不同氮形態處理條件下楊樹根尖差異表達基因的特征分析
2022-03-25 08:46:50李卓蓉吳江婷
林業科學研究 2022年2期
關鍵詞:差異
周 婧,李卓蓉,吳江婷
(林木遺傳育種國家重點實驗室,國家林業和草原局森林培育重點實驗室,中國林業科學研究院林業研究所,北京 100091)
氮素是農林植物生長發育必不可少的大量元素之一,也是農林植物根系從土壤中吸收最多的礦質元素,對其生長發育具有重要意義[1-2]。但在土壤中無機氮含量往往較低[3],從而導致農林植物生產力低下,限制了其經濟效益的發揮。大量含硝態氮或銨態氮無機氮肥的施用,成為維持農林土壤肥力的主要手段之一。但盲目、過量施肥往往造成土壤養分失衡、氮素流失等諸多環境問題[4]。因此,深入挖掘農林植物響應氮素的關鍵基因,解析其分子調控機制,提高農林植物對氮素吸收同化能力,既可以實現農林生態系統氮素高效利用的經濟效益,也能實現減少土壤氮肥施加,保護生態環境的環保效益。
楊樹(PopulusL.)是我國主要的速生豐產用材林樹種之一[5]。由于楊樹生長迅速、對養分消耗大,對氮素有著很強的需求[3]。已有研究表明,不同氮形態處理,能夠導致小黑楊(P.simonii×P.nigra)根系形態發生改變,從而影響其對氮素的吸收同化能力[6]。作者前期研究表明,硝態氮處理,能夠改變灰楊(Populus × canescens)根尖形態,并影響根尖不同區段對硝態氮的吸收速率[7]。上述研究均表明,楊樹根尖能夠通過響應不同氮形態,改變其自身形態特征,從而影響根尖對不同氮形態的吸收同化能力[6-8]。
近年來,隨著高通量測序技術的快速發展,轉錄組測序成為研究基因差異表達的重要手段。利用高通量測序技術對楊樹根尖響應氮素的研究也取得了顯著成果,但是研究大多集中在氮素虧缺方面[9-10]。例如,通過轉錄組重測序以及生物信息學分析表明,低氮處理導致灰楊根系中特異表達的PtaNAC1(NAC-domain protein)上調表達,且增加了楊樹根系生物量,并顯著改變了PtaNAC1下游基因的表達,從而影響楊樹根系形態結構[9]。Dash 等的研究也表明,低氮處理條件下,在灰楊根系中,PtaHWS(Hawaiian Skirt),PtaNAC1和PtaRAP 2.11(subfamilies of ERF/AP2 TF family)顯著差異表達,并影響楊樹根系形態建成[10]。上述研究均通過轉錄組學和生物信息學方法,對楊樹根系響應低氮脅迫的過程進行了研究。而不同氮形態處理條件下,楊樹根尖存在怎樣的差異表達模式尚不清楚,值得深入研究。
