華東葡萄抗逆相關基因VpSBP16原核表達及功能初步分析
2022-04-28 09:03:38柴生樾王西平
陜西農業科學 2022年3期
柴生樾,王 浩,王西平
(1.西北農林科技大學 園藝學院,陜西 楊凌 712100;2.旱區作物逆境生物學國家重點實驗室,西北農林科技大學,陜西 楊凌 712100)
隨著葡萄(VitisviniferaL.)在世界范圍內的栽培面積越來越廣,其果實品質口感及經濟價值成為了我們更加關注的話題。當今時代生物信息學和分子生物學迅速發展,利用現代分子生物技術快速提高葡萄的內在及外在品質,培育葡萄優良品種具有很大的意義及應用價值。SQUAMOSA啟動子結合蛋白近年來受到了越來越多的關注,它是SBP基因編碼的一種植物特異性的轉錄因子,包含一個大約76個氨基酸大小的SBP保守結構域和一個特殊的鋅指結構,在蛋白與DNA結合的過程中起著重要的作用[1~3]。SBP-box基因最早是在金魚草(Antirrhinummajus)的花序中被分離的,研究發現它通過調節MADS-box基因而結合在SQUAMOSA的啟動子上,從而控制早期花的發育[3]。之后,擬南芥(Arabidposisthaliana)[4],單細胞藻類衣藻(Chlamydomonasreinhardtii)[5],水稻(Oryzasativa)[6],小立碗蘚(Physcomitrellapatens)[7],玉米(Zeamays)[8]和葡萄(Vitisvinifera)[9]中的SBP-box基因相繼被發掘出來。大豆中SBP-box基因成員數量最多,有111個[10~11],蘋果中有SBP-box基因數量有42個[2]。而在擬南芥中有30個SBP-box基因,如AtSPL3可以調節擬南芥的芽發育[12],水稻中29個SBP-box基因,如水稻的OsSPL16是粒度、形狀和品質的調節劑[13~14]。
根據對已經發現SBP-box基因的植物進行功能研究發現,SBP-box基因在增強植物生物及非生物脅迫抗性方面起著重要的作用。銅是有氧生物中必須的金屬元素之一,故其代謝受到生命體嚴格的控制。銅信號傳導的調控位點CRR1(銅響應調節劑)有一個名為SBP的植物特異性DNA結合域,是激活和抑制衣藻中銅和低氧傳感途徑的靶基因所必需的。研究表明具有SBP結構域的家族成員AtSPL7激活了參與銅的多個基因的轉錄,包括miR398,一些銅轉運蛋白和CCH,是擬南芥中銅穩態的中央調節劑[15]。擬南芥AtSPL14參與對伏馬毒素B1的敏感性而控制細胞凋亡[16]。此外SBP基因調控植物生長及發育[17],包括植物葉、花、果實形成等方面。研究發現SBP基因參與植物葉的形態建成并且是葉原基形成間隔期調控的基因[18]。另外SBP基因在植物花的形成及發育[19~20]方面也起著重要作用,可能促進了植物營養生長與生殖生長間的轉換[21]并且影響著果實的發育與成熟[22~23],此外還涉及信號轉導[24]等諸多方面。隨著生物信息學的發展及葡萄基因組測序數據的完成,已經在葡萄中發現了18個SBP基因家族成員[9]。但在葡萄中對SBP-box基因的功能研究卻鮮有報道。我們初步推測,葡萄SBP轉錄因子與已知的其他植物功能相似,可能在抗非生物脅迫方面、植物體生長發育及果實成熟方面起著重要作用。