抗真菌劑三峽肽素抑制指狀青霉的轉錄組學研究
2022-03-17 08:58:56權雯婧薛艷紅劉士平
食品工業科技 2022年6期
關鍵詞:差異
權雯婧,劉 超,李 奧,薛艷紅,劉士平
(三峽大學生物與制藥學院,中國輕工業功能酵母重點實驗室,湖北宜昌 443002)
柑橘是我國南方地區農村經濟的支柱產業之一[1],每年都有大量的柑橘因指狀青霉(P.digitatum)等采后致腐真菌的影響而發生腐敗和品質劣變[2],造成巨大經濟損失[3]。化學殺菌劑百可得、咪鮮胺等雖對柑橘的防腐有一定的效果[4],但處理后的果品品質發生劣變,糖份降低,而且對人體有毒副作用,殘留時間長且對環境有污染,許多國家已禁止化學殺菌劑在果蔬采后處理中使用[5-6]。因此,開發出一種綠色健康的保鮮劑顯得極為緊迫[7-8]。
近年來抗真菌肽(Antifungal Peptide,AFP)因其抑菌能力強、結構多樣、無毒副作用受到了人們的廣泛關注,如表面活性素、芬薺素、伊枯草菌素、桿菌霉素和羊毛硫肽等[9],在醫藥、食品、農業和飼料添加劑等領域中發揮著越來越大的作用[10]。這類物質因其作用的靶點多與傳統的抗生素不一樣,因此微生物不易產生抗藥性,如羊毛硫肽通過在質膜上形成膜孔而達到殺菌的效果[11]。抗真菌肽的作用機制比較復雜,其可能的作用靶點為:a.影響細胞壁上的幾丁質或者聚糖代謝,阻礙細胞壁的功能[12];b.作用于細胞膜,使細胞膜通透性增加,內容物流出[13];c.作用于細胞內的細胞器或物質(如線粒體、蛋白質、核酸等),使細胞生長和繁殖受到抑制,從而達到抑菌的目的[14-15]。
在前期研究中,本實驗室從三峽河岸地區分布的草酸青霉(Penicillium oxalicum)中分離到一種新型的線性五肽,命名為三峽肽素(Sanxiapeptin),研究發現它在離體條件下或柑橘體內均對柑橘的主要采后致腐菌指狀青霉具有強烈的抑制作用[16],而且其抑菌能力遠優于目前市場上廣泛應用的化學殺菌劑百可得[17]。為了明確三峽肽素的抑菌機理,本研究通過形態學觀察、保護劑的抑菌效果以及轉錄組測序的方法,初步確定了三峽肽素可能的抑菌靶點。本研究不僅有助于豐富抗真菌肽的作用機理,還將有助于新型綠色健康的柑橘保鮮劑的研發,對促進我國農業經濟的發展具有重要意義。
