土壤微生物多樣性通過共現網絡復雜性表征高寒草甸生態系統多功能性
2022-04-25 02:12:04張君紅王健宇孟澤昕董政宏劉凱茜陳文青
生態學報 2022年7期
張君紅,王健宇,孟澤昕,何 佳,董政宏,劉凱茜,陳文青, *
1 西北農林科技大學草業與草原學院,楊凌 712100
2 西安交通大學人居環境與建筑工程學院,西安 710049
3 中華環境保護基金會,北京 100062
生物多樣性與生態系統功能(BEF)的關系是當前生態學研究的熱點問題[1]。土壤微生物群落是陸地生態系統的重要組成部分,在養分循環、有機物分解和促進植物生長等生態功能方面發揮著重要作用[2—5]。由于土壤微生物不僅具有高度的物種多樣性,而且物種間存在復雜的相互作用關系,因此,揭示土壤微生物多樣性與生態功能的關系除了關注其物種數量效應外還需進一步考慮微生物間復雜的相互作用關系[5—7]。早期對BEF的研究多側重于單一生態功能與生物多樣性的關系[8—10]。隨著研究的深入,人們逐漸認識到較高的生物多樣性能顯著提高生態系統對多種生態系統功能的支持能力,并將生態系統能夠同時維持多種生態系統功能和服務的能力定義為生態系統多功能性(EMF)[9]。相較于單一生態功能的研究,EMF可使生物多樣性的多功能冗余減小,并且使多樣性的重要性得以體現[11—12],因此EMF能更好地反映BEF關系。
土壤微生物多樣性與生態系統功能的關系仍然存在爭論[13—16]。雖然相關的原位調查研究[13, 17]和控制實驗[14, 18]的研究結果均表明微生物多樣性與生態功能存在顯著的正相關關系,但也有研究顯示兩者間存在負相關[15]和無顯著關聯關系[16]。上述研究均基于微生物多樣性中的物種數目與EMF的關系展開,忽視了復雜微生物群落間的相互作用關系在生態系統功能維持中的作用。在自然環境中,微生物通過協同、競爭及拮抗等一系列直接和間接的生態作用相互關聯形成微生物網絡[7, 19]。該網絡是生態系統實現能量流動、物質循環和信息傳遞基本功能的基礎[19—20]。此外,微生物間的相互作用關系受時間和空間尺度的影響,不同尺度上同一共存物種間的關系存在差異[21],相同的共存物種并不代表著相同的種間關系。以往的研究多單獨考慮微生物物種多樣性對生態系統功能的影響,缺少綜合考慮物種多樣性和微生物互作網絡對EMF影響的研究,因此不能全面反映BEF關系[2, 13, 19]。
微生物共現網絡可揭示微生物間復雜的相互作用關系,能反映物種多樣性不能反映的生態聯系和生態過程[7, 22—23]。當前研究多關注不同生境微生物共現網絡的模式或結構特征,微生物網絡結構與生態功能間的關系仍缺乏進一步研究[24—27]。Wagg等[19]通過室內微生物多樣性控制試驗,利用邊密度作為微生物網絡復雜性指標來表征微生物間復雜的相互作用關系,證實土壤微生物網絡復雜性在維持生態系統功能方面發揮著積極作用。這項研究通過不同孔徑篩網對天然草地生態系統采集到的土壤過篩,根據微生物個體尺寸差異構建了不同群落復雜性的微生物群落,盡管可在一定程度反映群落復雜性對生態功能的影響,但該方法劃分出的群落并不能代表自然環境中的微生物群落,微生物在自然環境中并非根據人為劃分的個體尺寸聚集成群落,而是由一系列生物和非生物因子長期共同作用形成群落[19, 28]。因此,本研究采用原位調查試驗分析原位生境中微生物網絡復雜性與EMF的聯系以期深化對微生物BEF關系的認識。
