于進一步解析其分子機理,為構建高產的優化穗型模式打好基礎。
利用分子標記輔助育種,挖掘控制株型和產量的基因,進行更加高效和精準的水稻分子設計育種,是實現株型改良、構建理想株型的重要手段。目前,很多關于控制水稻株型和產量的基因已被克隆,包括調控水稻分蘗的基因[31];控制水稻匍匐生長變成直立生長,改良株型的關鍵基因[32];控制粒長和粒重的主效QTL[33];調控水稻籽粒和葉傾角的基因[34];調控穗型和粒數的關鍵基因[35];調控分蘗和產量性狀的關鍵QTL[11]。這些基因的發現和相關功能的發掘為構建理想株型提供了分子育種基礎。尤其是理想株型基因的一個點突變后干擾miR156的識別,使得無效分蘗減少、千粒重和著粒數增加,同時莖稈變粗,抗倒伏能力增強,產量增加了10%以上,使理想株型育種實現了里程碑式的突破[12-13]。本研究在通過基因測序后發現,新種質在該位點并沒有發生點突變,其株型變異是由其他基因變異或者在該基因其他位點上發生突變所致。這兩份新株型種質將是株型分子調控研究的重要材料。
4 結論
新種質08yi和RIL60在莖、葉和穗等性狀上均比對應的常規種質發生了明確的有利變異。在農藝性狀方面,新株型種質的功能葉葉角顯著減小,葉長比例更合理,株型更緊湊,它們與少蘗粳在株型結構上具有相似性,但遺傳分析顯示兩新種質并未突變成少蘗粳。兩新種質的每穗有效穗數均增加了1~2穗,且在逆境下穗部性狀表現較好,產量降幅較小,抗逆能力增強。但在穗部性狀上也有不利變異,其單穗重、穗長、枝梗性狀、每穗著粒數和每穗實粒數均下降。此外,RIL60發生了糯性變異,但與常規株型非糯種質比,其單株產量并未下降。兩份新株型種質可作為優異的株型材料用于理想株型育種和株型分子調控研究。
[1] 吳比, 胡偉, 邢永忠. 中國水稻遺傳育種歷程與展望[J]. 遺傳, 2018, 40(10): 841-857.
Wu B, Hu W, Xing Y Z. The history and prospect of rice genetic breeding in China[J].(), 2018, 40(10): 841-857. (in Chinese with English abstract)
[2] 胡娟, 林晗, 徐娜, 焦然, 戴志俊, 魯草林, 饒玉春, 王躍星. 水稻葉傾角分子機制及育種應用的研究進展[J]. 中國水稻科學, 2019, 33(5): 391-400.
Hu J, Lin H, Xu N, Jiao R, Dai Z J, Lu C L, Rao Y C, Wang Y X. Advances in molecular mechanisms of rice leaf inclination and its application in breeding[J]., 2019, 33(5): 391-400. (in Chinese with English abstract)
[3] 李紅, 何煒, 連玲, 魏毅東, 蔡秋華, 王穎姮, 謝華安, 張建福. 水稻株型的研究進展[J]. 福建稻麥科技, 2020, 38(4): 61-66.
Li H, He W, Lian L, Wei Y D, Cai Q H, Wang Y H, Xie H A, Zhang J F. Research advances on plant type of rice[J]., 2020, 38(4): 61-66. (in Chinese with English abstract)
[4] Guo W, Chen L, Herrera-Estrella L, Cao D, Tran L S P. Altering plant architecture to improve performance and resistance[J]., 2020, 25(11): 1154-1170.
[5] Donald C M. The biological yield and harvest index of cereals as agronomic and plant breeding criteria[J]., 1976, 28(4): 361-405.
[6] Khush G S. Prospects and approaches to increasing the genetic yield potential of rice[M]//Evenson R E, Herdt R N, Hossain M. Rice Research in Asia: Progress and Priorities. CAB International and IRRI, 1996: 59-71.
[7] 楊守仁, 張龍步, 陳溫福, 徐正進, 王進民. 水稻超高產育種的理論和方法[J]. 中國水稻科學, 1996, 10(2): 115-120.