1 材料與方法
1.1 材料與儀器
新鮮土豆 市售;葡萄糖 科密歐化學試劑有限公司;瓊脂、麥角固醇、山梨醇 金燕海洋生物股份有限公司;色譜級乙腈 美國天地有限公司;真菌RNA提取試劑盒 美國Omega Bio-Tek公司;百可得上海日曹達貿易有限公司;反轉錄試劑Goldenstar?RT6 cDNA Synthesis Kit Ver.2、T3 Super PCR Mix北京擎科新業生物技術有限公司。
BS-224s電子天平 德國賽多利斯有限責任公司;SW-CJ-2FD型雙人實驗操作臺 蘇州凈化設備廠;XPX-9052 MBE數顯培養箱 上海博訊實業有限公司醫療設備廠;ZQZY-85BN全自動控溫搖床上海知楚儀器有限公司;Master-s15純水儀 上海和泰儀器有限公司;MyCyclerTM Thermal PCR儀 美國BIO-RAD公司;GDS-8000凝膠成像系統 美國UVP公司;Allegra 64R臺式高速冷凍離心機 美國BECKMAN公司;DYCP-31DN水平電泳儀 北京市六一儀器廠;YM30F不銹鋼智能型立式電熱蒸汽消毒器 上海三申醫療器械有限公司;光學顯微鏡下觀察 奧林巴斯株式會社;S3000N掃描電子顯微鏡、H-7500透射電子顯微鏡 Hitachi日本。
1.2 菌株及培養條件
本實驗所使用的菌株為指狀青霉P.digitatum(CICC 41633),本實驗室自行分離鑒定[18-19]。菌株保藏于4 ℃的PDA固體培養基斜面中,培養基配方:土豆 200 g,葡萄糖20 g,瓊脂15~20 g,水 1000 mL,pH自然,121 ℃滅菌20 min。菌株經活化后,采用不加瓊脂的PDA液體培養基進行培養,培養溫度為28 ℃,搖床轉速控制在120 r/min。
1.3 三峽肽素對菌株形態的影響
將指狀青霉在上述條件下培養3 d后,分別添加0~50 μg/mL的三峽肽素和正對照百可得,繼續培養12 h[20]。在光學顯微鏡觀察時,將培養物在5000 r/min離心5 min后,收集菌絲制備玻片進行觀察[21]。掃描電子顯微鏡和透射電子顯微鏡的觀察委托百奧生物科技有限公司(湖北,武漢)按照文獻所示方法完成[22],每次試驗重復3次。