本研究以灰楊根尖為試材,利用Illumina 測序平臺,篩選不同氮形態處理條件下,楊樹根尖差異表達基因,并結合生物信息學分析,構建楊樹根尖對不同氮形態響應過程的互作調控網絡,挖掘關鍵基因。最終,闡明楊樹根尖響應硝態氮或銨態氮,從而影響根尖生長發育過程的分子調控機制。研究成果為后續開發高氮素吸收利用效率的楊樹新種質提供了科學依據。
1 材料和方法
1.1 試驗材料
將生根的灰楊組培苗(4 周)轉移至水培,隔天換水澆灌改良的LA(Long Ashton)營養液(0.5 mmol·L-1NH4NO3、0.5 mmol·L-1KCl、0.9 mmol·L-1CaCl2、0.3 mmol·L-1MgSO4、0.6 mmol·L-1KH2PO4、42 μmol·L-1K2HPO4、10 μmol·L-1Fe-EDTA、2 μmol·L-1MnSO4、10 μmol·L-1H3BO3、7 μmol·L-1Na2MoO4、0.05 μmol·L-1CoSO4、0.2 μmol·L-1ZnSO4和 0.2 μmol·L-1CuSO4,pH 5.5)。2 周后,選取具有相似高度的48 株植株, 成2 組。分別添加0.5 mmol·L-1硝態氮(NO3-)和0.5 mmol·L-1銨態氮(NH4+)代替LA 營養液中的NH4NO3,進行水培,培養時間為10 d[7]。對根系形態特征進行分析。同時,收獲根尖0~4 cm樣品,迅速置于液氮中,放于-80℃冰箱備用。將8 株植物的樣本等量混合作為1 個重復,每種處理水平3 個生物學重復。
1.2 試驗方法
1.2.1 根尖形態特征分析 試驗苗進行不同氮形態處理10 d 后,對楊樹根系構型進行觀察測量,包括測量主根長度,以及測量可見側根距根尖的距離。每個處理水平測量8 株植株,且每種處理水平3 個生物學重復。進一步,對數據統計進行分析,利用Statgraphics 軟件,先對數據進行正態性檢驗,采用單因素方差分析(ANOVA),以P<0.05 作為統計意義上的顯著水平。
1.2.2 不同氮形態處理條件下轉錄組測序mRNAs 測序文庫構建和測序按照Illumina 公司提供的標準步驟執行。將上述收獲的2 種不同氮形態處理條件下的根尖(0~4 cm)樣品,每個處理3個重復,分別利用RNA 提取試劑盒提取總RNAs(TRK1001,聯川生物技術公司,杭州,中國)。將提取的總RNAs 經過DNase I 消化后,富集含有polyA 尾的mRNA,反轉錄成cDNA。合成第二鏈cDNA 后,用QIA quick PCR extraction 試劑盒進行純化。經過回收純化、PCR 富集后,獲得cDNA文庫。將構建好的文庫,利用杭州聯川生物技術公司IlluminaHiSeqTM4 000 高通量測序技術,進行轉錄組測序分析。