Yang S R, Zhang L B, Cheng W F, Xun Z J, Wang J M. Theories and methods of rice breeding for maximum yield[J]., 1996, 10(2): 115-120. (in Chinese with English abstract)
[8] 馬夢影, 鞏文靚, 康雪蒙, 段海燕. 水稻理想株型改良的研究進展[J]. 中國農學通報, 2020, 36(29): 1-6.
Ma M J, Gong W J, Kang X M, Duan H Y. The improvement of ideal plant type of rice: A review[J]., 2020, 36(29): 1-6. (in Chinese with English abstract)
[9] Chen W F, Xu Z J, Zhang W Z, Zhang L B, Yang S R. Creation of new plant type and breeding rice for super high yield[J]., 2001, 27(5): 665-672.
[10] Zhao S, Xiang J, Xue H. Studies on the rice, an IAA-amido synthetase, reveal the effects of auxin in leaf inclination control[J]., 2013, 6(1): 174-187.
[11] Jiao Y Q, Wang Y H, Xue D W, Wang J, Yan M, Liu G, Dong G, Zeng D, Lu Z, Zhu X. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice[J]., 2010, 42(6): 541-544.
[12] Miura K, Ikeda M, Matsubara A, Song X J, Ito M, Asano K, Matsuoka M, Kitano H, Ashikari M.promotes panicle branching and higher grain productivity in rice[J]., 2010, 42(6): 545-549.
[13] Wang J, Zhou L, Hui S, Mawsheng C, Hong Y, Hong Y, Min H, Yin J, Zhu X, Li Y. A single transcription factor promotes both yield and immunity in rice[J]., 2018, 361(6406): 1026-1028.
[14] Jeng T L, Wang C S, Tseng T H, Wu M T, Sung J M. Nucleotide polymorphisms in thegene of NaN3-induced waxy rice mutants[J]., 2009, 49(1): 112-116.
[15] 劉巧泉, 王興穩, 陳秀花, 王宗陽. 轉反義基因糯稻的顯性遺傳及對稻米粒重的效應分析. 中國農業科學, 2002, 35(2): 117-122.
Liu Q Q, Wang X Y, Chen X H, Wang Z Y. Effect of dominant waxy character on kernel weight of transgenic rice with antisensegene., 2002, 35(2): 117-122. (in Chinese with English abstract)
[16] 丁維龍, 謝濤, 徐利鋒, 張義凱. 基于虛擬模型的水稻冠層葉面積計算方法[J]. 農業工程學報, 2017, 33(2): 192-198.
Ding W L, Xie T, Xu L F, Zhang Y K. Calculation method of rice canopy leaf area based on virtual model[J]., 2017, 33(2): 192-198. (in Chinese with English abstract)
[17] 周振翔, 李志康, 陳穎, 王志琴, 楊建昌, 顧駿飛. 葉綠素含量降低對水稻葉片光抑制與光合電子傳遞的影響[J]. 中國農業科學, 2016, 49(19): 3709-3720.
Zhou Z X, Li Z K, Chen Y, Wang Z Q, Yang J C, Gu J F. Effects of reduced chlorophyll content on photo- inhibition and photosynthetic electron transport in rice leaves[J]., 2016, 49(19): 3709-3720. (in Chinese with English abstract)
[18] 陳家潤. 水稻品種試驗田間記載及室內考種項目與標準[J]. 湖北農業科學, 1957(2): 146-148.
Chen J R. Field records and laboratory test items and standards of rice variety test[J]., 1957(2): 146-148. (in Chinese with English abstract)
[19] Wendland J, Lengeler K B, Kothe E. An instant preparation method for nucleic acids of filamentous fungi[J]., 1996, 43(1): 54-55.
[20] 田志喜, 嚴長杰, 錢前, 嚴松, 謝會蘭, 王芳, 徐潔芬, 劉貴富, 王永紅, 劉巧泉, 湯述翥, 李家洋, 顧銘洪. 水稻淀粉合成相關基因分子標記的建立[J]. 科學通報, 2010, 55(26): 2591-2601.
