泡核桃凋落葉對生菜光合特性的影響
2022-04-25 05:25:42鄭崇蘭李志超祝銘謙巫玲琳劉永安
中國果菜 2022年4期
鄭崇蘭,龍 麗,黃 慶,李 培,李志超,祝銘謙,巫玲琳,劉永安*
(1.涼山彝族自治州林業(yè)草原科學(xué)研究院,四川西昌 615000;2.涼山彝族自治州張林國有林場,四川西昌 615000)
植物化感作用是指植物通過向外界環(huán)境中釋放有效的化合物,對自身和相鄰的其他植物(包括微生物)產(chǎn)生的促進或抑制作用[1]。泡核桃(Juglans sigillataDode)廣泛分布于我國西南地區(qū),是分布區(qū)重要的果用、油料、材用和綠化的經(jīng)濟林樹種,被許多地區(qū)列入經(jīng)濟建設(shè)的主導(dǎo)產(chǎn)業(yè)進行重點開發(fā)。開展泡核桃林下種植可充分利用土地資源,但已有研究表明核桃葉片中含有大量的醌類、黃酮類、二芳基庚烷類、萜類、多酚類等化感物質(zhì)[2-3],林下凋落葉勢必對林地環(huán)境及林下植物產(chǎn)生影響。研究發(fā)現(xiàn),核桃凋落葉在土壤中分解,顯著抑制蘿卜、白菜、芥菜葉片中的葉綠素合成、降低三種作物的光合能力[4],且抑制作用隨著凋落葉添加量的增加而增強[5]。而將核桃凋落葉化感作用耐受性強的植物用于開展林下套作,能有效提高土地利用率[5]。有研究表明生菜對麻風(fēng)樹葉子、種子殼的水浸提液[6]及橡膠葉水浸提液[7]的化感作用較不敏感,受銀杏凋落葉[8]和銀杏外種皮[9]的化感抑制作用卻很顯著。
本研究以生菜(Spinaciao leracea)為受體植物,采用盆栽試驗,向土壤中添加不同量的泡核桃凋落葉,模擬泡核桃凋落葉自然分解,探討泡核桃凋落葉對生菜光合作用的影響,為泡核桃林下種植提供理論依據(jù)。
1 材料與方法
1.1 試驗材料
采用四川省涼山州木里縣15 年生泡核桃人工林中采集的新鮮凋落葉,粉碎后,混合均勻,備用。
選擇大小均勻、飽滿的生菜種子,播種前用0.5%K2MnO4溶液消毒20 min,用蒸溜水沖洗5 次,備用。
栽種土壤選擇芬蘭進口泥炭土,基本理化性質(zhì)見表1。栽種器具選用上口徑24 cm、下口徑17 cm、高14 cm的聚乙烯塑料盆。
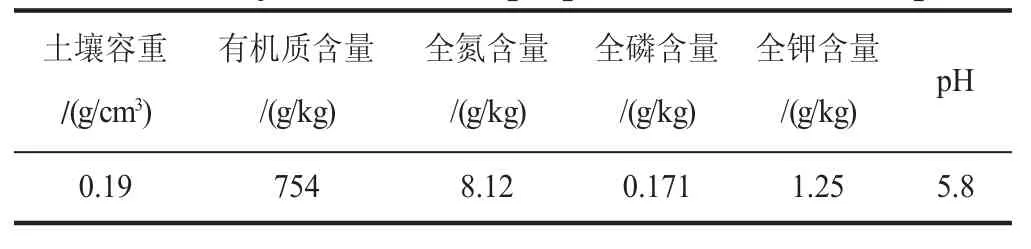
表1 供試土壤基本理化性質(zhì)Table 1 Physico-chemical properties of the soil samples
1.2 試驗設(shè)計
本試驗于2021 年5—9 月在涼山州林業(yè)草原科學(xué)研究院塑料大棚內(nèi)進行。試驗采取單因素隨機試驗設(shè)計,依據(jù)調(diào)查數(shù)據(jù),人工泡核桃林每年平均凋落葉量約為8 000 kg/hm2,按盆口徑22 cm 折合量為30 g/盆,故本試驗設(shè)置凋落葉添加量分別為T15(15 g/ 盆,約4 000 kg/hm2)、T30(30 g/ 盆,約8 000 kg/hm2)以 及T60(60 g/盆,約16 000 kg/hm2)3 個處理組,以不添加凋落葉為對照組。每個組設(shè)置10 個重復(fù),共計40 盆。