1.2.3 轉錄組測序結果分析 將測得的RNAs 序列,使用聯川生物技術公司開發的軟件進行分析。首先,將得到的原始序列進行去雜質處理,包括接頭序列的讀取片段、質量值Q ≤ 5 的低質量序列讀取片段和未知堿基數比例>10%的序列讀取片段。經過篩選得到的序列讀取片段使用SOAPaligner/soap2 將測序結果比對到灰楊基因組數據庫(http://aspendb.uga.edu/index.php/databases/spta-717-genome)[11]。采用FPKM(Fragments Per Kilobase of exon per Million mapped)對基因的表達水平進行定量[12]。基于FPKM 值,使用Ballgown package計算mRNAs 的差異表達水平。利用mRNAs 在硝態氮處理條件下的FPKM 除以銨態氮處理條件下FPKM 來計算基因的差異倍數(Fold change,FC)。差異表達mRNAs 篩選閾值為log2(FC)≥1 或≤ -1,且P< 0.05。
1.2.4 熒光定量PCR 引物設計及合成 以上述提取的不同氮形態處理楊樹根尖樣品為模板,使用Takara 公司PrimeScriptTMRT reagent Kit with gDNA Eraser 試劑盒反轉錄合成cDNA 第一鏈,作為熒光定量PCR 反應的模板。從轉錄組數據中選取10 個差異表達的基因,根據灰楊數據庫,利用Primer Premier 3.0 軟件設計引物,并由上海生工生物工程有限公司合成相關引物。Actin為RTqPCR 的內參基因。序列見表1。使用SYBR Premix Ex TaqTM熒光定量試劑盒進行熒光定量檢測。每個樣品進行3 次重復。將qPCR 得到的Ct 值進行歸一化,計算差異表達基因的相對表達量[7]。

表1 RT-qPCR 引物序列Table 1 Primers used for RT-qPCR
1.2.5 差異基因的功能注釋與富集分析 通過Blast2GO 軟件,對所有差異基因進行GO 功能注釋[13]。隨后進行GO 顯著性富集分析,以上述得到目的GO 條目(term)為單位,通過P值Bonferroni校正(P< 0.05)來定義差異表達基因中顯著富集的GO term。同時,結合生物信息學KEGG 軟件,分析差異表達基因的生物學功能[14]。同樣以correctedP< 0.05 為標準,篩選在差異表達基因中顯著富集的代謝途徑。利用MapMan 軟件、Popgenie 數據庫(https://popgenie.org/exnet)以及Cytoscape程序,構建楊樹根尖響應不同氮形態相關差異表達基因的互作調控網絡[15]。
2 結果與分析