1.4 麥角固醇和山梨醇的補償分析
為了初步判斷三峽肽素抑制指狀青霉的可能作用靶點,本研究分別采用絲狀真菌細胞膜的保護劑麥角固醇和細胞壁的保護劑山梨醇進行補償實驗。將0、31.25、62.5、125、250、500、1000 μg/mL 的麥角固醇或山梨醇添加在PDA固體培養基中,在培養皿中央放置含有1 mg/mL的三峽肽素或百可得的牛津杯[23-24],在28 ℃的培養箱中培養3 d后測定抑菌圈直徑,通過抑菌圈大小的變化判斷可能的補償效應。每次試驗設置4個重復。
1.5 轉錄組學分析
將指狀青霉在1.2所述培養條件下培養3 d后,分別添加15 μg/mL的正對照百可得(PD-2)和三峽肽素(PD-3),繼續培養6 h,同時設置相同時長生長的指狀青霉作為正常組(PD-1)。將培養物在5000 r/min離心5 min后,收集菌絲提取RNA后,送至華大基因進行轉錄組測序。將獲得的轉錄組數據使用華大基因自主研發的過濾軟件SOAPnuke(v1.4.0)進行統計,使用 Trimmomatic進行過濾,得到clean reads,并將過濾后的數據中大于20個堿基的序列所占的比例數進行統計;使用Bowtie2(v2.2.5)軟件將clean reads進行比對,并將能相互比對上的凈數據比例進行統計。根據比對結果,利用RSEM(RNA-Seq by Expectation Maximization,v1.2.8)軟件進行定量分析,并以FPKM(Fragments Per Kilobase Per Million reads)計算基因的表達水平,差異表達基因默認定義為 FDR(False Discovery Rate)≤0.001且倍數差異在1倍以上的基因。將篩選到的差異表達基因,采用 GO(Gene Ontology)和 KEGG(Kyoto Encyclopedia of Genes and Genomes)數據庫對結果進行注釋分類,同時使用R軟件中的phyper函數進行富集分析,同時將檢測到的基因進行在NR(Nucleotide Sequence Database)和 NT(Non-Redundant Protein Sequence Database)數據庫中進行注釋和歸類,判斷基因可能的功能[25]。