Tian Z X, Yan C J, Qian Q, Yan S, Xie H L, Wang F, Xu J F, Liu G F, Wang Y H, Liu Q Q, Tang S Z, Li J Y, Gu M H. Establishment of molecular markers of rice starch synthesis related genes[J]., 2010, 55(26): 2591-2601. (in Chinese with English abstract)
[21] 徐靜, 王莉, 錢前, 張光恒. 水稻葉片形態建成分子調控機制研究進展[J]. 作物學報, 2013, 39(5): 767-774.
Xu J, Wang L, Qian Q, Zhang G H. Research advance in molecule regulation mechanism of leaf morphogenesis in rice (L.)[J]., 2013, 39(5): 767-774. (in Chinese with English abstract)
[22] 楊從黨, 袁平榮, 周能, 朱德峰, 楊愛兵, 鄭學玉, 黃慶宇, 應繼鋒. 葉型特性與產量構成因素的相關分析[J]. 中國水稻科學, 2001, 15(1): 71-73.
Yang C D, Yuan P R, Zhou N, Zhu D F, Yang A B, Zheng X Y, Huang Q Y, Ying J F. Analysis on relationship between characters of leaf type and yield components[J]., 2001, 15(1): 71-73. (in Chinese with English abstract)
[23] 賀勇, 孫煥良, 孟桂元. 水稻葉片形態研究進展[J]. 作物研究, 2008(S1): 378-380.
He Y, Sun H L, Meng G Y. Advances in leaf morphology of rice[J]., 2008(S1): 378-380. (in Chinese with English abstract)
[24] 羅贛豐, 馬雪梅, 程建峰. 水稻劍葉角度與氮營養效率的關系[J]. 中國農學通報, 2014, 30(18): 29-34.
Luo G F, Ma X M, Cheng J F. The relationships between flag leaf angles of various rice germplasms and their nitrogen nutrition efficiencies[J]., 2014, 30(18): 29-34. (in Chinese with English abstract)
[25] Easterling W, Apps M. Assessing the consequences of climate change for food and forest resources: A view from the IPCC[J]., 2005, 70: 165-189.
[26] 孔飛揚, 江立庚, 文娟, 郝向陽. 直播水稻產量、產量構成因子和干物質積累的變化特點及其相互關系[J]. 華中農業大學學報, 2018, 37(5): 11-17.
Kong F Y, Jiang L G, Wen J, Hao X Y. Changes and relationships of yield, yield components and dry matter accumulation of direct-seeded rice[J]., 2018, 37(5): 11-17. (in Chinese with English abstract)
[27] Peng S B, Li Y, Cui K H, Yu T T, Wang T, Xu L, Zhan X W, Huang J L, Nie L X. Yield performance of direct-seed, double-season rice using varieties with short growth durations in central China[J]., 2018, 227: 49-55.
[28] 武玲, 向珣朝, 楊博文, 許亮, 顏李梅. 水稻秈爪交重組自交系穗部性狀變異分析[J]. 中國農業大學學報, 2017, 22(10): 26-35.
Wu L, Xiang X C, Yang B W, Xu L, Yan L M. Analysis on the panicle traits variation of recombinant inbred line derived from the hybridization ofand[J]., 2017, 22(10): 26-35. (in Chinese with English abstract)
[29] 陳小榮, 陳志彬, 賀浩華, 朱昌蘭, 彭小松, 賀曉鵬, 傅軍如, 歐陽林娟. 水稻單株有效穗數主基因+多基因混合遺傳分析[J]. 生物數學學報, 2011, 26(3): 555-562.
Chen X R, Chen Z B, He H H, Zhu C L, Peng X S, He X P, Fu J R, OuYang L J. Genetic analysis of panicles per plant in rice by the major genes plus polygenes mixed inheritance model[J]., 2011, 26(3): 555-562. (in Chinese with English abstract)
[30] 劉堅, 陶紅劍, 施思, 葉衛軍, 錢前, 郭龍彪. 水稻穗型的遺傳和育種改良[J]. 中國水稻科學, 2012, 26(2): 227-234.