筆者實驗室前期通過在擬南芥中過表達VpSBP16基因并進行干旱脅迫及高鹽處理,結果表明在擬南芥中過量表達VpSBP16可能通過調節SOS信號和ROS信號來增加幼苗和成熟植株對鹽和干旱脅迫的抗性[25]。為了更深入的研究SBP基因家族,揭示該基因家族的生物學功能及分子調控機制,筆者研究對VpSBP16基因進行原核表達,通過制備VpSBP16基因抗血清并對葡萄不同組織進行蛋白表達,這有利于對葡萄SBP基因家族進行更加全面的了解和分析,為進一步研究該基因家族提供了更有價值的信息。
1 材料與方法
1.1 材料
試驗所用的葉片、花序、莖、卷須以及不同時期的果實均采自種植于西北農林科技大學葡萄種質資源圃的華東葡萄(V.pseudoreticulata)‘白河-35-1’及毛葡萄(V.quinquangularis)‘商-24’;新西蘭雄性大白兔購于西北農林科技大學種兔場;大腸桿菌菌株Escherichia.ColiDH5α、Rosetta(DE3)、原核表達載體pGEX-6p-1均由西北農林科技大學旱區作物逆境生物學國家重點實驗室保存;DNA聚合酶、DNA、蛋白質Marker、限制性核酸內切酶、均購自大連TaKaRa公司;連接試劑盒及質粒小提試劑盒由北京天根公司提供;增強型HRP-DAB底物染色試劑盒與抗GST鼠源單克隆抗體分別購于北京天根公司和南京金斯瑞公司。弗氏完全和不完全佐劑、CAPS、氨芐青霉素、牛血清蛋白、IPTG為美國Sigma公司產品。辣根過氧化物酶標記羊抗兔IgG抗體及辣根過氧化物酶標記羊抗鼠抗體分別購自為美國ABGENT公司和美國JACKSON公司。
1.2 方法
1.2.1 轉錄因子VpSBP16在大腸桿菌中的轉化及融合蛋白誘導表達將構建好的T-Easy-VpSBP16與pGEX-6p-1載體用BamHI、XhoI雙酶切后,回收產物過夜連接,轉化大腸桿菌DH5α,菌液PCR鑒定測序結果正確后,提取pGEX-VpSBP16重組質粒,并進行酶切檢測。將重組質粒轉化表達菌株Rosetta(DE3),轉化方法為熱激法,轉化完成后涂布于含100 mg/L氨芐青霉素的LB平板中,37℃過夜培養。挑取單菌落接種于LB液體培養基,37℃下培養16 h,菌液活化后進行PCR擴增檢測條帶位置是否正確。在超凈工作臺中,將5μL鑒定正確的菌液接種于30 mL LB液體培養基中,于37℃搖床內培養至OD600約為0.4~0.6,再次按1∶100(V/V)的比例吸取菌液加入到新的100 mL LB液體培養基后繼續培養,當OD600為0.6時加入IPTG,用分光光度計測量此溶液終濃度達到0.6 mmol/L,于27℃培養箱誘導培養重組蛋白8 h;取誘導液1 mL,12 000 rmp/min離心1 min,沉淀中加入200 μL 1×SDS-PAGE上樣緩沖液,在開水中煮沸10 min,室溫下12 000 rmp/min離心3 min后吸取20 μL上清溶液上樣,電泳檢測后用考馬斯亮藍染液染色2 h后,加入適量的脫色液脫色對蛋白條帶進行可視化分析。
1.2.2 GST-VpSBP16融合蛋白可溶性分析及純化 取50 mL誘導菌液離心收集菌體,將離心管中的沉淀重懸于5 mL PBS緩沖液中,誘導菌液放置于冰上,超聲波破碎約15 min至菌液粘稠透亮,將全部的細胞裂解物4℃下,9 000 rpm離心5 min分離菌體,將菌液上清吸取到2 mL離心管中,沉淀溶解于PBS緩沖液中充分震蕩溶解。分別取等量上清與沉淀的溶解液,加入等體積上樣緩沖液,通過SDS-PAGE電泳檢測可溶性和不溶性部分的蛋白含量;以轉化空載體的重組菌和未添加IPTG誘導的陽性重組菌為陰性對照。