青藏高原是世界海拔最高和面積最大的高原,在涵養水源、調節氣候以及土壤有機碳固持等生態功能方面發揮著重要作用[29—31]。青藏高原高寒生態系統對氣候變化十分敏感,氣候變化會導致微生物多樣性喪失,進而對EMF產生不利影響[1]。高寒草甸作為該系統具有代表性的草地生態系統類型,研究該區域土壤微生物多樣性和網絡復雜性與EMF的關系對全球變化背景下高寒草地生態系統生態功能的維持具有重要意義[1, 32—33]。高寒生態系統中的氣候和土壤特性在不同海拔高度上差異較大,環境因子會影響微生物群落組成以及微生物間的相互作用[27, 34—35]。因此,本研究于青藏高原高寒草甸選取了6個海拔高度(3755 m、3994 m、4225 m、4534 m、4900 m、5120 m),分析了微生物多樣性和網絡復雜性與EMF的聯系,進一步揭示該區域微生物多樣性與EMF的關系,以期為全球變化背景下高寒草地生態系統生態功能的微生物調控機制提供更全面的見解。
1 材料與方法
1.1 研究區概況
研究區位于西藏自治區米拉山西坡(29°49′59"—29°49′42"N, 91°37′55"—92°20′22"E),海拔3700—5300 m,區域年均溫-0.8—5.3℃,年降水量250—640 mm,雨熱同期,多集中于生長季(5—9月)。土壤類型以高山草原土、高山草甸土和高山寒凍土為主[36—37]。植被類型以高寒草甸為主,主要有三刺草(Aristidatriseta)、固沙草(Orinusthoroldii)、喜馬拉雅草沙蠶(Tripogonhookerianus)、白草(Pennisetumflaccidum)、絲穎針茅(Stipacapillacea)、長芒草(Stipabungeana)等[37—38]。沿海拔高度(3755 m、3994 m、4225 m、4534 m、4900 m和5120 m)選取坡度、坡向一致且無明顯的家畜和人為活動干擾,具有代表性的植被和土壤特性的6個50 m×50 m樣地。
1.2 樣品采集
于2019年8月(該地區植物生物量最大時期),在6個樣地內以五層嵌套的四邊形取樣方式設置20個1 m2(1 m×1 m)樣方(圖1),樣方分別位于四邊形的四個角。以單位面積內物種的數量作為植物多樣性(Plant richness, PSR)指標。齊地刈割收集地上植物部分,置于105℃烘箱0.5 h殺青后,60℃烘至恒重,稱重即得地上生物量。本研究使用植物生物量表征地上凈初級生產力(Aboveground net primary production, ANPP)[28]。用土鉆采集表層土壤樣品(直徑5 cm,深度20 cm),每個樣方內采集6—7個子樣,去除雜質混合均勻后過2 mm土篩,將所得土壤樣品分兩部分:一部分帶回室內冰箱置于4℃儲存,用于測定土壤理化性狀和部分生態功能指標;另一部分儲存于-80℃冰箱,用于微生物分析。

圖1 取樣示意圖
1.3 氣候和土壤因子測定
將土壤與水1:2.5混合成懸濁液,使用pH計(UV- 1800, Shimadzu, Japan)測定土壤pH值。采用烘干法測定土壤含水量(Soil moisture, SM),具體操作為稱取5 g土壤在105℃烘12 h。取1 g風干土壤,加入20 mL的0.11 mol/L乙酸,先于離心機中以30 rpm離心20 h,后3000 rpm離心20 min,得到土壤可交換陽離子(Na+、K+、Ca2+、Mg2+、Al3+)提取物,隨后使用ICP-OES(Spectro Analytical UK, Halesowen, UK)測定其含量[39]。參照Kettler等人[40]的方法測定土壤質地,即砂粒、粉粒和黏粒含量(Sand%, Silt%, Clay%)。具體操作步驟如下:取15 g風干土壤,加入3%的六偏磷酸鈉溶液45 mL,于往復式振動篩120 rpm振動2 h,后將土壤懸浮液過0.053 mm篩收集砂粒。將過篩后的土壤溶液充分攪拌,室溫下靜置2 h,上部懸浮液部分即為黏粒,沉淀部分為粉粒。將收集到的砂粒和粉粒于105℃下烘干至恒重,計算其占原始樣品質量的百分比,即為砂粒和粉粒含量,100%減去砂粒和粉粒含量得黏粒含量。通過CHELSA(http://chelsa-climate.org/)在30弧秒(約1 km)分辨率下得到6個海拔的年均溫(Mean annual temperature, MAT)和年降水量(Mean annual precipitation, MAP)[41—42]。