Liu J, Tao H J, Shi S, Ye W J, Qian Q, Gu L B. Genetics and breeding improvement for panicle type in rice[J]., 2012, 26(2): 227-234. (in Chinese with English abstract)
[31] Li X, Qian Q, Fu Z, Wang Y, Xiong G, Zeng D, Wang X, Liu X, Sheng T, Hiroshi F. Control of tillering in rice[J]., 2003, 422: 618-621.
[32] Jin J, Huang W, Gao J P, Yang J, Lin H X. Genetic control of rice plant architecture under domestication[J]., 2008, 40: 1365-1369.
[33] Fan C, Xing Y, Mao H, Lu T, Zhang Q., a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein[J]., 2006, 112: 1164-1171.
[34] Feng Z, Wu C, Wang C, Roh J, Zhang L, Chen J, Zhang S, Zhang H, Yang C, Hu J, You X, Liu X, Yang X, Guo X, Zhang X, Wu F, Terzaghi W, Kim S K, Jiang L, Wan J.controls grain size and leaf angle by modulating brassinosteroid homeostasis in rice[J]., 2016, 67(14): 4241-4253.
[35] Huang X, Qian Q, Liu Z, Sun H, He S, Luo D, Xia G, Chu C, Li J, Fu X. Natural variation at thelocus enhances grain yield in rice[J]., 2009, 41: 494-497.
Analyses on Agronomic Traits and Genetic Characteristics of Two New Plant-architecture Lines in Rice
LIANG Cheng, XIANG Xunchao*, ZHANG Ouling, YOU Hui, XU Liang, CHEN Yongjun
(Laboratory of Plant Molecular Genetics and Breeding, Southwest University of Science and Technology, Mianyang 621010, China;*Corresponding author, E-mail: xxc@swust.edu.cn)
【】Ideal plant-architecture (IPA) can increase photosynthetic efficiency, economic yield and resistance to disease of rice. The objective is to develop excellent germplasms with IPA for high-yield and resistance breeding by exploringvariation in traits and genetic characteristics of two rice germplasm with new plant-architectures. 【】The two germplasms ‘08yi’ and ‘RIL60’ with new plant-architecturefrom two recombinant inbred lines (RIL) by natural recombination, and their corresponding conventional germplasm ‘08yc’ and ‘RILc’ from the same RIL were used asmaterials. The constitute factors of plant architecture, yield traits in different developmental periods, as well as sequencing ofgene and waxy identification in four germplasm were comparatively analyzed. 【】The functional leaf traits, dry matter accumulation at different developmental stages as well as panicle and yield traits of the two new lines were significantly different from those of conventional germplasms. Compared to the corresponding conventional germplasm, ‘08yi’ and ‘RIL60’ were featured with favorable variations including reduced angles of functional leaf, proper leaf length,compact plant architecture, an increase of 1-2 panicle in the effective panicles per plantand increased seed setting rateby 7.37% and 5.09%, respectively. In addition, the chlorophyll contents of the new germplasm ‘08yi’ significantly increased, and the stem of ‘RIL60’ was thicker than that of ‘RILc’. The panicle traits of the two new germplasm under stress condition were better because they had smaller reduction of economic yield and stronger stress resistance compared to their corresponding conventional germplasm. However, their unfavorable variations include decreased panicle weight, shorter panicle length, decreasednumber of grains per panicle, seeds, branch length and branch number. The results ofgene sequencing showed that there were no mutations at 874 site of(C→A) in the two new germplasm compared with the Shaoniejing, and the variation of plant architecture was attributed to other factors. The result of waxy identification showed that ‘RIL60’ had waxy variation. 【】Thanks for preferred variation in agronomic traits, the two new lines can be used as excellent germplasms for IPA breeding and research. As a glutinous rice variety with IPA and economic yield equal to non-glutinous rice, RIL60 has enormous potential for extension.
rice (L.); ideal plant architecture; agronomic trait; genetic characteristics;gene
10.16819/j.1001-7216.2022.210208
2021-02-06;
2021-06-03。
四川省科技廳重點研發計劃(重大科技專項)(2019YFN0019);西南科技大學研究生創新基金資助項目(20ycx0038)。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
音樂探索(2022年2期)2022-05-30 21:01:37
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