2.1 根系形態特征分析
在不同氮形態處理條件下,灰楊根系形態特征具有顯著差別(圖1)。硝態氮處理下的主根長度幾乎是銨態氮處理條件下的一倍,這說明不同氮形態處理影響楊樹根系生長發育過程。同時,在兩種處理水平下,能夠發揮吸收營養和水分作用的可見側根發生的位置相對一致。因此選取側根形成前的0~4 cm 根尖部分進行后續研究。

圖1 不同氮形態處理后,灰楊根系表型Fig.1 Morphological parameters of P. × canescens roots with different nitrogen forms
2.2 轉錄組測序數據分析
基于上述形態特征分析的結果,以不同氮形態處理楊樹根尖為材料,進行高通量轉錄組測序分析。測序結果顯示,兩個處理水平的測序分別獲得57 001 409 和57 517 359 條有效序列(表2),占各自文庫的比例均超過了99.20%(表2)。兩個處理水平的測序數據,能夠比對到灰楊數據庫中的堿基數比例分別為81.59%和82.60%,說明測序所得序列與灰楊數據庫比對性較好,可以用于后續分析。

表2 轉錄組文庫測序數據統計Table 2 Distribution of RNA-seq in different categories
2.3 不同氮形態處理條件下,楊樹根尖差異表達基因分析
在不同氮形態處理條件下,以銨態氮處理水平為對照,在硝態氮處理條件下,共篩選出2 207 個差異表達基因(p< 0.05)。相比較于銨態氮處理,硝態氮處理有1 414 個基因上調表達,793 個基因下調表達。有32 個基因顯著差異表達(p< 0.001),見圖2a。其中,Potri.013G102100基因差異變化最顯著,log2(FC)為-6.56,該基因注釋為II 型過氧化物酶(peroxiredoxin type 2);Potri.004G138100基因上調倍數最顯著,log2(FC)為6.18,然而該基因尚未被注釋,功能未知。同時,篩選出71 個與植物生長發育相關的差異表達基因。其中,轉錄因子作為一大類調控基因,在楊樹根尖通過響應不同氮形態而參與其生長發育過程中發揮著重要作用。例如,NFYA(nuclear transcription factor Y subunit alpha)、GRF(growthregulating factor)和ARF(auxin response factor)類轉錄因子,它們均能夠通過調控硝態氮轉運體NRT(nitrate transporters)基因的表達響應不同氮形態,從而調節植物根系生長發育過程[16-17]。進一步,挑選了10 個顯著差異表達基因,利用RTqPCR 進一步證實了測序結果的可靠性(圖2b)。