1.4 DNA提取、高通量測序及生物信息學分析
每個土樣各稱取3份0.25 g土壤,按PowerSoil DNA提取試劑盒(MO BIO laboratories, Carlsbad, CA, USA)說明書提取微生物DNA,然后將其合并以減少誤差。在1%瓊脂糖凝膠上分離和識別DNA提取物,并使用NanoDrop UV-Vis分光光度計(ND- 2000c, NanoDrop Technologies, DE, USA)測定提取DNA的濃度和純度。使用515F(5′-GTGCCAGCMGCCGCGGTAA- 3′)和806R(5′-GGACTACHVGGGTWTCTAAT- 3′)擴增細菌16S rRNA基因[43],ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA- 3′)和ITS2- 2043R(5′-GCTGCGTTCTT CATCGATGC- 3′)擴增真菌ITS基因[44],并通過上海美吉生物醫藥有限公司在Illumina MiSeq測序平臺對擴增產物進行高通量測序。
按Caporaso等人[45]的方法使用QIIME 1.9.1對序列進行優化和數據質控,具體步驟如下:(i)在50 bp的滑動窗口內,去除序列平均質量小于20,長度小于50 bp和模糊堿基數;(ii)對長度大于10 bp的重復序列進行組裝,無法組裝則去除;(iii)根據barcode和引物區分樣品,并調整序列方向。隨后使用UPARSE在97%的相似性閾值下劃分分類操作單元(Operational taxonomic units, OTUs)和去嵌合體[46]。基于SILVA數據庫(SILVA 132)和UNITE真菌ITS數據庫(7.2版),利用RDP分類器對75%相似閾值的OTU進行分類學分析[47—48]。細菌和真菌群落的OTUs分別作為細菌(Bacterial richness, BR)、真菌多樣性(Fungal richness, FR)指標。
1.5 生態系統功能測定和生態系統多功能性指數計算
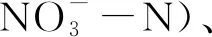
使用平均值法計算EMF指數。首先將上述10種生態功能參數通過(Xraw-Xmin)/(Xmax-Xmin)標準化,再計算平均值即得EMF指數;其中,Xraw代表每個樣方的生態功能值,Xmax代表所有樣方的最大生態功能值,Xmin代表所有樣方的最小生態功能值[5, 52]。
1.6 微生物共現網絡構建與分析
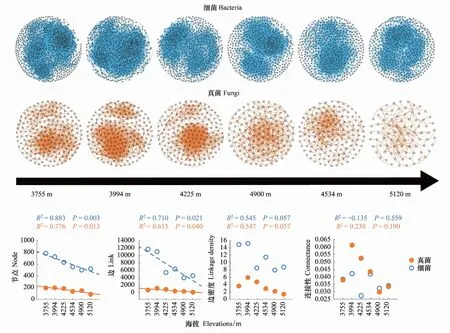
由于微生物共現網絡模式受擴散限制和環境過濾等某些生態過程影響[55],因此本研究對每個海拔高度20個樣方的土壤微生物群落構建綜合網絡,可最大程度減少上述生態過程對微生物網絡的影響。在構建網絡之前,移除豐度小于0.015%和單個樣地物種出現頻率小于5的OTUs。通過計算Spearman相關系數和Jaccard距離,基于隨機矩陣理論(Random matrix theory, RMT)方法,確定構建微生物共現網絡的閾值[26]。通過Permutation和Bootstrap,迭代1000次獲得Spearman相關系數和Jaccard距離的P值。