1.6 RT-PCR驗證
使用Oligo7軟件設計引物,由生工生物工程(上海)股份有限公司合成,使用反轉錄試劑盒Goldenstar?RT6 cDNA Synthesis Kit Ver.2將所提RNA反轉錄成cDNA,測定并調整cDNA濃度之后,采用半定量RT-PCR結合電泳條帶的亮度進行驗證[26],所用的引物序列見表1。PCR反應體系為模板DNA:2 μL,T3 Super PCR Mix:20 μL,上游和下游引物各 1 μL,總體系 50 μL。PCR 程序 95 ℃ 3 min,然后是 95 ℃變性20 s,依據各引物的不同將退火溫度設置為Tm+5 ℃,復性 20 s,72 ℃ 延伸 20 s,經過 30~35 循環后再在72 ℃延伸5 min。將PCR產物采用1.2%瓊脂糖凝膠進行電泳檢測。
2 結果與分析
2.1 三峽肽素對指狀青霉菌絲的形態影響
在培養條件下,三峽肽素可以顯著抑制指狀青霉的生長(圖1),而且在同等濃度下其抑制能力優于市場上廣泛使用的農藥百可得(雙胍三辛烷苯基磺酸鹽,一種以細胞膜為靶點的殺真菌劑)[27-28]。為了進一步了解三峽肽素對指狀青霉菌絲形態的影響,本實驗采用百可得為陽性對照,設置了0~50 μg/mL的濃度梯度,光學顯微鏡觀察顯示,三峽肽素和百可得在濃度超過20 μg/mL時均會導致指狀青霉的菌絲發生消融,在15 μg/mL時,不同組別的菌絲的形態變化是最為明顯的,因此選用此濃度處理后的菌絲進行掃描電子顯微鏡和透射電子顯微鏡的觀察。觀察結果顯示:三峽肽素和百可得兩者都使指狀青霉的菌絲明顯變粗,但經三峽肽素處理后,菌絲體明顯形成結節扭曲狀(圖2A~圖2C)。
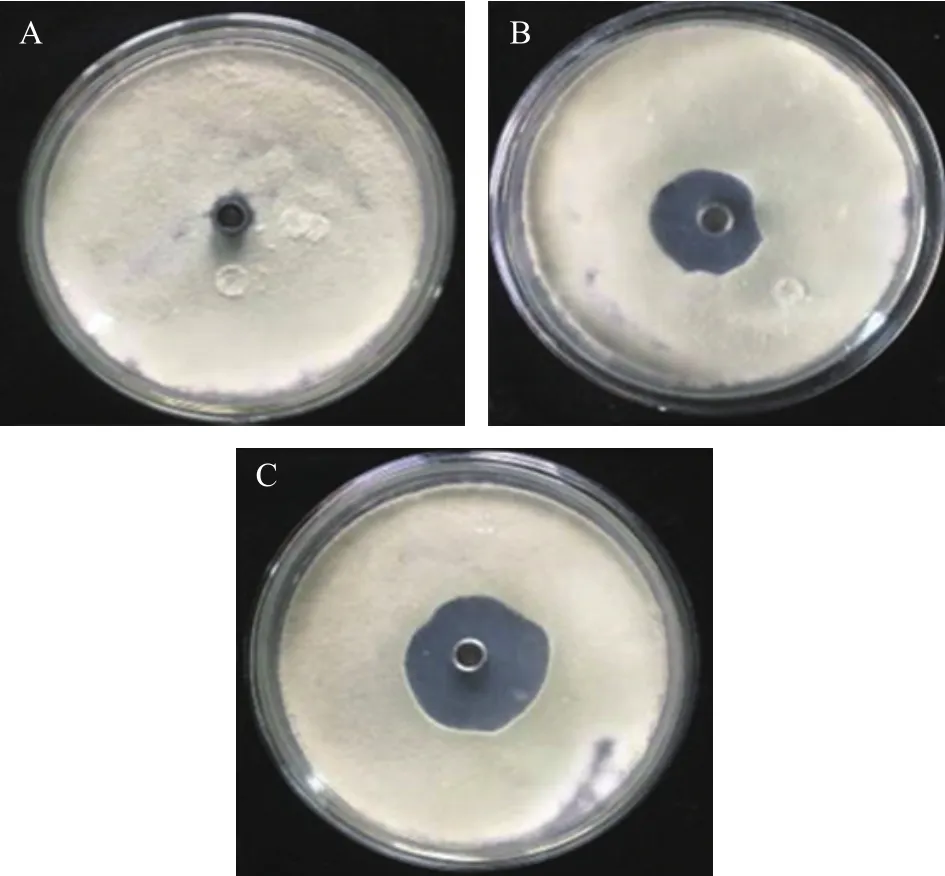
圖1 三峽肽素對指狀青霉的拮抗作用Fig.1 Antagonistic effect of Sanxiapeptin on P.digitatum
