凡納濱對蝦多代選育群體生長和耐綜合脅迫性狀的配合力及雜種優勢分析
2022-04-18 10:16:22王倫王崇懿劉建勇傅學麗
海洋學報 2022年4期
關鍵詞:生長
王倫,王崇懿,劉建勇,傅學麗*
( 1. 廣東海洋大學 水產學院,廣東 湛江 524088)
1 引言
凡納濱對蝦(Litopenaeus vannamei)又名南美白對蝦,具有生長快、抗逆能力強、適宜高密度養殖等優點,是世界三大對蝦養殖品種之一[1-4]。據統計,2020年我國凡納濱對蝦海水養殖產量達119.774×104t,占海水蝦類養殖總產量的80.50%以上,是我國最主要的海水對蝦養殖品種[5]。由于該品種并非我國的本地種,大多數親本種群是從國外進口或經過多代選育的親本種群[6-7],經過多代選育后有效種群規模較小,可能會帶來重要經濟性狀近交衰退的風險[8]。雜交是解決以上問題的有效方法之一[7],是創造變異的重要途徑;雜交可以產生親本從未表現的超親優勢(獲得雜種優勢)[9],從而顯著提高子代的生活力。因此,利用雜交將2個及2個以上不同遺傳背景品種的個體基因自由組合,創造出新的遺傳類型,經人工選擇優良親本,繼而培育出新品種。在凡納濱對蝦育種領域,我國雖已培育出多個新品種,但采用的方法多為選擇育種(家系選擇、群體選擇及品系選擇),采用雜交育成的新品種僅見“壬海1號”[10],導致對雜種優勢的利用遠遠不夠。
近年來,對蝦的放養密度不斷增加,但高密度養殖會降低水環境中的pH,增加總氮和氨氮濃度[11-12](對蝦高密度養殖氨氮濃度高達46 mg/L[13],pH低至4.1[14]);而夏季的高溫則會增加養殖水體中鹽度[15]。生活在較高鹽度、氨氮和較低pH水體中,對蝦會出現生長緩慢、免疫力下降、對病原菌的易感性提高等不良現象,致使養殖成活率降低[16-18]。凡納濱對蝦耐高鹽度、耐高氨氮和耐低pH的雜交育種遺傳改良是解決以上問題的重要途徑。
配合力分析是水產動物雜交育種研究中的重要工作之一,是遺傳改良的重要手段,也是雜種優勢利用的前提[19]。有關凡納濱對蝦生長或單因子脅迫的配合力研究已有相關報道,王浩等[20]研究了凡納濱對蝦7個引進群體體質量的配合力;胡志國等[21-22]報道了凡納濱對蝦耐鹽度和耐高氨氮脅迫的雜種優勢和配合力,分別選出耐高低鹽一般配合力(General Combining Ability, GCA)較高的泰國正大 (TZ)群體及耐高氨氮的強優勢組合(美國邁阿密SIS群體(UM)與美國夏威夷SIS群體(UH)的雜交組合,即(UM×UH)組合)群體。然而在凡納濱對蝦實際養殖生產中,其所受的脅迫常常為多個環境脅迫因子的綜合脅迫,但在凡納濱對蝦耐綜合脅迫(高鹽、低pH與高氨氮共同脅迫)配合力分析方面的研究未見報道,因此有必要開展該方面的研究,以實現育種目標的進一步提高。本研究以凡納濱對蝦“興海1號”選育核心群體與國外引進的不同遺傳背景的凡納濱對蝦群體交配,構建12個群體,以生長和耐受性狀為主要選育目標,探究凡納濱對蝦的生長和耐綜合脅迫的配合力及雜種優勢,以期為凡納濱對蝦生長和耐受性狀選育提供依據與參考。
2 材料與方法
2.1 實驗材料
2.1.1 實驗所用親蝦來源
凡納濱對蝦“興海1號”新品種核心群體(GX):以我國廣東湛江和廣西東興的7個凡納濱對蝦養殖群體(3 880尾)為基礎群體,以成活率和體質量為目標性狀,采用BLUP選育技術,從2011年開始經連續7代選育而成,并在2018年通過了全國水產原種和良種審定委員會審定。該品種的主要優點為:生長速度快、養殖存活率高(養殖100日齡的凡納濱對蝦“興海1號”平均成活率為77.80%,平均體質量為15.42 g,與SIS一代苗相比,平均成活率提高了15.0%)、抗逆性強(主要表現為抗高氨氮)。該品種核心群體經本課題組選育至今已達10年。