然后使用Brown′s方法合并上述P值[56]。根據Benjamini-Hochberg等人[57]的方法控制False discovery rate(FDR)對P值進行校正,使用FDR校正后具有統計顯著性(P<0.05)的關系構建綜合網絡。使用R語言igraph包和Gephi對網絡進行可視化。隨后我們使用R語言igraph包的subgraph函數從上述所構建的綜合網絡中提取每個樣方(每個海拔各20個)的子網絡(Sub-network),每個子網絡由每個樣方所包含的OTUs以及OTUs間的關系構成[19, 58]。由于表征網絡的復雜性的節點(Node)、邊數(Link)、連接性(Connectance)和邊密度(Linkage density)存在極顯著相關性,因此本研究使用邊密度作為真菌(Fungal network complexity, FNC)和細菌網絡復雜性(Bacterial network complexity, BNC)指數[19, 25—26, 59],以探究其與EMF的關系。節點為OTU,邊為OTUs間的聯系,連接性表示網絡中節點間實際發生的相互作用數之和(邊之和)占總的潛在相互作用數(邊數)的比例,邊密度由邊數除以節點數得到[19, 25—26, 59]。
1.7 統計分析
使用單因素方差分析(One-way analysis of variance)和Duncan多重比較檢驗比較不同海拔高度間環境因子(SM、pH、Na+、K+、Ca2+、Mg2+、Al3+、Clay、Silt、Sand)的變化。采用主成分分析(Principal component analysis, PCA)分別提取土壤Na+、K+、Ca2+、Mg2+和H+、Al3+含量的第一主成分作為代表鹽基離子(Soil base mineral cations, SBC)和酸性離子(Soil acid cations, SA)的綜合指標,并引入后續的分析,通過提取主成分可減少上述離子間的共線性[60]。使用普通最小二乘法回歸(Ordinary least squares linear regressions)分別分析微生物屬性(BR、FR、BNC、FNC)與EMF,海拔與微生物屬性(BR、FR、BNC、FNC)的關系。使用Pearson相關分析分別篩選出與微生物屬性(BR、FR、BNC、FNC)和EMF均顯著相關的非生物因子,通過偏線性回歸(Partial least squares regression, PLSR)排除上述非生物因子影響,分析微生物屬性(BR、FR、BNC、FNC)和EMF指數的關系。將非生物因子對EMF的影響納入方差分解分析(Variance partition analysis, VPA),評估氣候(MAT、MAP)、土壤因子(SM、SBC、SA、Clay、Silt、Sand)、微生物多樣性(BR、FR)和微生物網絡復雜性(BNC、FNC)對EMF的影響。通過構建結構方程模型(Structural equation model, SEM)進一步分析環境因子(MAT、SM、SBC、SA、Clay)、微生物多樣性(BR、FR)、微生物網絡復雜性(BNC、FNC)與EMF的直接和間接關系。在進行SEM構建之前,基于先驗知識預先設定系統內因子間的關系(表1,圖2)。SEM分析的數據均使用R語言的scale函數進行標準化,且通過Pearson相關分析保證所有變量間均存在顯著的線性關系(P<0.05),并且不存在多重共線性(r<0.8)。