為了進一步了解指狀青霉的顯微形態變化,在相同濃度處理后分別進行了掃描電鏡和透射電鏡觀察。掃描電鏡結果發現,在添加百可得后,菌絲體的扭曲程度和正常生長的指狀青霉沒有明顯形態差異,而三峽肽素可以明顯導致菌絲體發生結節和扭曲(圖2D~圖2F)。透射電鏡觀察結果顯示,經三峽肽素處理后的指狀青霉細胞內部出現了空腔,細胞邊緣發生破裂、不完整,而作用于細胞膜的物質百可得處理后細胞邊緣仍然較為完整,細胞內部也沒有出現空腔(圖2G~圖2L)。上述結果表明,三峽肽素與百可得的作用機理可能不一樣,可能通過破壞細胞壁,造成滲透壓紊亂,使得胞內物質外流,從而導致胞內空腔的形成。
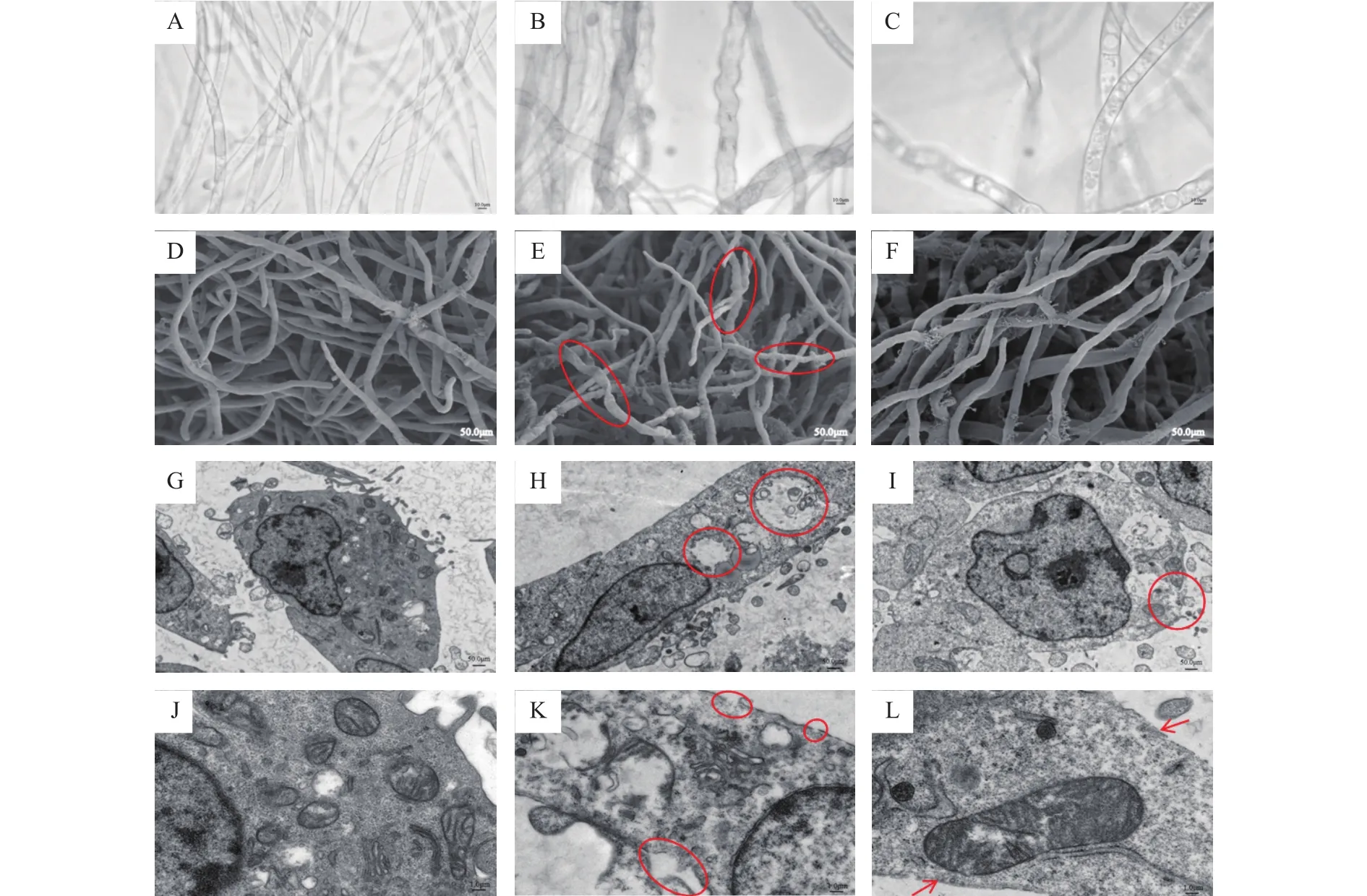
圖2 三峽肽素對指狀青霉的形態變化影響Fig.2 Morphological responses of P.digitatum induced by Sanxiapeptin
2.2 麥角固醇和山梨醇對三峽肽素介導的抑菌作用的補償作用
麥角固醇是真菌細胞膜的重要組成成分,該化合物的結構穩定,專一性強;山梨醇是一種滲透保護劑,可以用于穩定真菌的原生質體,這兩種物質在許多研究中分別用作細胞膜和細胞壁的保護劑[29]。為了明確三峽肽素抑制指狀青霉的可能靶點,本研究分析了麥角固醇和山梨醇在不同濃度下對其抑菌能力的影響。實驗結果表明:在低濃度下上述物質均沒有明顯的效果,但是1 mg/mL濃度下的山梨醇對三峽肽素的抑菌能力不僅沒有補償效應,反而會促進三峽肽素的抑菌能力(圖3A)。而麥角固醇則相反,在1 mg/mL的濃度下,會顯著(P<0.05)緩解三峽肽素對指狀青霉的抑制程度,表現出一種明顯的補償效應(圖3B)。相比正對照百可得,在同濃度下,山梨醇和麥角固醇均未對百可得的抑菌能力造成影響(圖3C~圖3D)。以上結果表明,三峽肽素與百可得的作用機理不一致,但可能通過影響細胞膜中麥角固醇的變化來達到抑制真菌的能力;同時,細胞壁及滲透壓的變化也會影響到三峽肽素的抑菌能力。
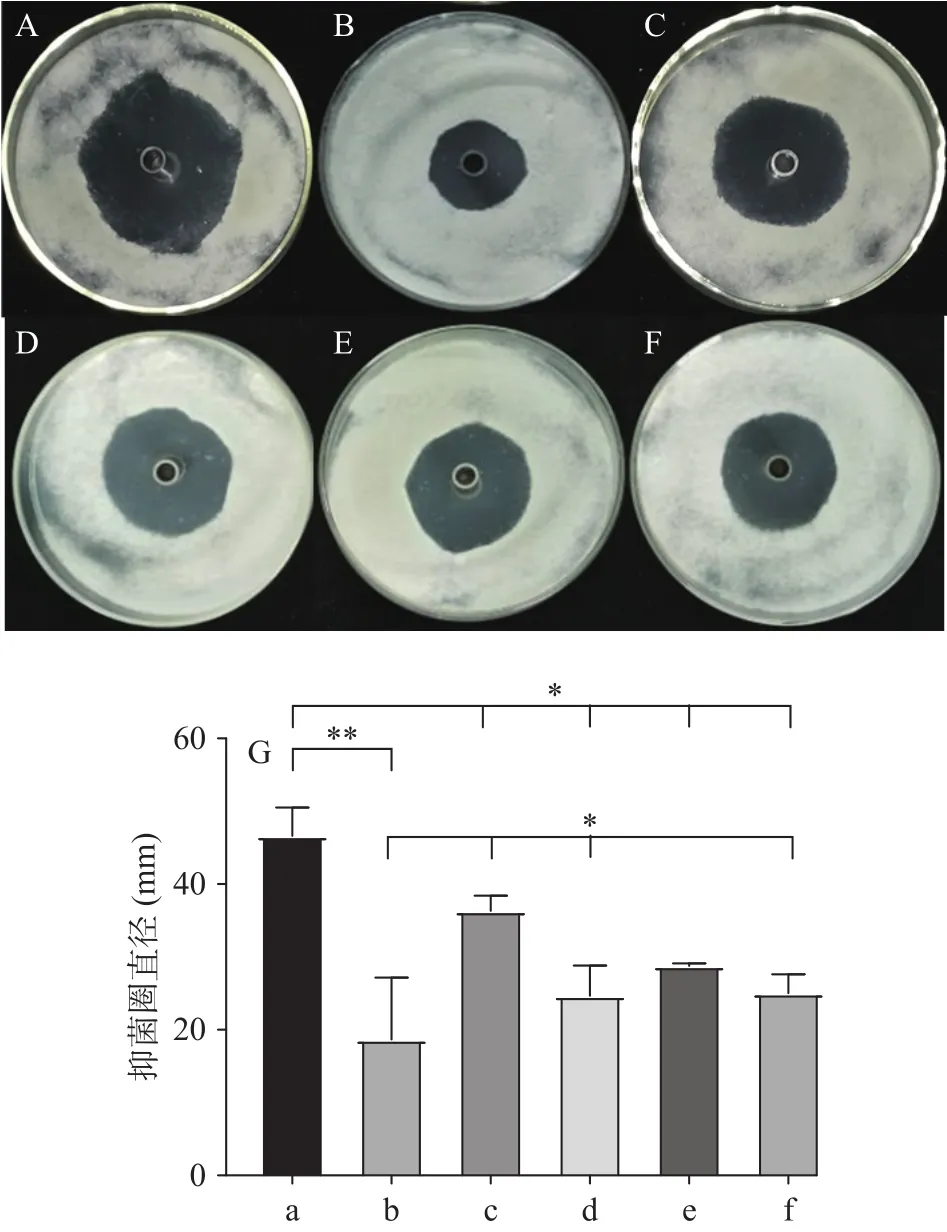