按處理要求,將定量的凋落葉與試驗用土充分混勻后澆透水,1 d 后播種生菜。播種時覆蓋0.5 cm 試驗用土,澆透水以便種子發(fā)芽。各處理全部出苗后10 d 進行間苗,然后實施常規(guī)田間管理。
1.3 指標(biāo)測定及方法
葉片光合生理指標(biāo)采用便攜式光合作用儀(ADClci-t,英國)測定,光合儀選擇開放氣路,人工設(shè)置光照強度為1 000 μmol/(m2·s),選取莖中部完全成熟的葉片測定葉片凈光合速率(net photosynthetic rate,Pn)、蒸騰速 率(transpiration rate,Tr)、氣孔導(dǎo)度(stomatal conductance,Gs)和胞間CO2濃度(intercellular CO2concentration,Ci),每個處理重復(fù)測定3 次,每次重復(fù)記錄5 個數(shù)據(jù)。日進程于8:00—18:00 進行測定,光合生理特征參數(shù)在上午9:00—11:00 進行測定。
1.4 計算方法
化感作用效應(yīng)敏感指數(shù)簡稱化感指數(shù)(RI),計算公式如式(1)(2)所示。

式中,C為對照CK 光合參數(shù)測定值,T為處理T15、T30、T60 光合參數(shù)測定值。
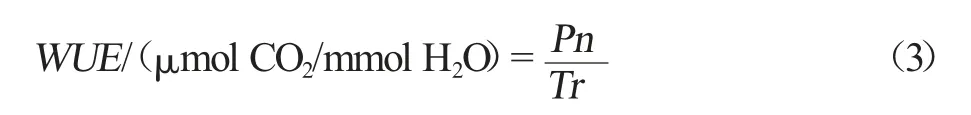
葉片水分利用效率(WUE)計算公式如式(3)所示。式中,Pn 為葉片凈光合速率,μmol CO2/(m2·s);Tr為葉片蒸騰速率,mmolH2O/(m2·s)。
1.5 數(shù)據(jù)處理
采用Excel 2007 進行數(shù)據(jù)整理,SPSS 19.0 進行單因素方差分析,LSD 法進行多重比較。
2 結(jié)果與分析
2.1 泡核桃凋落葉對生菜光合特征參數(shù)的影響
由表2 可以看出,T15 處理的生菜凈光合速率最大,然后依次為CK、T30,最小為T60,泡核桃凋落葉對生菜凈光合速率的影響總體表現(xiàn)為低量促進、高量抑制。低量處理T15 的凈光合速率為13.08 μmol CO2/(m2·s),顯著高于CK,且其化感指數(shù)為0.081,表現(xiàn)為促進效應(yīng)。高量處理T30、T60 的凈光合速率分別為6.32 μmol CO2/(m2·s)和4.65 μmol CO2/(m2·s),化感指數(shù)為-0.474 和-0.613,表現(xiàn)為抑制效應(yīng),且各處理間凈光合速率測定值的差異顯著(P<0.05)。由此可見,每盆土添加15 g 泡核桃凋落葉,可顯著促進生菜的凈光合速率,添加量超過30 g 之后,變?yōu)橐种谱饔谩?/p>