為了純化重組蛋白,再次將重組菌擴大培養后并用SDS-PAGE電泳檢測。我們使用電透析,切膠的方式純化蛋白,為便于切割必須使目的融合蛋白清晰可見,電泳結束后首先用考馬斯亮藍染色液染色10 min,再將配置好的KCl溶液置于冰上,再次染色直到背景清晰,用手術刀快速精確的將蛋白條帶切于透析袋中。將少量Tris-Glycine緩沖液裝入透析袋放置在水平電泳槽中,90V水平電泳3 h,反向電泳30 min。電泳結束后,取50 mL溶液液氮速凍,-80℃冰箱靜置4 h,于冷凍凍干機凍干48 h。待純化后的蛋白質充分凍干后,將粉末溶解于PBS緩沖液中,測定蛋白質溶液濃度后,通過SDS-PAGE電泳檢測蛋白質是否被純化。純化的蛋白溶液可存放在4℃冰箱內在一周內使用,或者在-80℃冰箱中冷凍保存。
1.2.3 轉錄因子VpSBP16抗血清的制備與Western blot檢測 用PBS緩沖液將保存的純化蛋白稀釋至1 500 mg/mL,取1.5 mL純化蛋白溶液與等體積的完全弗氏佐劑混勻,充分乳化后作為注射所用的抗原。免疫接種選用新西蘭大白兔,每隔10 d進行皮下注射,注射位點選用背部或腹股溝,每次注射400 g,共注射3次。第一次免疫后完全弗氏佐劑換為不完全弗氏佐劑,最后一次免疫7 d后心臟穿刺取血,常規方法獲得抗血清,將獲得的抗血清4℃儲存。
根據美國伯樂半干轉儀的說明,轉膜時轉膜儀上依次為厚濾紙-PVDF膜-膠-厚濾紙,轉膜成功后在搖床上封閉3 h,倒掉封閉液,加入TBST緩沖液,并將所制備的抗血清按1∶20、1∶102、1∶103、1∶104、1∶105倍稀釋作為一抗,一抗孵育完畢后,倒掉溶液,用TBST緩沖液洗脫,再用HRP標記的二抗孵育2 h,倒掉溶液,用TBST緩沖液沖洗三次后按照HRP-DAB底物顯色試劑盒說明進行顯色。
1.2.4 葡萄不同組織總蛋白的提取及VpSBP16蛋白的表達分析 將新鮮的葡萄組織加入液氮預冷的研缽中充分研磨,研磨完成后利用Tris-丙酮-酚法[26]提取葡萄不同組織及果實不同時期中的全蛋白,并在95℃加熱5 min,使用含5%的濃縮膠10 mL,10%的分離膠15 mL,以‘商-24’葡萄的葉、莖、花序、卷須4種組織及果實發育中的6個時期的蛋白為樣品,每個樣品上樣量為30μg,進行SDS-PAGE電泳檢測,一抗添加3 μL的VpSBP16抗血清,Western blot與轉印方法同前。
2 結果與分析
2.1 轉錄因子VpSBP16在大腸桿菌中的轉化及融合蛋白誘導表達
將構建好的重組質粒pGEX-VpSBP16進行菌液PCR擴增和雙酶切鑒定,并用1%的瓊脂糖凝膠電泳檢測獲得了如圖1所示的約1 674 bp的目標片段,序列比對結果正確。重組原核表達載體轉化到大腸桿菌Rosetta中后,將陽性重組菌及對照菌于搖床培養至OD600為0.6時,于0.6 mmol/L的IPTG,27℃誘導培養8h。電泳結果(圖2)表明,IPTG誘導的陽性重組菌高表達出約87 kD的特異條帶(黑色箭頭),與預期大小基本一致。與陰性對照對比可知,轉化空載體的重組菌經誘導后則表達出分子量為26 kD的條帶。這個條帶的大小與GST標簽蛋白大小一致,充分證明了華東葡萄轉錄因子VpSBP16蛋白誘導表達成功。