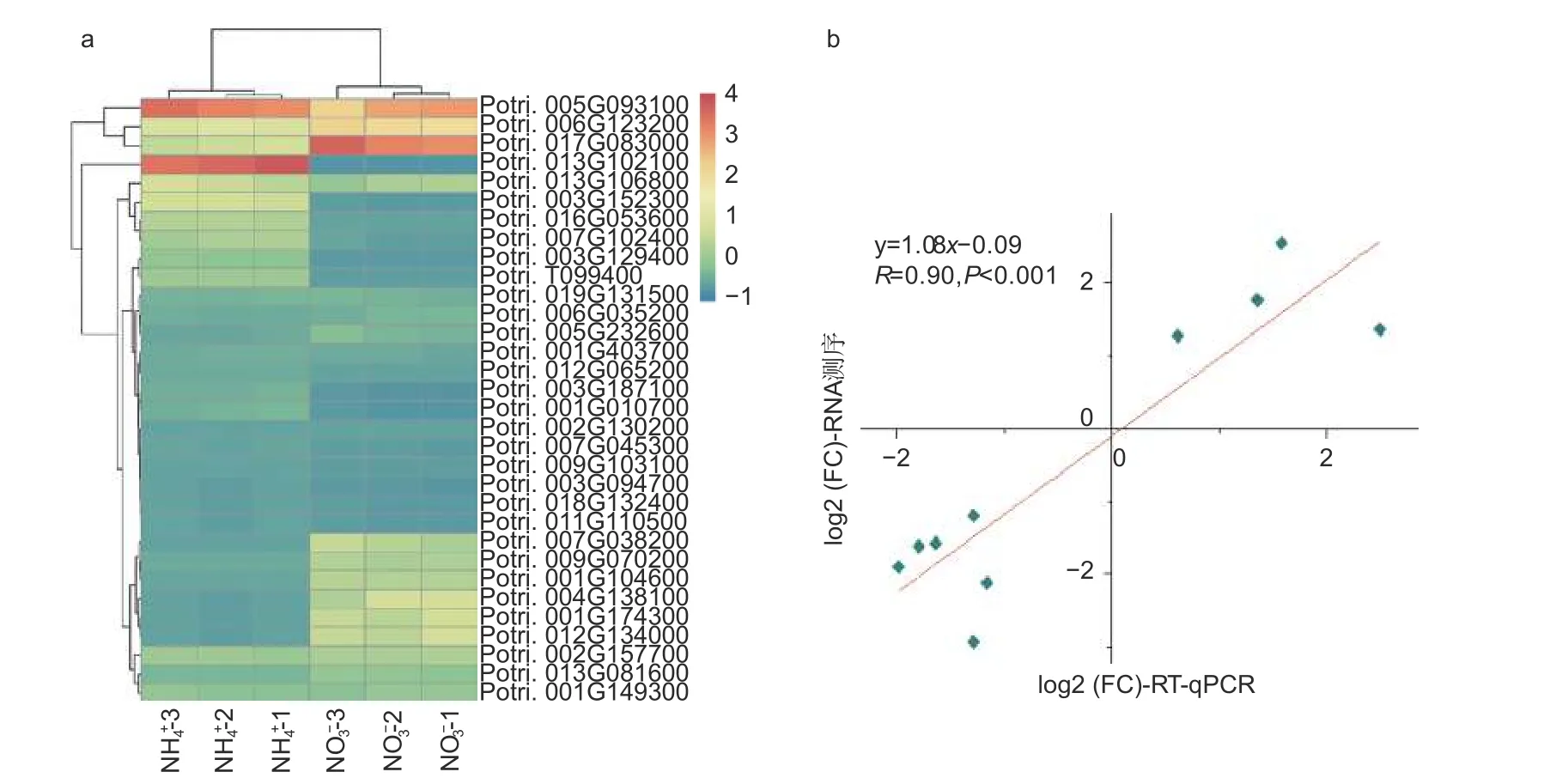
圖2 不同氮形態處理條件下,楊樹根尖顯著差異表達的基因Fig.2 Significantly differentially expressed mRNAs in root tips of P. × canescens under different nitrogen form treatments
2.4 不同氮形態處理條件下,灰楊根尖差異表達基因的GO 功能劃分
為進一步了解差異表達基因的功能,將差異表達基因映射GO 數據庫,根據序列同源性,將不同氮形態處理條件下,2 207 個差異表達基因分為3 大類總共50 個功能組,包括生物學過程(biological process,BP)、細胞組分(cellular component,CC)和分子功能(molecular function,MF)(圖3)。結果顯示,生物學過程主要集中在轉錄調控、刺激反應和代謝過程。在細胞組分類別中,差異基因被注釋到最多的亞類依次為細胞核、質膜和細胞質部分。在分子功能中,注釋為ATP 結合和蛋白結合的差異基因最多,其次為轉錄因子活性和DNA 結合。以上注釋較多的過程所涉及到的差異基因,可能參與楊樹根尖對不同氮形態的響應,從而影響根尖生長發育過程。進一步挖掘這些基因的功能,研究其響應不同氮形態的作用機制,將有助于為培育高氮素吸收利用效率的楊樹新種質奠定基礎。
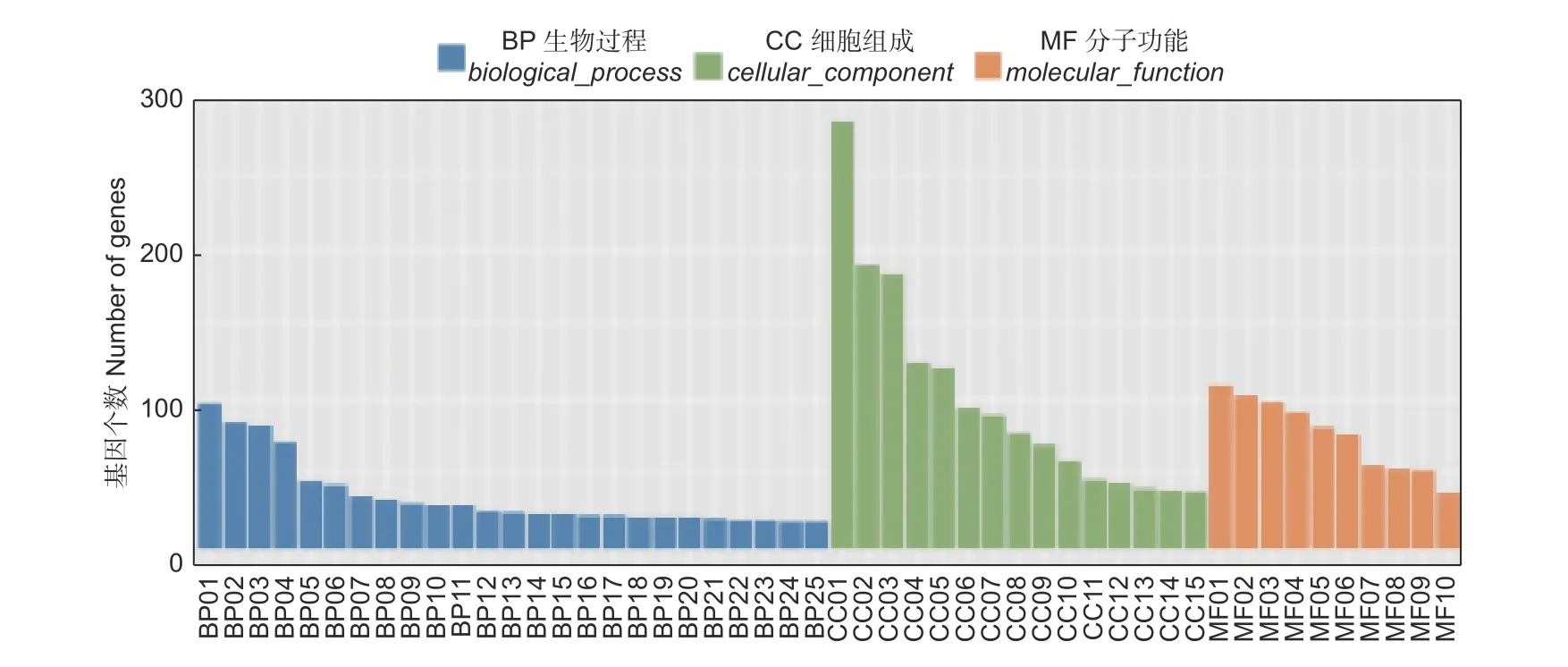
圖3 不同氮形態處理條件下,灰楊根尖差異表達基因的GO 功能聚類注釋Fig.3 Gene Ontology classification annotation of differentially expressed genes in root tips of P. × canescens under different nitrogen form treatments
2.5 不同氮形態處理條件下,灰楊根尖差異表達基因的KEGG 分析
為進一步了解上述差異表達基因的功能,對識別出的差異表達基因進行KEGG 通路分析。共獲得1 192 個差異表達基因,涉及到20 條KEGG 通路(圖4)。其中,有9 條與代謝相關,并且大多與氮代謝和氨基酸代謝相關,有4 條與生物合成相關。KEGG 分析從功能的角度鎖定基因,直觀的顯示了楊樹根尖響應不同氮形態差異基因的代謝過程,有助于更好地研究楊樹根尖響應不同氮形態的分子機制。
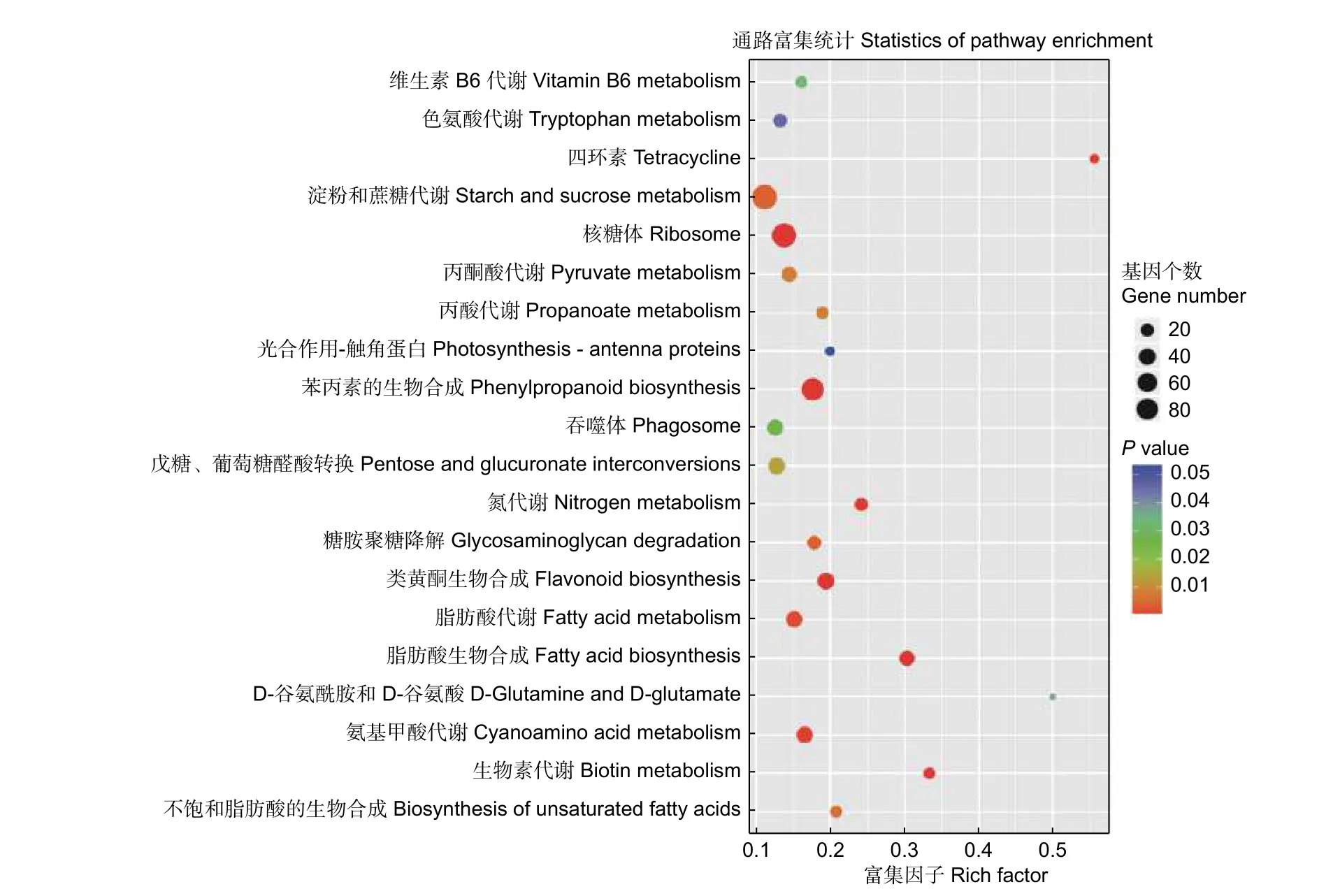