由表2 可知,胞間CO2濃度表現(xiàn)為隨泡核桃葉添加量的增加逐漸增加,CK 最低為271.59 mmol/mol,T15 較CK 稍高,為272.69 mmol/mol,但與CK 無顯著差異,化感指數(shù)僅為0.004。處理T30 與T60 的胞間CO2濃度顯著高于CK的,分別為 308.37 mmol/mol 與 332.16 mmol/mol。化感指數(shù)達到了0.119 和0.182,表現(xiàn)為促進作用。這表明,每盆土添加量15 g 泡核桃凋落葉對生菜胞間CO2濃度影響不大,添加量達30 g 之后,胞間CO2濃度表現(xiàn)出上升趨勢。
氣孔導(dǎo)度變化與凈光合速率類似,表現(xiàn)出低促高抑的趨勢,T15 氣孔導(dǎo)度為0.21 mol/(m2·s),顯著高于對照CK,化感指數(shù)為0.098,表現(xiàn)為促進作用。當(dāng)每盆泡核桃凋落葉添加量達到30 g 時,其對生菜葉片氣孔導(dǎo)度的影響變?yōu)橐种疲琓30、T60 氣孔導(dǎo)度下降為0.11 mol/(m2·s)和0.12 mol/(m2·s),化感指數(shù)為-0.401 和-0.350,T30 和T60較CK 相比顯著下降,但處理間無較大差異。
泡核桃凋落葉對生菜片蒸騰速率的影響結(jié)果見表2。由表知,低量處理T15 的蒸騰速率為4.90 mmol/(m2·s),顯著高于對照,化感指數(shù)為0.147,表現(xiàn)為促進作用。高量處理(T30、T60)降低了生菜葉片蒸騰速率,化感指數(shù)為-0.097 和-0.033,表現(xiàn)為抑制效應(yīng)。
由表2 可以看出,土壤中添加泡核桃凋落葉降低了生菜葉片的水分利用效率,且降低程度隨添加量的增加而增加,且各處理間差異顯著(P<0.05),化感指數(shù)達到了-0.072、-0.417 和-0.595。

表2 不同泡核桃葉添加量處理下生菜的凈光合速率及氣體交換參數(shù)Table 2 Net photosynthetic rate and gas exchange paramaters of lettuce under different amount of J. sigillate leaf litter
2.2 泡核桃凋落葉對生菜光合特征參數(shù)日變化的影響
由圖1A(見下頁)可看出,CK、T15、T30 和T60 四種處理凈光合速率日變化均呈現(xiàn)“雙峰型”,在10:00 時和14:00 分別達到峰值,有明顯的“光合午休”現(xiàn)象。4 個處理均在10:00 時達到全天的最高峰,T15 最大,為11.63 μmol/(m2·s),之后依次為CK、T30 和T60,且各處理間有顯著差異(P<0.05)。在14:00 時達到第二次高峰,各處理均低于第一次高峰,其峰值以CK 為最高,凈光合速率達到8.01 μmol/(m2·s),與T15(凈光合速率為7.99 μmol/(m2·s)無顯著差異,均顯著高于T30 和T60。