模型擬合度符合以下條件:0.97≤CFI≤1.00;0.95≤GFI≤1.00;0.95≤NFI≤1.00;0≤χ2/df ≤2且0.05 圖2 氣候、土壤和微生物屬性預測生態系統多功能性的結構方程先驗模型 在青藏高原高寒草甸地帶,不同海拔高度間年均溫變幅為-0.8—5.3℃,年均降水量變幅為250—640 mm。土壤K+、Al3+、粉粒和砂粒含量沿海拔高度呈顯著升高的趨勢(P<0.05),但土壤pH、Ca2+、Mg2+、Na+和黏粒含量沿海拔高度呈顯著降低的趨勢(P<0.05)(表2)。 表2 氣候和土壤因子沿海拔高度的變化 與真菌沿海拔高度變化的共現網絡相比,細菌形成了更復雜的共現網絡(圖3)。細菌和真菌的節點和邊沿海拔高度顯著減少(P<0.05),而細菌和真菌的連接性、邊密度在海拔高度上無顯著變化(圖3)。隨著海拔高度的升高,細菌和真菌的多樣性和網絡復雜性均呈現顯著下降(P<0.05)的趨勢,其中細菌多樣性和網絡復雜性的下降趨勢均比真菌明顯,細菌和真菌網絡復雜性的下降趨勢均比多樣性明顯(圖4)。 圖3 不同海拔高度微生物的共現網絡和網絡拓撲結構屬性 圖4 微生物多樣性和網絡復雜性與海拔高度的關系 Pearson相關分析顯示,MAP、MAT、SM、Clay、SBC和SA均與BR、BNC、FNC和EMF顯著相關(P<0.05),其中MAP、SM與其顯著負相關(P<0.05),MAT、Clay、SBC、SA與其顯著正相關(P<0.05)。此外,Sand與FNC、EMF顯著正相關(P<0.05),Clay、SBC與FR、EMF顯著正相關(P<0.05)(圖5)。 圖5 生態系統多功能性指數與環境因子和微生物參數的相關性分析 細菌和真菌的網絡復雜性和多樣性與生態功能多樣性指數均顯著正相關(P<0.05),其中細菌和真菌的網絡復雜性對EMF的解釋度高于多樣性(圖6)。排除環境因子的影響后,細菌和真菌復雜性與多功能性指數的顯著負相關(P<0.001)關系仍然存在,但R2分別從0.558下降至0.185,0.482下降至0.200;而細菌和真菌多樣性與生態系統多功能性指數的顯著正相關關系(P<0.05)在排除環境因子影響后呈現出不相關關系(圖6,圖7)。 圖6 土壤細菌、真菌網絡多樣性及復雜性與生態系統多功能性的關系 圖7 土壤細菌、真菌網絡多樣性及復雜性與生態系統多功能性的偏回歸分析 VPA分析顯示微生物多樣性、氣候、土壤因子單獨作用及其交互作用對EMF的總貢獻率達73%,而微生物網絡復雜性、氣候、土壤因子及其交互作用對EMF的總貢獻率可達80%。其中微生物多樣性、氣候因子和土壤因子分別單獨解釋了1%、17%和7%的EMF差異,氣候和土壤因子的交互作用解釋了28%,土壤因子和微生物多樣性共同解釋了1%,三者共同解釋了21%。而微生物網絡復雜性、氣候因子和土壤因子單獨對EMF的貢獻率分別為8%、3%和7%,微生物網絡復雜性單獨貢獻率高于多樣性;氣候和土壤因子共同解釋了8%的EMF差異,氣候因子和網絡復雜性共同解釋了13%,土壤因子和網絡復雜性共同解釋了1%。微生物網絡復雜性與環境因子交互作用對EMF的貢獻率高于多樣性;微生物網絡復雜性、氣候因子和土壤因子三者共同解釋了41%的EMF差異,高于微生物多樣性、氣候因子和土壤因子三者共同作用。以上結果表明微生物網絡復雜性與非生物因子較微生物多樣性與非生物因子對EMF的解釋度更好(圖8)。 圖8 微生物參數(微生物多樣性和網絡復雜性)、氣候和土壤因子解釋生態系統多功能性變異百分比的方差分解分析 使用SEM進一步分析環境因子、微生物屬性和EMF間的作用關系,SEM解釋了EMF變化的83%。在考慮年均溫、土壤含水量、土壤黏粒含量、土壤鹽基離子和酸性離子影響時,細菌和真菌網絡復雜性仍直接正向影響EMF;微生物多樣性通過正向促進網絡復雜性間接影響EMF。