泰國群體(W和L):2019由泰國引進兩個多世代選育群體,分別為生長速度快的W品系及養殖適應性強,適合在高位池塘、普通蝦塘、大棚等多種養殖模式養殖的L品系。
美國群體(K和M):2019由美國引進兩個多世代選育群體,分別為兼具快長、養殖適應性強的K品系和養殖適應性強的M品系。
以上5個不同遺傳背景的凡納濱對蝦群體均經多世代選育而成,具有較高的遺傳純度。
2.1.2 家系構建和對蝦養殖
實驗于2020年3月在湛江市國興水產科技有限公司(國興公司)進行,按袁瑞鵬等[23]的方法強化親蝦,從每個群體中分別挑選雌、雄親蝦各20尾,進行交配實驗。本課題組選育的“興海1號”核心群體(GX)具有長速快、耐氨氮及養殖適應性強等優點,以其為父母本(續代選育)分別同W、L、K、M的母本和父本雜交,成功構建了GX♀×W♂、GX♀×K♂、GX♀×M♂及W♀×GX♂、K♀×GX♂;以GX、W、L、K、M群體分別為父母本進行自交(保留純系及提純),構建了GX♀×GX♂、W♀×W♂、L♀×L♂、K♀×K♂、M♀×M♂;以長速快的W和K群體分別與養殖適應性強的L和M群體雜交,成功構建了W♀×M♂、K♀×M♂,因構建家系較困難和育苗過程中的其他因素導致僅成功構建及保留12個子代組合(表1)。孵化后,選取每尾雌蝦孵化出的無節幼體約3 500尾,根據標準化育苗方式于500 L的桶中獨自育至仔蝦,而后移至獨立水泥池進行標粗培育。培育體長至3 cm左右時,每個交配群體隨機取900尾對蝦進行熒光標記,將所有標記好的對蝦置于長×寬×高為13 m×10 m×0.6 m的池內同環境養殖60 d。
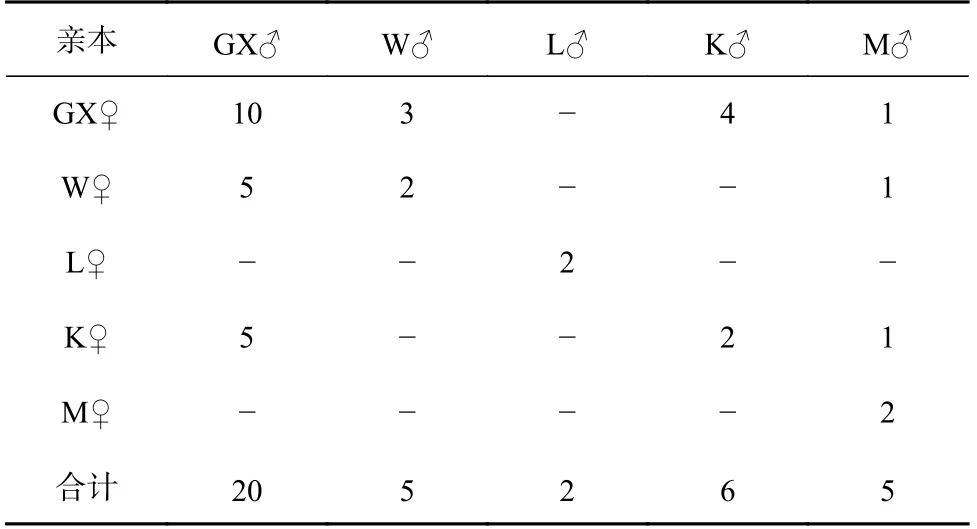
表1 凡納濱對蝦5個不同遺傳背景群體的不完全雙列雜交Table 1 Incomplete diallel crossing of five populations of Litopenaeus vannamei with different genetic backgrounds
2.2 高鹽、低pH和高氨氮綜合脅迫實驗
2019年由國興公司對凡納濱對蝦養殖土塘塘底水質指標進行監測,結果顯示:鹽度上限值為35,pH下限值為6,參照袁瑞鵬[24]的方法開展高氨氮脅迫預實驗,確定同環境養殖60 d,48 h高氨氮脅迫半致死濃度(LC50%)為120 mg/L(銨根離子)。綜合脅迫濃度的確定方式為控制鹽度為35、pH為6±0.1,改變氨氮濃度(可變范圍為1~120 mg/L),當半致死時間(LT50%)為48 h時對應的鹽度、pH及氨氮濃度即為最終的綜合脅迫濃度。
綜合脅迫預實驗:對蝦同環境養殖60 d后,設置高鹽為35、低pH為6±0.1,將高氨氮脅迫48 h的LC50%(120 mg/L)折中(60 mg/L),往1 mg/L或120 mg/L方向以間隔1 mg/L為梯度依次遞增或遞減,可遞分為120個梯度,1次實驗難以完成,因此,分40個梯度為1個大組(共3個組)。第1組為(1)由60 mg/L遞減的20個濃度(鹽度、pH和氨氮濃度)梯度分別為:35、6±0.1和60 mg/L;35、6±0.1和59 mg/L;35、6±0.1和58 mg/L;35、6±0.1和57 mg/L;35、6±0.1和56 mg/L;35、6±0.1和55 mg/L;35、6±0.1和54 mg/L;35、6±0.1和53 mg/L;35、6±0.1和52 mg/L;35、6±0.1和51 mg/L;35、6±0.1和50 mg/L;35、6±0.1和49 mg/L;35、6±0.1和48 