圖4 KEGG 通路分析鑒定差異表達基因Fig.4 KEGG pathway analysis of significantly differentially expressed target genes
2.6 與氮代謝相關差異表達基因的MapMan分析
利用MapMan 軟件,篩選出36 個與氮代謝通路過程、各類氨基酸的生物合成以及代謝過程相關的差異表達基因(表3)。其中,有8 個基因參與到氮代謝過程,26 個基因參與到氨基酸代謝與合成過程,還有2 個基因參與到硝態氮轉運過程。已有研究表明,硝態氮處理能夠改變NRTs 的表達水平,進一步影響植物根尖生長發育過程[18-19]。
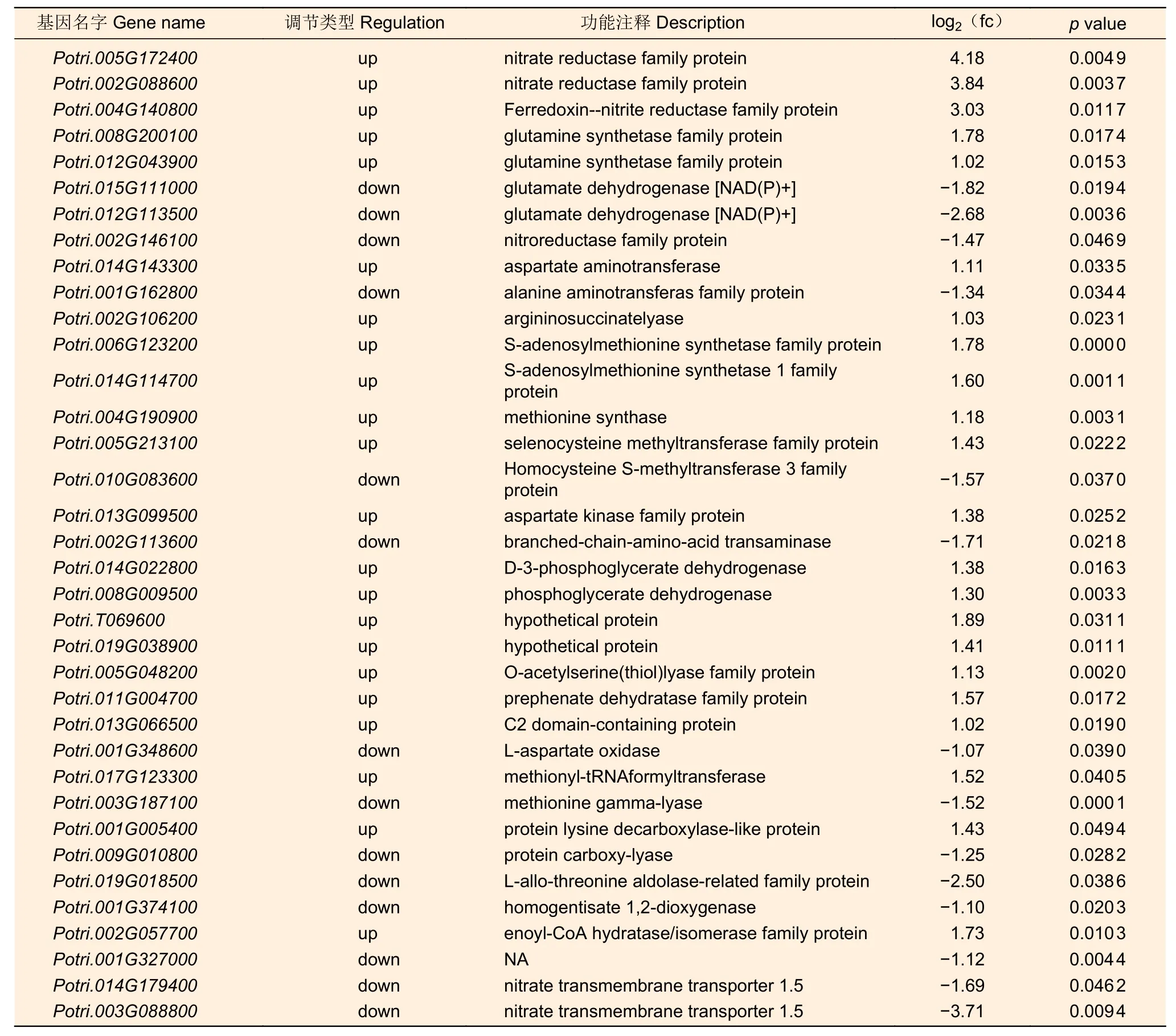
表3 氮代謝相關差異表達基因Table 3 Differentially expressed genes related to nitrogen metabolism
2.7 不同氮形態處理條件下,灰楊根尖差異表達基因的互作網絡分析
為了進一步研究這些基因的功能,利用Popgenie數據庫以及Cytoscape 程序,構建了灰楊根尖對不同氮形態響應過程的互作調控網絡。在基因作用網絡中,檢測到一組相互作用的與氮代謝過程相關的基因簇,共包含13 個基因(圖5)。其中,硝酸還原酶(Potri.005G172400)基因作為硝態氮同化過程中的關鍵酶,占據了網絡的核心位置。已有研究表明,硝酸還原酶作為植物氮素同化的關鍵酶之一,能夠調控根系生長素水平,從而影響根尖生長發育過程[20-21]。進一步,挑選了調控網絡中7 個顯著差異表達基因,選取不同氮形態處理10 天的灰楊根尖材料進行RT-qPCR 實驗。結果顯示:這7 個基因在RT-qPCR 的結果與轉錄組分析的趨勢一致(圖6)。
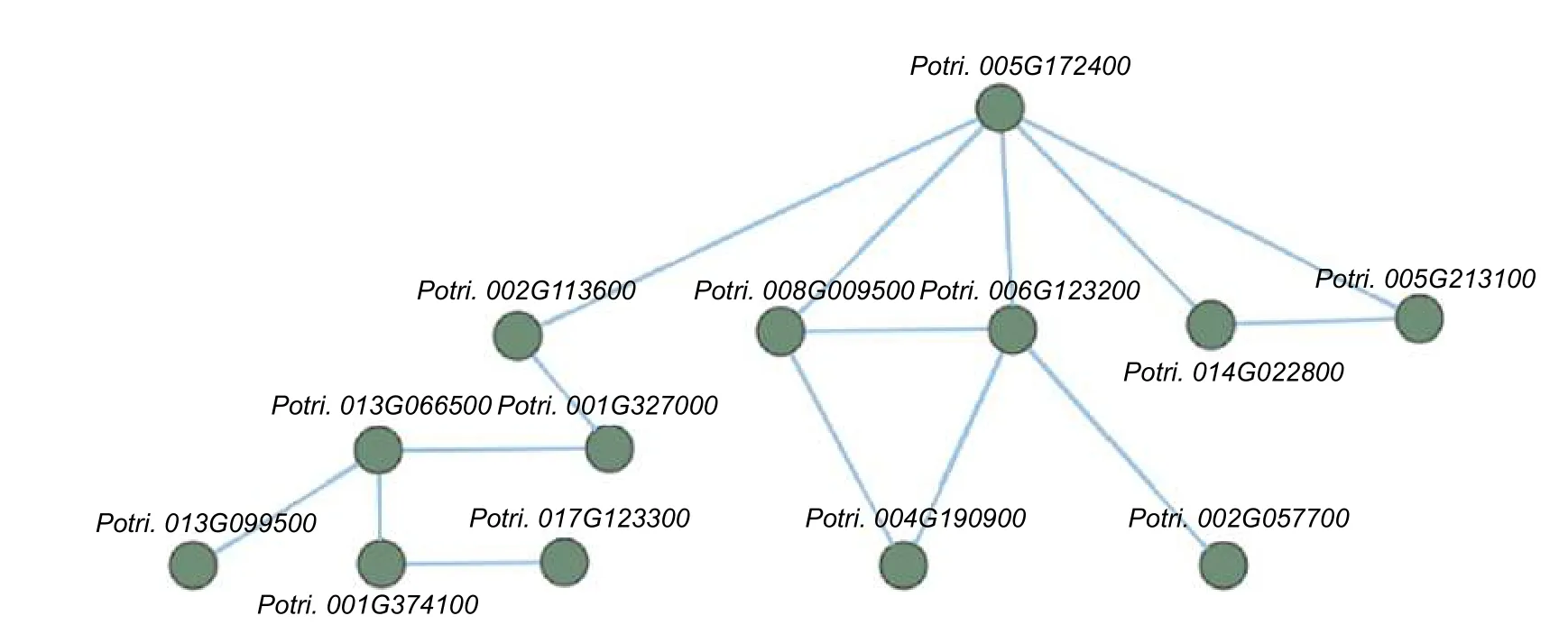
圖5 不同氮形態處理條件下,灰楊根尖差異表達基因的互作網絡Fig.5 The interaction network of differentially expressed genes in root tips of P. × canescens under different nitrogen forms treatments

圖6 不同氮形態處理條件下,灰楊根尖差異基因表達分析Fig.6 Validation of significantly differentially expressed genes under different nitrogen forms treatments by sRNA-seq and RT-qPCR
3 討論