由圖1B(見下頁)可知,CK、T30 和T60 處理下生菜苗氣孔導(dǎo)度均呈現(xiàn)“雙峰型”,在10:00 出現(xiàn)第一個峰值,14:00 出現(xiàn)第二個峰值。CK 處理的最大峰值出現(xiàn)在下午16:00,達到0.188 mol/(m2·s),而T30 和T60 峰值出現(xiàn)在12:00,分別為0.128 mol/(m2·s)和0.154 mol/(m2·s)。T15 的氣孔導(dǎo)度在14:00 以前未出現(xiàn)明顯的峰值,最大值為0.148 mol/(m2·s),但各時間(除CK 16:00 外)均顯著高于其它處理。
由圖1C 可知,四個處理的胞間CO2濃度均呈現(xiàn)“W型”,在早上8:00 的胞間CO2濃度較高,然后下降,在中午12:00 出現(xiàn)第二個峰值,隨后下降,在下午14:00 出現(xiàn)第二個低谷,隨后逐漸上升,在下午18:00 達到全天的峰值。胞間CO2濃度大小依次為T60>T30>CK>T15。
由圖1D 可看出,不同泡核桃葉添加量處理的生菜蒸騰速率日變化均呈現(xiàn)為“單峰型”,各處理均從8:00 開始上升,在中午12:00 達到峰值,隨后逐漸下降。T15、CK、T60 和T30 各處理的峰值大小分別達到8.809、7.211、6.450、5.971 mmol/(m2·s)。
由圖1E 可知,不同泡核桃凋落葉處理生菜的水分利用效率的日變化,總體呈現(xiàn)下降趨勢,在中午12:00 時出現(xiàn)一個低谷,在下午14:00 時出現(xiàn)一個小高峰,其峰值遠低于早上8:00,然后開始下降。各處理的全天平均水分利用效率由大到小依次為CK、T15、T30 和T60,分別為1.095、1.075、0.909、0.655 μmol CO2/mmol H2O。
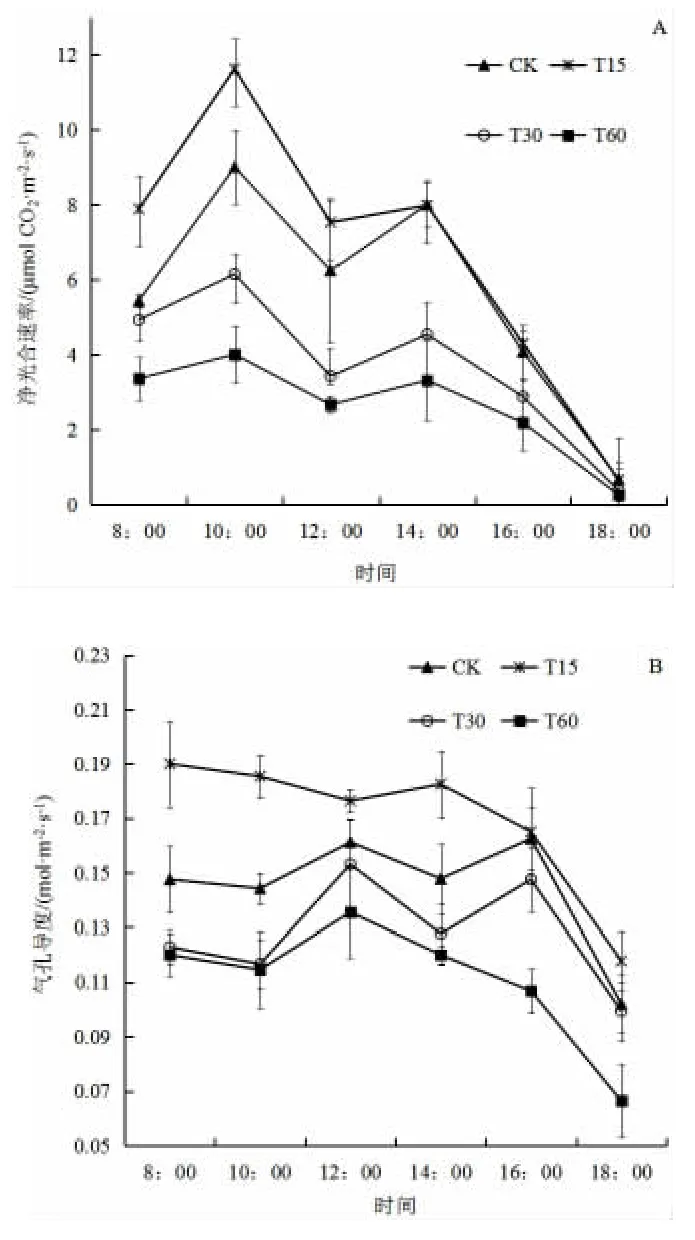
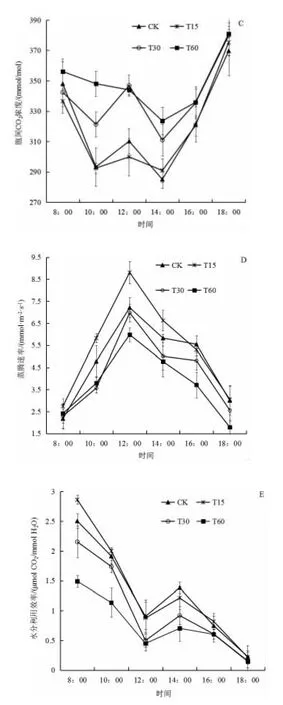
圖1 不同泡核桃凋落葉添加量處理下生菜葉片氣體交換參數(shù)日變化Fig.1 Diurnal variation characteristics of gas exchange parameters of lettuce under different amount of J. sigillate leaf litter
3 討論與結(jié)論
本試驗中,生菜的凈光合速率隨凋落葉添加量增加先升高后降低,胞間CO2濃度隨凋落葉添加量的增加而增加,且均高于對照,而氣孔導(dǎo)度則表現(xiàn)為先升高后降低。植物葉片光合效率降低有氣孔部分關(guān)閉導(dǎo)致的氣孔限制和葉肉細胞光合活性下降導(dǎo)致的非氣孔限制兩類[10]。