mg/L;35、6±0.1和47 mg/L;35、6±0.1和46 mg/L;35、6±0.1和45 mg/L;35、6±0.1和44 mg/L;35、6±0.1和43 mg/L;35、6±0.1和42 mg/L;35、6±0.1和41 mg/L;35、6±0.1和40 mg/L;(2)由60 mg/L遞增的20個濃度梯度分別為:35、6±0.1和61 mg/L;35、6±0.1和62 mg/L;35、6±0.1和63 mg/L;35、6±0.1和64 mg/L;35、6±0.1和65 mg/L;35、6±0.1和66 mg/L;35、6±0.1和67 mg/L;35、6±0.1和68 mg/L;35、6±0.1和69 mg/L;35、6±0.1和70 mg/L;35、6±0.1和71 mg/L;35、6±0.1和72 mg/L;35、6±0.1和73 mg/L;35、6±0.1和74 mg/L;35、6±0.1和75 mg/L;35、6±0.1和76 mg/L;35、6±0.1和77 mg/L;35、6±0.1和78 mg/L;35、6±0.1和79 mg/L;35、6±0.1和80 mg/L。其他兩個組以此類推分別繼續往前和往后遞減和遞增。3組預實驗分開進行,先分別對第1組的40個梯度開展綜合脅迫實驗。調高鹽、低pH及高氨氮的藥品及測量儀器分別為:粗鹽(鹽度計折射儀HT211ATC)、1 mg/L HCl和1 mg/L NaOH[25](高精度pH測試筆ATC)和NH4Cl分析純晶體(水質分析儀W-I)。預實驗每個梯度設3個重復(每組30尾蝦)于調好的實驗海水中進行,因水體中的低pH會緩慢恢復正常,故每2 h調整1次實驗海水pH并統計死亡數,并全程保持溶氧不低于6.0 mg/L,時間持續至對蝦死亡過半為止。當高鹽為35、低pH為6±0.1和高氨氮為70 mg/L時,由SPSS 19.0算出LT50%為47.31 h,最接近48 h,故將該3因子濃度作為綜合脅迫的濃度(因在第1組中便確定了綜合脅迫的濃度,故并未繼續開展第2組和第3組預實驗)。
綜合脅迫正式實驗:對蝦暫養3 d后開始實驗,由綜合脅迫預實驗確定的濃度開展正式實驗,其他脅迫條件與綜合脅迫預實驗相同,每個群體隨機取實驗用蝦90尾 (設3個重復,每組30尾)置于小筐中放于水深30 cm的池(長×寬:5 m×5 m)中進行綜合脅迫。按交配組合群體排序,生長性狀與耐綜合脅迫性狀一一對應,每2 h調一次實驗海水pH、統計死亡數、記錄存活時間并測量生長性狀(用精度為0.01位的分析天平連接電腦自動錄入體質量數據,數碼相機拍照后用ImageJ軟件測體長、頭胸甲長及腹節全長),實驗持續至全部對蝦死亡時結束。
2.3 雜種優勢及配合力估算
2.3.1 凡納濱對蝦生長及耐綜合脅迫雜種優勢
雜種優勢的公式[7, 26-29]為
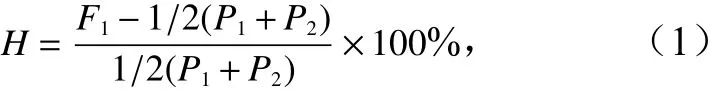
式中,F1、P1和P2分別表示親本1和親本2正反交子一代、親本1自交子一代表型均值、親本2自交子一代表型均值;H為正反交子一代雜種優勢率。
2.3.2 凡納濱對蝦生長及耐綜合脅迫性狀的配合力分析模型
模型公式為

式中,yijk為第i個母本群體與第j個父本群體雜交的第k個個體的生長及耐綜合脅迫性狀的觀測值;μ為總體均值;Dami為第i個群體母本的GCA;Sirej為第j個群體父本的GCA;Parentsij為第i個母本群體與第j個父本群體雜交的特殊配合力 (Specific Combining Ability, SCA);eijk為隨機效應殘差。