氮素是植物生長發育所必需的大量礦質營養元素之一,是蛋白質、核酸、葉綠素等重要物質的組成成分[22]。土壤中的氮素都需要在適宜的溫度、水分和通氣條件下,在土壤微生物和酶的作用下,將其水解或氧化為硝態氮或銨態氮,才能夠被植物根系直接吸收利用[23-24]。而土壤中的不同氮形態可能導致楊樹根尖氮代謝相關基因表達模式發生改變,從而影響植物根尖生長和發育過程[25]。本研究在不同氮形態處理10 d 后,對楊樹根尖進行觀察發現,硝態氮處理下的主根長度幾乎是銨態氮處理條件下的一倍。類似的現象在小黑楊根系中(P.simonii×P.nigra)也有報道[6]。為了進一步了解木本植物根尖對不同氮形態響應的過程,本研究利用轉錄組測序技術,對不同氮形態處理條件下,楊樹根尖差異表達基因進行了分析。
在不同氮形態處理10 d 后,經過高通量測序分析,獲得了2 207 個差異表達基因。其中,Potri.013G102100基因編碼一個II 型過氧化物酶(peroxiredoxin type 2),其下調倍數最為顯著,前人研究表明,II 型過氧化物酶是植物中分布最廣的一類非典型2-半胱氨酸過氧化物酶(2-Cysteine Peroxiredoxin),其在對抗氧化能力和共生固氮中發揮著重要作用[26]。另有研究表明,2-半胱氨酸過氧化物酶可以與氮同化過程中亞硝酸還原酶(nitrite reductase)和谷氨酸合成酶(glutamate synthase)相互作用,從而影響氮代謝過程[27]。Potri.004G138100基因上調倍數最顯著,但該基因功能未知,推測其可能參與楊樹根尖對不同氮形態的響應,從而影響楊樹根尖生長發育過程。同時,在本實驗中,部分差異表達轉錄因子通過響應不同氮形態,影響楊樹根尖生長發育過程,這與在草本植物中的研究結果相似[6,8,28]。例如,在水稻(Oryza sativaL.)和小麥(Triticum aestivum)中,過表達NFYAs 能夠調控NRTs的表達[18,29]。同時,增加了轉基因小麥根尖對硝態氮的吸收,促進了側根生長[18]。在本研究中,相比較于銨態氮處理,硝態氮處理導致灰楊根尖PcNFYA7(Potri.011G 101000)下調表達,同時促進楊樹主根伸長。在擬南芥(Arabidopsis thaliana)根中柱鞘細胞中,5mM 硝態氮可以促進ARF6/8的表達水平,促進了擬南芥側根起始和隨后萌發的過程[17,30]。在本研究中,相比較于銨態氮處理,硝態氮處理導致灰楊根尖中PcARF8/16(Potri.004G078200和Potri.010G223200)上調表達,同時促進了主根伸長。上述研究表明,灰楊根尖部分差異表達的基因,能夠通過響應不同氮形態而參與調控根尖生長發育過程。