圖3 山梨醇和麥角固醇對三峽肽素抑菌能力的補償效應Fig.3 Compensatory effect of sorbitol and ergosterol on the antifungal ability of Sanxiapeptin
2.3 轉錄組測序質量分析
將測序產生的原始數據(raw reads)中低質量、接頭污染以及未知堿基含量過高的數據過濾后,平均每個菌株產生了4253萬條凈序列(clean reads),平均容量達6.38兆堿基(Gb),高質量數據約96%。將所獲得的的clean reads使用HISAT進行參考基因組序列比對,平均比對率達到93.66%。上述結果表明(表2),此次測序的質量有保證,結果可靠[30]。
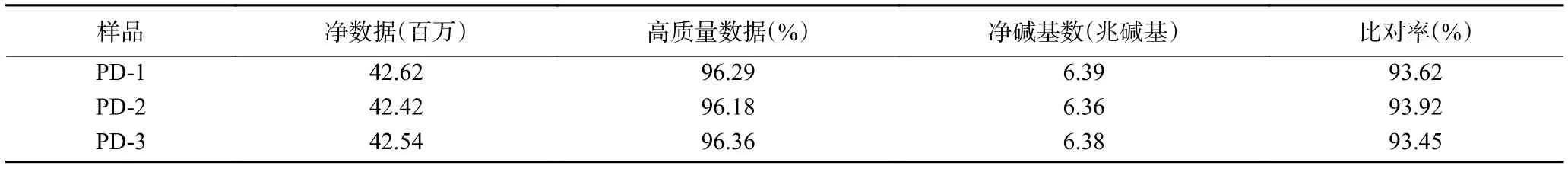
表2 轉錄組測序數據概況Table 2 Overview of transcriptome sequencing data
2.4 差異表達基因分析
為了進一步分析三峽肽素抑菌作用的分子機理,按照差異的倍數|log2(FoldChange)|和錯誤發現率(FDR)來篩選差異表達基因,分析中將同時滿足|log2(FoldChange)|≥1 且 FDR≤0.001 的轉錄本確定為差異表達基因[31]。三個轉錄組中的表達發生顯著變化的基因數如表3。
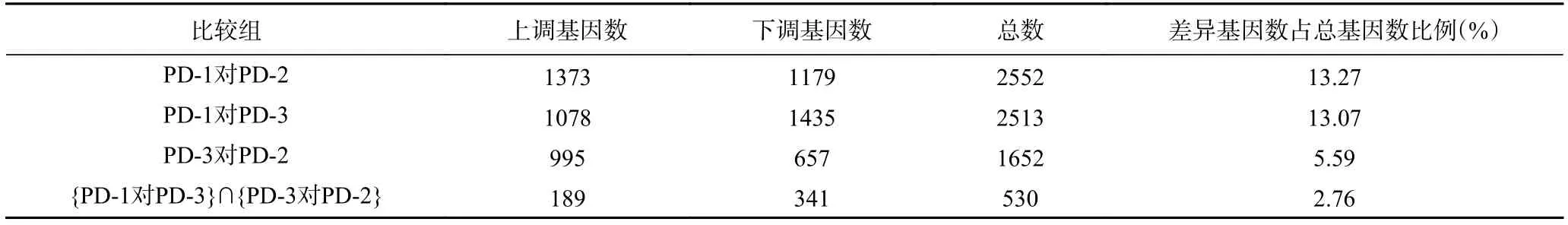
表3 差異表達的基因數Table 3 Number of differentially expressed genes
上述結果表明,加入三峽肽素處理后的菌株(PD-3)和正常生長的菌株(PD-1)相比,其差異表達的基因數有13.07%,該結果與百可得處理后的菌株結果非常接近,即PD-1對PD-2的差異基因為13.27%,從數量上反應二者作用的趨同性。但是這些差異表達的基因大多數并不相同(圖4),且由于二者作用后的顯微觀察結果及麥角固醇和山梨醇的補償實驗存在明顯差異,說明二者的作用途徑明顯不同,因此,我們對于差異基因的研究集中在兩者的交集({PD-1對PD-3}∩{PD-3對PD-2})包含的530個基因中,即聚焦在僅由三峽肽素引起的表達差異的基因上。
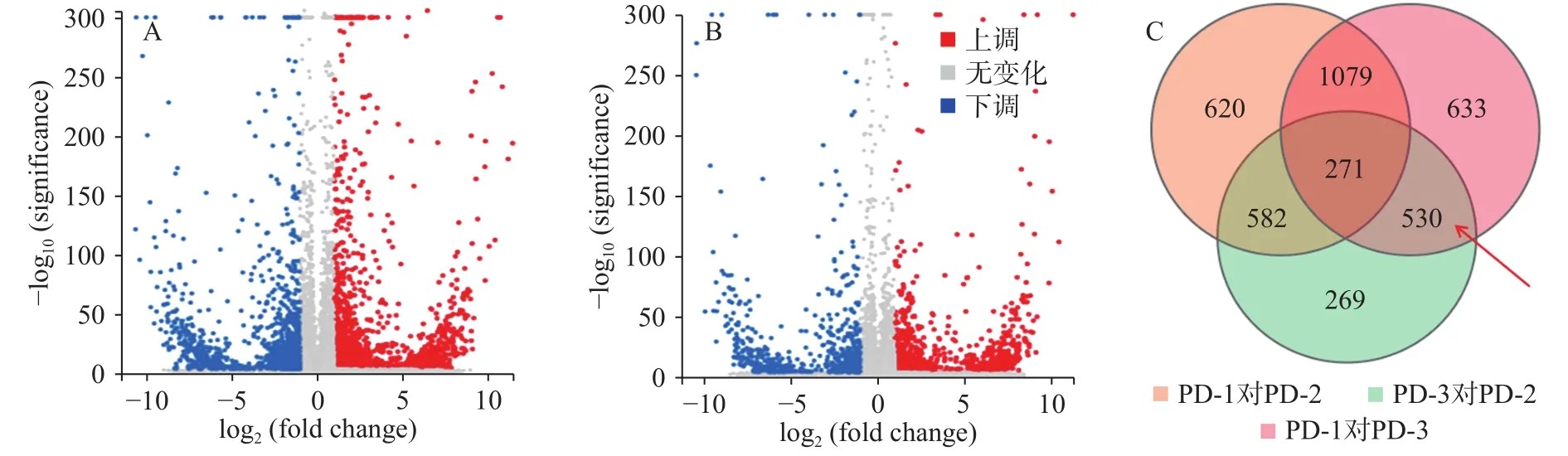