利用ASReml4[30]軟件和親本模型進行方差組分剖分及配合力估計。估計顯著性時分別將親本模型中的GCA或SCA 剔除掉,分別構建兩個模型,用ASReml4軟件中的lrt.asreml函數進行顯著性檢驗,根據一般配合力和特殊配合力的顯著性,決定是否進行下一步的LSD多重比較[31];參照文獻[32-36]方法計算GCA方差、SCA方差分別占表型方差的比例。
3 結果與分析
3.1 凡納濱對蝦親本群體的GCA
凡納濱對蝦5個不同遺傳背景群體的GCA效應值見表2。結果顯示:父本群體體質量、體長和頭胸甲 長 的GCA范 圍 分 別 為(-8.90×10-7±0.000 5)~(3.62×10-7±0.000 5)、(-9.34×10-6±0.003 3)~(5.22×10-6±0.003 3)、(-1.92×10-6±0.000 8)~ (2.03×10-6±0.000 8),母本群體體質量、體長和頭胸甲長的GCA范圍分別 為:(-1.23×10-6±0.000 5)~(5.31×10-7±0.000 5)、(-5.70×10-6±0.002 6)~(2.81×10-6±0.002 6)、(-2.71×10-6±0.000 8)~(1.24×10-6±0.000 8),均 較 接 近 于0;父母本群體耐綜合脅迫的GCA范圍為:(-0.93±1.687 87)~(1.46±1.614 2),其中父本群體的GCA范圍為(-8.55×10-6±0.005 4)~(7.74×10-6±0.005 4),均較接近于0。

表2 凡納濱對蝦生長和耐綜合脅迫性狀的GCA效應值Table 2 GCA effect value of growth and comprehensive stress tolerance traits of Litopenaeus vannamei
3.2 凡納濱對蝦12個組合的SCA
凡納濱對蝦12個組合生長和耐綜合脅迫性狀的特殊配合力效應值見表3。結果顯示:12個組合體質量、體長、頭胸甲長和腹節全長的SCA范圍分別為(-0.92±0.25)~(0.35±0.25)、(-5.82±1.35)~(2.02±1.60)、(-0.61±0.29)~(0.31±0.27)和 (-0.39±0.39)~(0.17±0.4),其中,組合K♀×K♂(0.35±0.25)、GX♀×W♂(0.31±0.22)和GX♀×M♂(0.29±0.29)體質量的SCA排名靠前;12個組合耐綜合脅迫的SCA范圍為(-2.79±2.02)~(3.65±1.79),其中組合GX♀×GX♂(3.65±1.79)、L♀×L♂(2.19±2.33)和W♀×GX♂(1.30±2.03)耐綜合脅迫的SCA排名靠前。此外,母本群體生長(4.98×10-6%~0.43%)和耐綜合脅迫(0.73%)及父本群體生長(5.00×10-6%~0.54%)和耐綜合脅迫(5.08×10-6%)的GCA方差占表型方差的比例總體來說小于生長(0.25%~4.06%)和耐綜合脅迫(1.43%)的SCA方差占表型方差的比例(表4),表明親本的SCA對子代性狀的表現起主導作用。

表3 凡納濱對蝦生長和耐綜合脅迫性狀的SCA效應值Table 3 SCA effect value of growth and comprehensive stress tolerance traits of Litopenaeus vannamei

表4 凡納濱對蝦生長和耐綜合脅迫性狀配合力的方差組分Table 4 The variance components of combining ability for growth and comprehensive stress tolerance of Litopenaeus vannamei