對差異基因進行GO 和KEGG 通路分析。結果表明,一些差異表達基因參與氮代謝和氨基酸代謝途徑。例如,氮代謝、色氨酸代謝、丙酮酸代謝和氨基甲酸代謝途徑等。同時,利用MapMan篩選出36 個氮代謝相關差異表達基因。其中,值得關注的是兩個硝態氮轉運體基因PcNRT1.5(Potri.014G179400和Potri.003G088800)。相比較于銨態氮處理,硝態氮處理導致灰楊根尖中PcNRT1.5顯著下調表達。在擬南芥中,AtNRT1.1作為第一個被鑒定出的NRT 在根中有轉運和感應硝態氮兩個作用[31]。同時,在小麥和水稻中,硝態氮處理均能改變NRT 基因的表達水平,從而影響植物根尖生長發育過程[18-19]。該結果說明PcNRT1.5在楊樹根尖響應不同氮形態過程中發揮著重要的調控作用。進一步,通過Popgenie 數據庫以及Cytoscape程序,構建了灰楊根尖響應不同氮形態的互作調控網絡。在基因作用網絡中,硝酸還原酶(Potri.005G 172400)作為硝態氮同化過程中第一步的關鍵酶,在楊樹根尖對不同氮形態響應過程中發揮著關鍵作用。這與在草本植物中的研究結果類似[20]。在擬南芥根中,硝酸還原酶作為氮素同化和根系構型重塑的關鍵酶之一,能夠調控根系生長素水平,從而影響根尖生長發育過程[20]。同時,在水稻中,硝酸還原酶生成的一氧化氮,通過誘導水稻側根形成和無機氮的吸收,提高水稻對氮的吸收能力[21]。而楊樹作為多年生木本植物,其根系比淺根系草本植物發達得多,且構型更為復雜[9]。同時,木本植物往往遭受季節性、物候性土壤氮素反復變化,面臨的土壤氮素環境復雜程度遠遠大于草本植物[4,32-33]。因此,楊樹根系中硝酸還原酶可能存在著更為復雜的氮適應性變化的作用機制,值得深入研究。同時,通過基因互作網絡的分析,可以逐步了解參與互作網絡的基因在響應不同氮形態過程中的作用,其相互作用關系也有待進一步確認。
4 結論
本研究在不同氮形態處理10 d 后,對楊樹根尖進行觀察發現,硝態氮處理下的主根長度幾乎是銨態氮處理條件下的一倍。同時,通過轉錄組測序分析,篩選出2 207 個差異表達基因。通過GO 功能和KEGG 通路分析,理清了差異表達基因富集的分子功能與代謝通路。利用MapMan篩選出36 個氮代謝相關差異表達基因,并構建了這些基因的互作網絡,得到了關鍵基因。綜上所述,本研究為挖掘和尋找楊樹根尖響應不同氮形態關鍵調控基因提供了參考,并闡述了這些關鍵基因可能通過響應不同氮形態,影響楊樹根尖生長發育過程,這為進一步培育出高效吸收利用氮素的楊樹新種質奠定了基礎。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