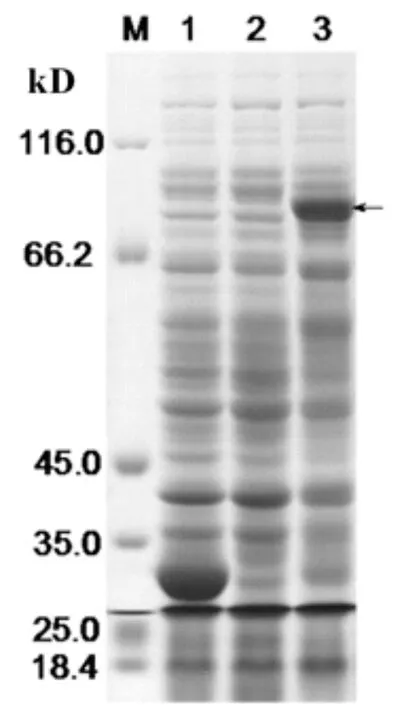
圖2 pGEX-VpSBP16在Rosetta(DE3)菌株中經IPTG誘導的表達產物SDS-PAGE分析
2.2 GST-VpSBP16融合蛋白可溶性分析及純化
SDS-PAGE電泳檢測離心后收集的上清與沉淀,結果表明沉淀中融合蛋白表達量明顯比上清中要高(圖3,泳道5),而在上清中目的蛋白的表達非常微弱(圖3,泳道4),這表明目的融合蛋白主要存在于在沉淀中,并且GST-VpSBP16融合蛋白的主要表達形式為包涵體。通過電透析法,切膠回收法將目的融合蛋白從凝膠中分離經SDS-PAGE發現GST-VpSBP16融合蛋白已經得到較好的純化(圖4)。
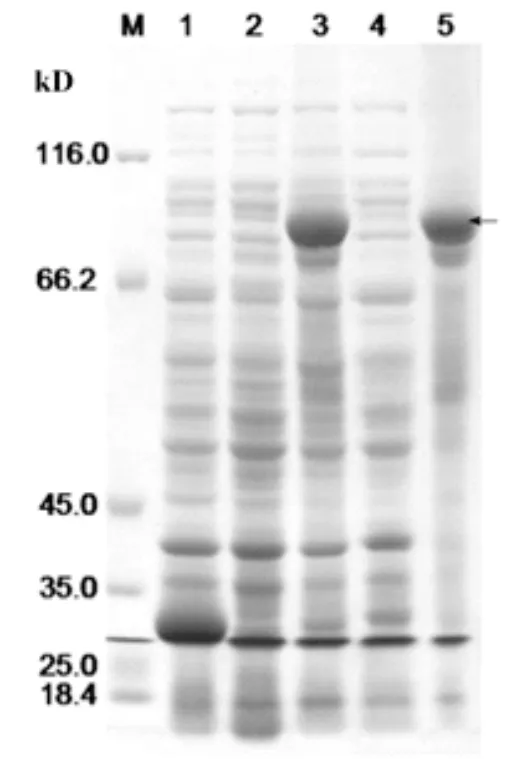
圖3 轉錄因子VpSPB16原核表達蛋白的可溶性分析
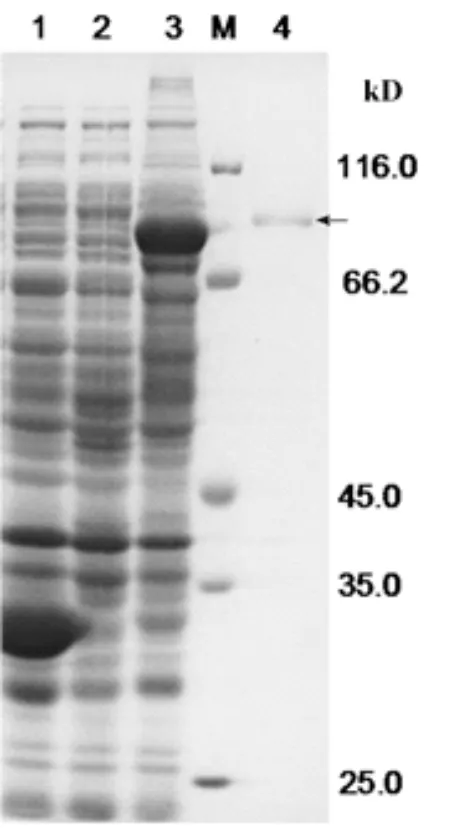
圖4 GST-VpSBP16融合蛋白純化產物的電泳分析
2.3 轉錄因子VpSBP16抗血清的制備與Western blot檢測
抗原選用純化后的GST-VpSBP16融合蛋白注射新西蘭大白兔產生免疫應答,并且產生抗體,從而獲得VpSBP16基因多克隆抗血清。從圖5顯示的結果可以看出,當用不同稀釋倍數的抗血清進行Western blot分析時,均可以產生87KD,清晰且單一的雜交信號,這個結果充分表明以純化蛋白包涵體形式所制備的抗血清可特異性地與抗原產生反應,而且泳道7的結果顯示在稀釋比例達到1:105時依然能夠明顯的檢測到單一的蛋白條帶。陽性對照在檢測目的融合蛋白時以GST鼠源單抗進行孵育,陰性對照選用pGEX-6p-1空載經IPTG誘導后的表達產物并且用目的蛋白抗血清進行孵育。
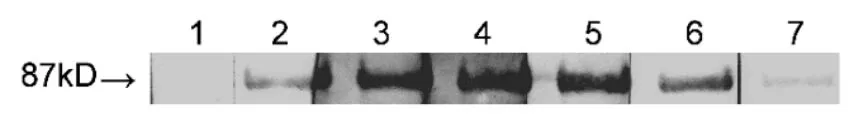
圖5 轉錄因子VpSBP16多克隆抗血清的Western blot分析
2.4 VpSBP16基因在葡萄不同組織中的Western blot檢測
為了進一步驗證所得抗體是否可以檢測到葡萄中的VpSBP16基因表達蛋白,我們提取了毛葡萄‘商24’中5個不同組織的總蛋白,利用Western blot檢測葡萄花,果實,莖,卷須,葉中的VpSBP16蛋白表達情況。結果表明,實驗中制備的抗體可以特異性的與‘商24’中的VpSBP16表達蛋白結合,并在染色后顯示出清晰的蛋白條帶。不僅如此,我們還觀察到在葡萄生長發育時期卷須中的VpSBP16蛋白表達含量最高而花序中的VpSBP16蛋白表達含量最低,在葡萄果實發育時期的綠果期相較于花后45~100 dVpSBP16蛋白表達含量偏高。(如圖6)。
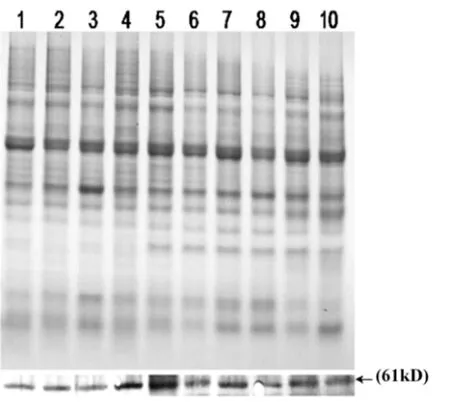
圖6 毛葡萄‘商-24’不同組織中VpSBP16蛋白的表達分析
3 討論與結論