3.3 雜交組合生長和耐綜合脅迫性狀的雜種優勢比較
雜交組合生長和綜合脅迫耐受性雜種優勢見圖1。結果顯示:6個雜交組合GX♀×W♂、GX♀×K♂、GX♀×M♂、W♀×GX♂、K♀×GX♂和K♀×M♂的體質量、體長、頭胸甲長、腹節全長及耐綜合脅迫性狀的平均雜種優勢率(H)范圍分別為(-5.95%~25.525%)、(-1.00%~80.79%)、(-3.43%~24.90%)、(-4.14%~55.87%)和(-8.03%~42.86%),其中組合GX♀×K♂的體質量(-5.95%)、體長(-1.00%)、頭胸甲長(-3.43%)、腹節全長(-4.14%)及耐綜合脅迫(-8.03%)性狀的H均為負值,其余均為正值。其中組合GX♀×W♂體質量的H最高(25.52%),組合K♀×M♂耐綜合脅迫的H最高(42.86%),組合W♀×GX♂次之(41.72%)。

圖1 凡納濱對蝦雜交組合生長和耐綜合脅迫性狀的雜種優勢Fig. 1 Heterosis of growth and comprehensive stress tolerance in hybrid combinations of Litopenaeus vannamei
4 討論
4.1 凡納濱對蝦生長和耐綜合脅迫性狀的配合力分析
配合力是雜交育種中評估雜交效果與篩選優良雜交組合的重要遺傳參數;配合力又分一般配合(GCA)和特殊配合力(SCA),前者由基因的加性效應決定可穩定遺傳[33,37];后者由基因非加性效應(顯性、超顯性及上位效應)決定不可穩定遺傳[38],是產生雜種優勢的必要條件[21,34,39]。對凡納濱對蝦配合力的研究已有相關報道[20-22],然而有關凡納濱對蝦耐綜合脅迫配合力的研究未見報道,本研究以5個不同遺傳背景的多世代選育凡納濱對蝦群體(具有較高純度)為親本構建子代組合,并嘗試估計各組合耐綜合脅迫和生長性狀的配合力。結果顯示,父本群體體質量、體長和頭胸甲長的GCA分別為(-8.90×10-7±0.000 5)~(3.62×10-7±0.000 5)、(-9.34×10-6±0.003 3)~(5.22×10-6±0.003 3)、(-1.92×10-6±0.000 8)~(2.03×10-6±0.000 8),母本群體體質量、體長和頭胸甲長的GCA分別為(-1.23×10-6±0.000 5)~(5.31×10-7±0.000 5)、(-5.70×10-6±0.002 6)~(2.81×10-6±0.002 6)、(-2.71×10-6±0.000 8)~(1.24×10-6±0.000 8),均較接近于0,父本群體耐綜合脅迫性狀的GCA為(-8.55×10-6±0.005 4)~(7.74×10-6±0.005 4),均較接近于0,與王浩等[20]報道的凡納濱對蝦7個群體生長性狀GCA(-0.53~0.42)及胡志國等[21-22]報道的凡納濱對蝦父母本群體耐低溶氧與耐高氨氮及耐高鹽與耐低鹽的GCA(-0.215~0.152 2和-0.215 3~0.152 2)存在差異;與林明雪等[40]報道的多世代選育的高純度“吉富”系列尼羅羅非魚雜交,生長性狀的GCA為0且表現出較高的雜種優勢結果相似,推測選育親本群體的GCA可能會隨選育世代和遺傳純度的增加而逐漸趨近于0,具體原因還有待進一步的研究(包括對多代選育純系內交配及多代選育純系間雜交的配合力進行更加全面、詳細的研究和探討)。