圖4 差異表達基因的火山圖分布Fig.4 Volcano figure of differentially expressed genes
2.5 差異表達基因的功能注釋
對不同處理下差異表達的基因,分別采用GO和KEGG分類富集分析。對照百可得處理后的指狀青霉(PD-2)和正常生長的菌株(PD-1)相比,糖類、脂質的代謝、能量代謝、遺傳過程和細胞自噬等途徑發生了變化(圖5A~圖5B)。三峽肽素處理后的菌株(PD-3),其差異基因也主要集中在上述途徑(圖5C~圖5D),不管是基因的數量還是基因可能的功能,暗示三峽肽素和百可得在抑制真菌的代謝通路上可能有部分重疊。
為了深入分析三峽肽素和百可得抑菌機理的差異,根據三峽肽素所導致的指狀青霉的形態變化以及相關保護劑的補償實驗結果,將僅由三峽肽素影響的530個基因進行了代謝通路分析,結果發現很多基因與聚糖、鞘脂類物質、固醇、疏水蛋白及GPI(糖基磷脂酰肌醇,Glycosylphosphatidylinositol)的代謝相關(圖5E~圖5F,表4),說明了三峽肽素有可能影響了細胞膜和細胞壁等細胞表面的功能。
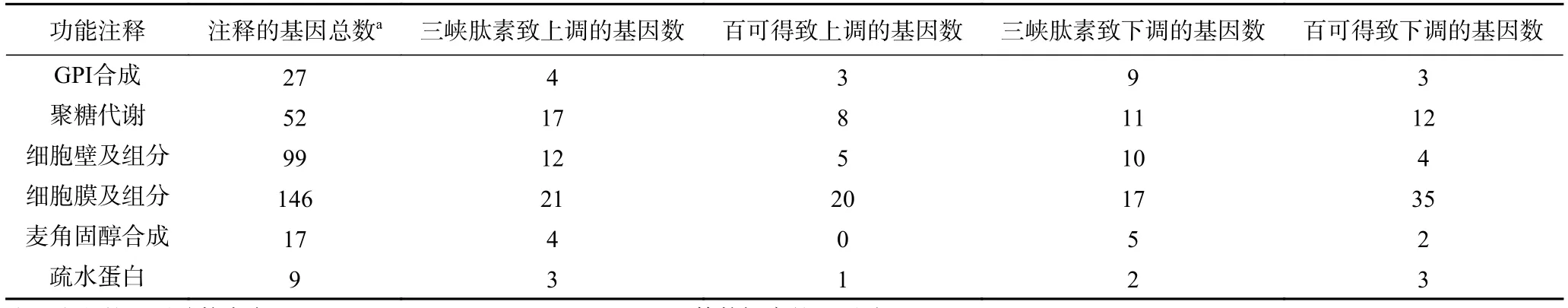
表4 部分可能參與細胞表面功能的差異表達的基因Table 4 Some differentially expressed genes possibly involved in cell surface metabolism
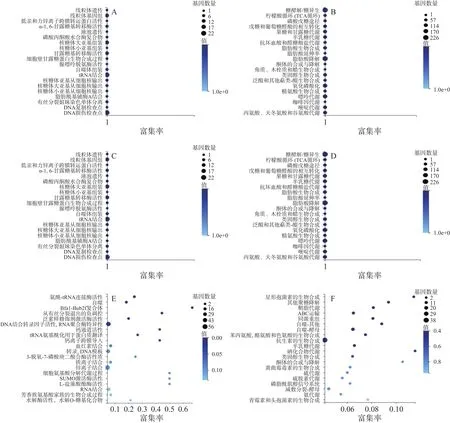
圖5 差異表達基因的注釋分析Fig.5 Annotation of differentially expressed genes
2.6 差異表達基因的驗證分析
為了驗證上述轉錄組測序分析的準確性,本研究選取了三峽肽素影響指狀青霉表達變化的17個候選基因(表5)進行RT-PCR半定量表達驗證,這些基因的功能大多與細胞表面的代謝相關。在進行RT-PCR之前,先選擇P450基因CL1226.Contig2作為管家基因,因為它在不同時期、不同條件下均可穩定的表達,是一種較為理想的內參基因[32-33]。