實驗構建表達質粒pGEX-VpSBP16轉化菌株后,在27℃培養的條件下經IPTG誘導8 h可以使融合蛋白穩定表達,純化后的蛋白通過免疫新西蘭大白兔制備抗血清。結果表明我們獲得了純化的目的融合蛋白GST-VpSBP16,還制備了效價高達105的特異性抗血清。研究表明重組菌能否被成功誘導表達,經誘導后表達產物的多少及存在形式是制備抗體的重要環節,故表達載體、菌株選擇、誘導時間和誘導物IPTG濃度都是實驗中需要考慮的重要因素[27]。研究選用pGEX-6p-1載體是由于該載體上含有一段GST標簽,使得融合蛋白的產量和可溶性得到提高,其次便于檢測和純化目的融合蛋白。Rosetta(DE3)菌株具有眾所周知的遺傳背景,并包含對氯霉素具有抗性的質粒[28]有利于外源基因在大腸桿菌內的順利表達。采用27℃過夜誘導的表達條件,是因為在22~37℃的范圍之內降低誘導溫度可提高可溶性蛋白的含量[29]。根據前人研究經驗,宿主細菌蛋白酶隨誘導時間的延長會對表達蛋白質的降解作用增加,所以達到蛋白表達最大量后蛋白增加將不明顯[30],故筆者實驗采用IPTG濃度為0.6 mmol/L,8 h過表達目標融合蛋白,根據Western blot檢測結果顯示該條件足以滿足后期蛋白質回收,純化和抗原制備的要求。由于包涵體蛋白復性會造成二硫鍵錯配使蛋白質失活[30],所以研究采用切膠,電透析的方式獲得高效價的純化蛋白。
SBP轉錄因子家族具有一個共同的脫氧核糖核酸結合域(SBP域)和一個不同尋常的鋅指結構,這一類轉錄因子中的許多基因被認為在植物生長發育及抗非生物脅迫中起著重要的作用[9]。筆者實驗室前期在擬南芥上過表達VpSBP16基因發現該基因可以提高擬南芥對干旱及高鹽脅迫的抗性。為深入探索VpSBP16的功能,筆者試驗制備VpSBP16多克隆抗血清并運用Western blot技術對毛葡萄果實,花序,莖,葉,卷須等不同器官的目的蛋白表達量進行檢測,結果顯示VpSBP16多克隆抗血清可以與目的基因蛋白雜交出特異性的條帶,這表明我們所制備的多克隆抗血清可以運用于后續VpSBP16功能研究試驗。通過觀察VpSBP16蛋白表達條帶發現花序的蛋白表達量偏低,而花后15~30 d的綠果期含量偏高,這與之前報道的SBP-box基因家族影響果實發育和成熟的結論一致[22],可能暗示著VpSBP16轉錄因子同樣也參與葡萄果實成熟的調控機制,且在葡萄從營養生長到生殖生長的轉換過程中發揮著重要作用。此外,在葡萄生長發育時期,卷須中的VpSBP16蛋白表達含量最高可能暗示著VpSBP16轉錄因子調控葡萄卷須的生長發育,具體調控機理需進一步深入研究。
總之,我們獲得了具有生物活性的重組蛋白,制備了VpSBP16基因的多克隆抗血清,它能夠特異的識別天然的VpSBP16表達蛋白,在蛋白水平上利用此抗血清對毛葡萄中VpSBP16基因功能進行了進一步的探索,為下一步開展VpSBP16基因功能的深入研究奠定了理論基礎,為研究葡萄SBP家族的分子機制及蛋白功能提供了依據。
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
今日農業(2021年19期)2022-01-12 06:16:36
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年11期)2021-08-22 03:15:44
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
無線電工程(2020年11期)2020-10-29 01:25:46