利用LRT似然比進行配合力的顯著性檢驗,發現GCA不顯著,SCA顯著,續而對SCA進行多重比較(LSD-t檢驗)。結果顯示,組合K♀×K♂(0.35±0.25)、GX♀×W♂(0.31±0.22)和GX♀×M♂(0.29±0.29)體質量的SCA排名靠前、效應值為正且3組合的t檢驗差異不顯著(p>0.05);組合GX♀×GX♂(3.65±1.79)、L♀×L♂(2.19±2.33)和W♀×GX♂(1.30±2.03)耐綜合脅迫的SCA排名靠前、效應值為正且3組合的t檢驗差異不顯著(p>0.05),說明前3個組合存在較高的體質量非加性效應,后3個組合存在較高的耐綜合脅迫非加性效應。可考慮將組合K♀×K♂、GX♀×W♂和GX♀×M♂作為子二代快長的備選組合,將組合GX♀×GX♂、L♀×L♂和W♀×GX♂作為子二代耐綜合脅迫的備選組合。該研究可為凡納濱對蝦配套系雜交提供參考資料,隨著研究進一步的深入, 筆者后續將結合配合力評估凡納濱對蝦生長和耐受性能的三系、四系配套效果。
4.2 凡納濱對蝦生長和耐綜合脅迫性狀的雜種優勢分析
雜種優勢是兩個不同群體的親本雜交子代在繁殖、存活和生長等生活力方面優于親本的現象[34],在生產上,被用來評估不同雜交方案的可行性。中國明對蝦(Fenneropenaeus chinensis)雜交、凡納濱對蝦(L.vannamei)雜交,在生長性狀上表現出雜種優勢[29,41-42]。在本研究中,雜交群體GX♀×W♂、GX♀×M♂、W♀×GX♂、K♀×GX♂和K♀×M♂的體質量、體長、頭胸甲長、腹節全長及耐綜合脅迫的雜種優勢均為正(0.10%~80.79%和6.87%~42.86%),說明雜交群體在生長及耐綜合脅迫性狀上表現出一定的雜種優勢。Lu等[7]認為,大量的雜種優勢可能是由于有利顯性等位基因的積累或雜種中顯性等位基因掩蓋隱性有害等位基因而產生的;從近交使有害基因暴露的角度考慮,雜種優勢僅是近交衰退的補償,也就是說雜交子代不表現優勢而可能表現劣勢[23,43],田燚等[43]報道中國對蝦兩群體雜交,F1代早期群體YP♂×KN♀存活率表現出一定雜種優勢,而群體YP♀×KN♂表現出雜種劣勢;袁瑞鵬等[23]報道凡納濱對蝦群體雜交F1代群體YH♀×ZX♂低溶氧耐受性具有雜種優勢,而群體ZK♀×YH♂低溶氧耐受性表現雜種劣勢;在其他水產動物中,雜交子代也表現出雜種優勢與雜種劣勢[44]。在本研究中,雜交群體GX♀×K♂的體質量(-5.95%)、體長(-1.00%)、頭胸甲長(-3.43%)、腹節全長(-4.14%)及耐綜合脅迫(-8.03%)性狀的H為負值,表現出一定的雜種劣勢。總的來說,雜交中表現出的雜種優勢表明,雜交對提高凡納濱對蝦生長和耐綜合脅迫性狀具有正向效應。此外,本研究篩選出組合GX♀×W♂體質量的H最高,為25.52%;組合GX♀×M♂體長、頭胸甲長和腹節全長的H均最高,分別為80.79%、24.90%和55.87%;組合K♀×M♂耐綜合脅迫的H最高,為42.86%,組合W♀×GX♂的次之,為41.72%,可考慮將組合GX♀×W♂和組合GX♀×M♂作為快長配套雜交的備選組合,將組合K♀×M♂和組合W♀×GX♂作為耐綜合脅迫配套雜交的備選組合。
5 結論
本研究利用親本模型分析了“興海1號”新品種核心群體(GX)、泰國引進群體(W和L)和美國引進群體(K和M)5個凡納濱對蝦種質群體之間的GCA和SCA,并在此基礎上分析了5個群體之間的雜種優勢。結果顯示,雜交后代生長和耐綜合脅迫性狀的表現主要受到親本SCA的影響,雜交組合生長(體質量、體長、頭胸甲長與腹節全長)和耐綜合脅迫性狀均表現出一定的雜種優勢;雜交組合GX♀×W♂體質量的GCA和H最高;雜交組合W♀×GX♂耐綜合脅迫的SCA最高且H較高,可考慮分別加強該兩組合在生長和耐綜合脅迫性狀方面的配套雜交應用。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