年均溫、土壤酸性離子和土壤鹽基離子環境因子主要通過影響微生物網絡復雜性間接影響EMF,其中土壤酸性離子負向影響細菌網絡復雜性,土壤鹽基離子正向影響網絡復雜性,年均溫正向影響細菌網絡復雜性和真菌網絡復雜性。綜上,環境因子和微生物多樣性對EMF的影響主要通過微生物網絡復雜性實現(圖9)。 圖9 氣候、土壤和微生物屬性預測生態系統多功能性的結構方程模型 研究表明海拔高度涵蓋了降水、溫度、土壤理化性質等諸多環境因素的綜合效應[33, 82],本研究中年均溫、土壤可交換陽離子和土壤質地等環境因素隨海拔高度變化,符合上述研究結果。SM隨海拔的升高而升高,與張鵬等[83]在藏東橫斷山區的研究結果一致。這是由于海拔會影響水熱分配,高海拔地區土壤溫度低,降水和冷凝水多,導致SM隨海拔高度的升高而升高[84]。與西藏色拉山[85]的試驗結果一致,我們的研究結果表明土壤pH隨海拔的升高而下降。土壤的酸化往往促使土壤固相鋁轉變成交換性Al3+,因此土壤Al3+含量隨海拔的升高而升高[86]。此外,本研究中的土壤Ca2+、Mg2+、Na+含量隨海拔的升高而下降,其原因可能是隨海拔升高而降低的土壤pH改變了土壤酸緩沖體系,使得高海拔主要通過消耗土壤鹽基離子發揮酸緩沖作用,導致土壤Ca2+、Mg2+、Na+含量隨海拔的升高呈下降趨勢[87]。但K+與Ca2+、Mg2+、Na+鹽基離子含量的變化趨勢相反,這可能是由于K+含量主要受成土母質的影響[88]。本研究區高海拔成土母質為石灰巖風化物,石灰巖母質發育而來的土壤K+含量較高[89],導致高海拔K+含量高。此外,本研究中土壤鹽基離子含量呈現Ca2+>Mg2+>K+>Na+,符合一般規律,而且土壤交換性Ca2+在4個鹽基離子中占比最大[90]。 環境因子會影響微生物群落組成以及微生物間的相互作用[27, 34—35],本研究結果表明年均溫、酸性離子、鹽基離子和黏粒含量是微生物多樣性和網絡結構復雜性的重要環境驅動因子。其中,年均溫對微生物多樣性和網絡復雜性的影響最顯著,這可能是由于年均溫與微生物的代謝過程和生化過程緊密相關,導致年均溫對微生物影響最明顯[91]。一項位于高草草原生態系統的原位模擬增溫研究發現,增溫提高了微生物分類多樣性[92]。該研究認為溫度是起著決定性作用的環境過濾因子,可對微生物進行選擇,例如升溫促進碳利用率更高的微生物生長,降低微生物群落演替過程的隨機性,使群落發生趨異演替(Divergent succession),從而改變微生物群落結構[92]。酸性離子則可能抑制細胞外酶活性,使得微生物多樣性和網絡復雜性發生改變[93—94]。對臥龍自然保護區真菌群落的研究和東北地區細菌群落的研究均表明,土壤pH是土壤真菌和細菌群落組成與多樣性的重要影響因子[94—95]。隨著土壤酸化程度的加劇,土壤固相鋁會轉變成交換性Al3+,增多的Al3+會抑制胞外酶活性并且對土壤微生物產生生理脅迫,從而改變微生物群落結構[81, 93]。Ca2+和Mg2+的減少則會使微生物對環境脅迫的敏感性增加,使土壤微生物豐度減少[60]。張廣帥等[96]對土壤微生物群落結構與環境因子關系的研究也發現,土壤Ca2+和Mg2+對土壤微生物群落結構具有顯著性影響。此外,一項關于高寒草甸的觀察研究發現,土壤黏粒影響了微生物數量[97]。因此,隨著海拔的升高,年均溫、鹽基離子、酸性離子和土壤質地可能通過調節土壤中微生物的代謝活動或聚集情況,影響微生物多樣性和共現網絡的形成。 許多微生物多樣性的控制試驗結果表明多樣性與生態功能間呈正相關關系,較高的多樣性可為各種時空環境條件下功能的維持提供更大的保障[14, 23, 98—99]。本研究發現,盡管土壤真菌和細菌多樣性與EMF存在顯著正相關關系,但在排除同時影響微生物多樣性和EMF的環境因素后,細菌和真菌多樣性與EMF不存在顯著相關關系。