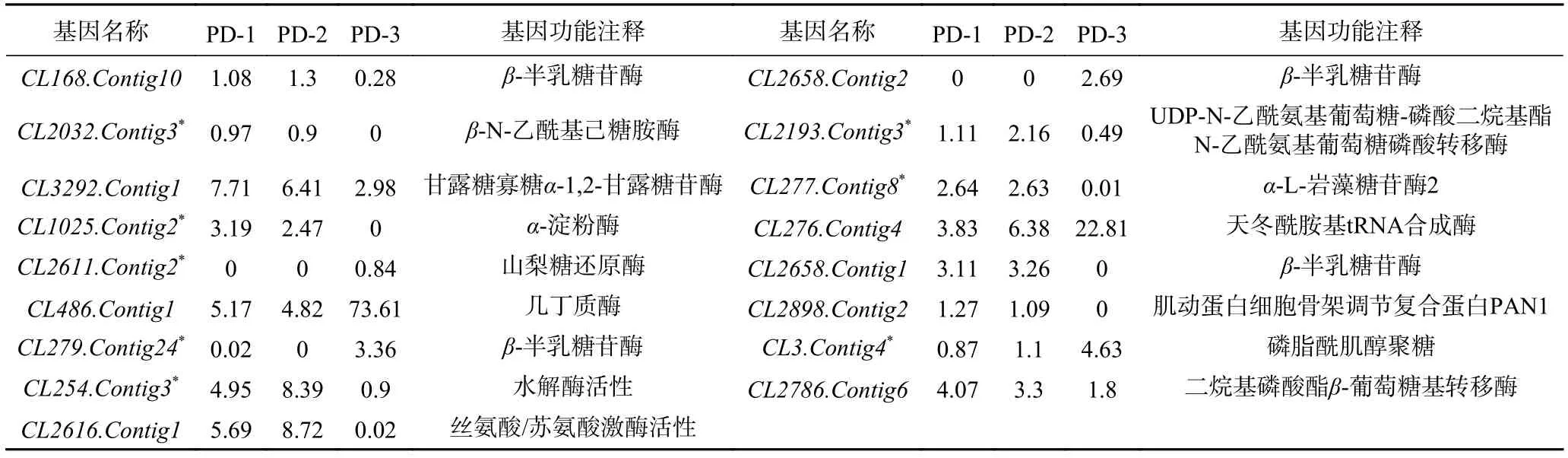
表5 指狀青霉中候選基因的表達量及注釋Table 5 Expression and annotation of candidate genes in P.digitatum
半定量RT-PCR結果顯示,有8個參與細胞表面代謝的基因CL254.Contig3、CL279.Contig24、CL1025.Contig2、CL2611.Contig2、CL3.Contig4、CL2032.Contig3、CL2193.Contig3、CL277.Contig8基因的表達與轉錄組測序結果完全一致(表5、圖6)。功能預測結果表明,這些基因大部分位于細胞壁或細胞膜中,可能與細胞表面的磷脂酰肌醇聚糖的合成和代謝的生物活動相關,從而影響了細胞表面的完整性,導致細胞內溶物外溢,從而起到抑菌的作用。
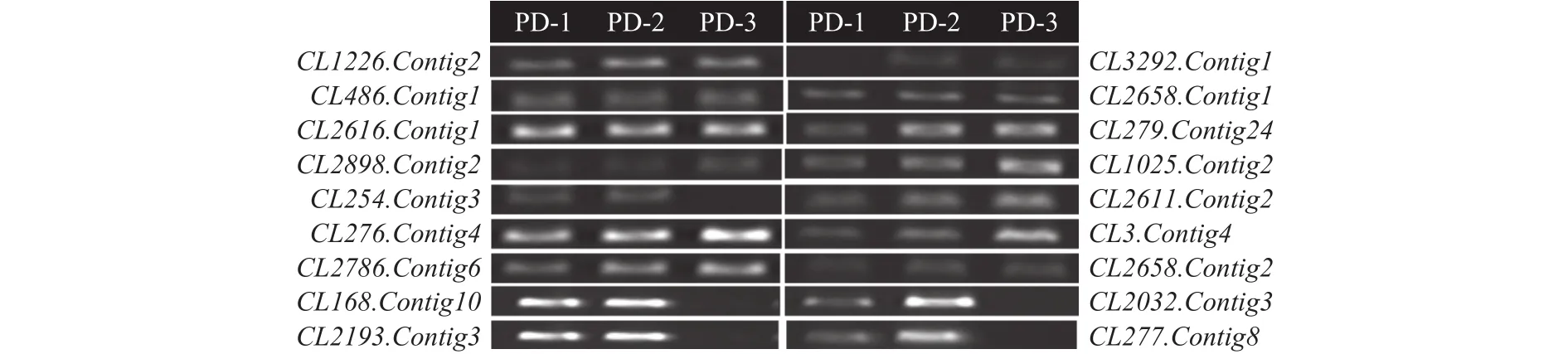
圖6 候選基因的RT-PCR表達驗證Fig.6 RT-PCR expression verification of the candidate genes
3 結論
本研究分析了三峽肽素對指狀青霉的形態學影響,明確了細胞膜保護劑麥角固醇及細胞壁保護劑山梨醇對三峽肽素抑菌作用的補償效應,同時基于BGISEQ-500測序平臺,分析比較了指狀青霉在三峽肽素處理后的無參轉錄組測序數據,并對差異表達基因進行了分析,將可能的抑菌代謝途徑初步聚焦在530個差異基因中。重點對影響細胞表面的候選基因進行了分析和驗證,初步確定三峽肽素通過影響疏水蛋白、麥角固醇合成酶、聚糖及GPI代謝途徑相關的蛋白等細胞膜及細胞壁等細胞表面的功能蛋白來發揮作用,所有這些嘗試為明確其抑菌真菌的機理奠定了基礎。后續工作的主要內容將集中在對上述候選基因在草酸青霉中進行超表達或者基因敲除以構建突變體,在明確基因功能的基礎上,通過分析突變體對三峽肽素敏感程度的變化解析其作用機理,進一步確定三峽肽素的作用靶點。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