Farquhar 等[11]認為,如果凈光合速率的降低伴隨著胞間CO2濃度的降低,則凈光合速率的下降主要是受部分氣孔關(guān)閉的氣孔限制所致;反之,如果凈光合速率下降伴隨著胞間CO2濃度升高或不變,則凈光合速率的下降主要是由葉肉細胞光合活性下降導(dǎo)致的非氣孔限制引起的。初步可以推斷,本試驗中生菜凈光合速率的下降是非氣孔限制導(dǎo)所致。低量添加泡核桃凋落葉提高了生菜的凈光合速率,而土壤中泡核桃葉凋落葉增多時其凈光合速率降低,且顯著低于CK,說明添加一定量的泡核桃凋落葉能提高生菜的光合效率,可能是因為低量的凋落葉分解產(chǎn)生的化感物質(zhì)較少,不足以對生菜生長產(chǎn)生影響,反而通過凋落葉分解產(chǎn)生的營養(yǎng)物質(zhì)促進了生菜的生長;超過一定量后,過多凋落葉分解產(chǎn)生的化感物質(zhì)積累,破壞了生菜葉片光合器官的結(jié)構(gòu),從而導(dǎo)致凈光合速率顯著下降。
另外,與對照相比,不同處理生菜水分利用效率均顯著下降,T15 可能是因為化感物質(zhì)使生菜葉片的氣孔導(dǎo)度增加,從而提高了蒸騰速率;而高添加量處理T30、T60 生菜的水分利用效率下降,可能是因為高濃度的化感物質(zhì)抑制了生菜的光合作用。光合作用是植物能量的來源,也是植物生長、發(fā)育及代謝的物質(zhì)基礎(chǔ)。環(huán)境因子的改變會影響植物的光合特性[12],化感物質(zhì)對植物體光合作用的影響主要表現(xiàn)在使葉綠素含量和光合速率的降低上[13-15]。曾有研究表明,核桃凋落葉的化感物質(zhì)能顯著地降低小麥、燕麥、黑麥草和萵筍等植物的葉綠素含量[16-17]。化感物質(zhì)可以通過影響氣孔導(dǎo)度、胞間CO2濃度、蒸騰速率等氣孔因素直接影響植物的光合作用[18]。
本試驗中,低濃度泡核桃凋落葉處理的生菜凈光合速率在上午12:00 以前顯著高于對照,在下午16:00 其凈光合速率從“午休”后恢復(fù)較慢,其凈光合速率和氣孔導(dǎo)度提高較小,胞間CO2濃度降低較小,由此暗示經(jīng)泡核桃凋落葉處理后,因泡核桃葉中的化感物質(zhì)影響了氣孔開張,從而影響了生菜的光合效率和水分利用效率。馬紅葉等[19]研究也表明化感物質(zhì)可通過降低葉片中的葉綠素含量、影響蒸騰速率和氣孔導(dǎo)度等方式來降低植物的光合速率。
綜上,泡核桃凋落葉中的化感物質(zhì)對生菜植物的光合生理產(chǎn)生顯著的影響,盆土中泡核桃凋落葉添加量為15 g,即約4 000 kg/hm2時,能促進生菜光合作用;盆土中泡核桃凋落葉添加量達到30 g,即約8 000 kg/hm2時,能提高胞間CO2濃度,降低氣孔導(dǎo)度和蒸騰速率和水分利用效率,從而降低植物吸收、轉(zhuǎn)化和利用光能的能力,導(dǎo)致光合速率的降低,影響植物對碳的固定和同化,從而抑制植物的光合效率。試驗結(jié)果表明,在去除部分凋落物之后,可在泡核桃林下套種生菜,以提高土地利用率,但具體栽培方式需進行大田試驗進一步確定。