上述結果表明,自然界中環境因素很大程度上影響了土壤微生物多樣性與EMF的聯系。自然生態系統具有空間和時間尺度的環境異質性,環境因素會同時影響微生物群落和生態系統功能,進而影響土壤微生物多樣性與生態功能的關系,例如,環境因子同時正向或負向影響微生物多樣性和生態功能會導致兩者出現正向關系,若對兩者產生相反的影響則會導致兩者出現負向關系[100]。本研究中的年均溫、鹽基離子和酸性離子均正向影響細菌多樣性和EMF,黏粒含量和鹽基離子正向影響真菌多樣性和EMF,導致細菌和真菌多樣性與EMF出現正向關系。此外,本研究通過控制影響微生物復雜性和EMF的環境因子后,發現兩者關系不同于微生物多樣性與EMF的關系,微生物網絡復雜性與EMF關系的顯著正相關關系在控制環境因子后仍然存在。一項微生物多樣性控制試驗結果也表明,網絡復雜性與生態系統功能正相關[19]。這可能是由于生態系統生態功能并不是通過物種單獨作用維持,而是由不同物種間的相互聯系(即互作網絡)共同維持[19]。鑒于微生物群落間的關聯是生態系統物質循環、能量流動和信息傳遞等基礎功能的基礎[19—20],因此,生態系統土壤養分有效性、碳循環和氮循環等諸多生態過程是土壤微生物間相互作用、協同驅動的結果[101]。有研究表明,即使單一生態過程也需要多個微生物類群通過彼此間相互作用共同驅動[101]。例如,異養細菌群落間的相互作用可提高有機碳分解和氮礦化速率[101];纖維素的分解依賴疣微菌門(Verrucomicrobia)、綠彎菌門(Chloroflexi)和浮霉菌門(Planctomycetes)等微生物間的協同作用[102]。由此可見,個體微生物間網絡復雜性的提高可能意味著彼此間功能聯系的增加,網絡復雜性的增加可提高生態系統能量流動和物質循環的效率,進而對EMF產生正向影響[20]。因此,相較于微生物物種多樣性,網絡復雜性與EMF的聯系更加緊密。 SEM結果進一步顯示細菌和真菌多樣性通過促進相應的網絡復雜性間接正向影響EMF。雖然物種多樣性并不能直接反映微生物所發揮的生態功能,但較高的物種多樣性可以促進微生物網絡的復雜程度,進而使得微生物間的生態聯系越緊密[19, 103]。綜上所述,微生物網絡復雜性比物種多樣性能更好地預測EMF,較高的微生物物種多樣性通過促進網絡復雜性維持生態系統功能。 沿青藏高原高寒草甸海拔高度,研究土壤微生物多樣性和網絡結構復雜性對生態系統多功能性的影響發現:相較于微生物物種多樣性,網絡復雜性可更好地預測生態系統多功能性;微生物多樣性通過促進網絡復雜性間接正向影響生態系統多功能性。因此在進行土壤微生物多樣性與生態功能關系的研究時,有必要將微生物網絡復雜性納入。本研究的結果擴展了BEF關系的研究,有利于加深對高寒草地生態系統生態功能微生物調控機制的認識。
2 結果與分析
2.1 不同海拔高度環境因子的特征
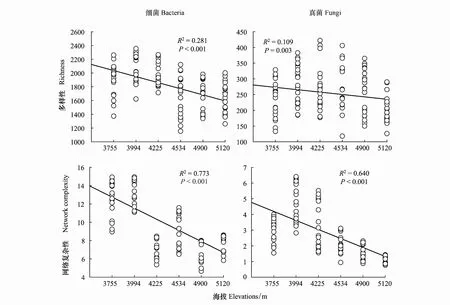
2.2 不同海拔高度微生物共現網絡和屬性的變化
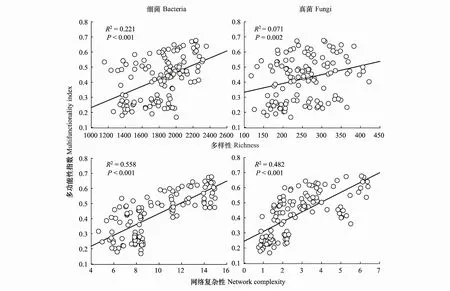
2.3 微生物多樣性、網絡復雜性與生態系統多功能性的關系


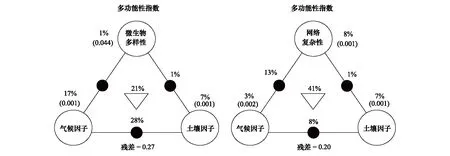
2.4 環境因子和微生物參數對生態系統多功能性的影響

3 討論
3.1 環境因子隨海拔高度的變化
3.2 環境因子對微生物屬性的影響
3.3 微生物物種多樣性和網絡復雜性對生態系統多功能性的影響
4 結論
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
領導文萃(2015年4期)2015-02-28 09:19:05
