山藥品種蘇蕷8號抗炭疽病基因的轉錄組測序與表達分析
2022-05-12 10:58:26韓曉勇殷劍美張培通
華北農學報 2022年2期
關鍵詞:途徑
韓曉勇,蔣 璐,殷劍美,金 林,張培通
(江蘇省農業科學院 經濟作物研究所,江蘇 南京 210014)
炭疽病是為害山藥最為嚴重的病害之一,如果未及時有效控制,其損失可達80%以上,甚至絕收,嚴重制約著山藥產業發展。山藥炭疽病主要為害山藥葉片,也可為害莖蔓,其病癥狀復雜,一般先從植株下部開始發病。化學藥劑是防治山藥炭疽病的主要手段,但易帶來環境污染、生態失衡及食品安全等問題,而且頻繁使用農藥可能會導致致病菌株變異,出現致病性更強的毒株[1]。培育抗炭疽病的山藥品種是控制田間山藥病害最有效和理想的方法,但山藥以營養繁殖為主,存在生理限制。盡管通過常規育種在培育抗炭疽病雜交種方面取得了一些進展[2],但進展仍然緩慢。傳統技術和分子技術的結合將是開發具有穩定抗性山藥品種的更好方法,以保護山藥免受廣譜真菌病原體的侵害,從而改善山藥品質。近年來,基因組學輔助育種(GAB,Genome-assisted breeding)已成為一種強有力的植物育種策略,它將基因組學與高通量表型分析相結合,以改良作物[3]。
炭疽病抗性基因的挖掘及分子標記輔助選擇,是培育山藥廣譜、高效和穩定的抗病品種的重要途徑。通過對易感基因型(TDa95/0310)和2種炭疽病抗性山藥基因型(TDa87/01091和TDa95/0328)的比較轉錄組學分析,TDa95/0328和TDa87/01091分別有115,180個高表達的EST,它們與碳水化合物代謝、細胞壁生物合成、脂質和氨基酸代謝、次生和激素代謝、轉錄因子、蛋白質合成以及多種病程相關信號蛋白和宿主防御相關基因有關[4-5]。Upadhyaya等[6]利用14 739個SNP標記定位到了8個與高粱抗炭疽病有關的位點。Yang等[7]通過圖位克隆的方法從苜蓿中克隆獲得RCT1基因,該基因屬于TIR-NBS-LRR類基因,將RCT1轉化到對炭疽病敏感的苜蓿植株中后,轉基因植株表現出對炭疽病的廣譜抗性。采用同源克隆和RT-PCR方法,從草莓久香克隆到FaNBS20全長cDNA序列,草莓抗炭疽病品種甜查理接種草莓炭疽病菌后,FaNBS20的表達水平顯著提高,而在感病品種久香FaNBS20的表達則一直處于較低水平,表明該基因可能與炭疽病的抗性相關[8]。編碼ABA-響應蛋白、亮氨酸富集蛋白、黃酮-3-羥化酶及谷胱甘肽-S-轉移酶的基因在抗炭疽病高粱植株內被明顯誘導表達但在易感植株內幾乎檢測不到,表明這些基因在植株抗炭疽病方面可能發揮著非常重要的作用[9]。目前,利用RNA-seq技術挖掘山藥炭疽病抗性基因尚未見報道。
本試驗利用RNA-seq技術分析不同時期高抗和高感品種的山藥葉片受炭疽菌侵染后的基因表達變化,挖掘山藥參與炭疽病抗性相關基因,對關鍵抗性基因進行表達分析,構建山藥響應炭疽病侵染的分子途徑,旨在為揭示山藥抗炭疽病的分子機制,培育山藥廣譜、高效和穩定的抗病品種奠定基礎。
1 材料和方法
試驗于2019—2021年在江蘇省農業科學院經濟作物研究所實驗室完成。
1.1 試驗材料及處理
1.1.1 試驗材料 膠孢炭疽菌菌株4-2由筆者保存于本實驗室。山藥品種蘇蕷8號,是江蘇省農業科學院利用浙江省臺州市地方紫山藥品種臺州紫山藥經系統選育的高感炭疽病品系024,組培誘變所形成的品種,其薯形好,產量高,高抗炭疽病。用于差異基因表達分析的高感品系024和江西短紫山藥、高抗品種蘇蕷8號和蘇蕷6號、中抗品種紅河山藥均種植于江蘇省農業科學院南京市六合試驗基地。當年繁殖的蘇蕷8號和品系024的種薯,次年用作接種材料。
1.1.2 試驗處理 選取蘇蕷8號和品系024的健康、無病、大小一致的種薯,播種于鋪滿基質的花盆中,置于恒溫恒濕的培養環境中培養。待植株長至5~6葉期,用2 mm左右接種針針刺展開葉葉片,將PDA平板培養7 d后的菌株用無菌水配制成濃度為1×106個/mL的孢子懸浮液,均勻噴霧于植株葉片并遮防護罩保濕,24 h后摘罩。為防止誤差,每片葉針刺2孔,葉片正面和背面分別噴霧2次孢子懸浮液。收集蘇蕷8號接種0,24,48,72,96 h(R0h、R24h、R48h、R72h、R96h),品系024接種24,48,72,96 h(S24h、S48h、S72h、S96h)的葉片,液氮速凍后置于-80 ℃冰箱中保存。每個樣品為3~5片葉的混合樣,每個處理3個生物學重復。
1.2 RNA-seq測序及原始數據處理、分析
基于Illumina HiSeq 2500測序平臺對構建好的cDNA文庫進行測序,測序工作由南京集思慧遠生物科技有限公司完成。測序得到的原始圖像數據文件,經堿基識別(Base Calling)分析轉化為原始測序序列(Raw Data或Raw Reads),用Sanger質量值Q來評估原始數據的測序質量,獲得Clean data。經De novo組裝后,獲得山藥的Unigene庫。
1.3 差異基因的篩選和功能注釋
采用Bowtie 2將測序得到的Reads與Unigene庫進行比對,根據比對結果,結合RSEM進行表達量水平估計。利用FPKM值計算各樣品中每個基因的表達量。采用DESeq 2軟件進行2組樣品間的差異表達分析,將|Fold Change|≥4且 FDR<0.05作為篩選標準。篩選獲得的差異基因進行GO功能富集分析和KEGG Pathway富集分析,以P<0.05為閾值。
1.4 Q-PCR驗證
選取3個差異表達基因,利用Oligo 6軟件設計引物,以Actin基因為內參(表1),用轉錄組測序同批次采集樣品的cDNA為模板進行Q-PCR,驗證轉錄組測序數據的可靠性。設3次生物學重復,按照2-ΔΔCt算法計算基因的相對表達量。
1.5 差異基因表達分析
在炭疽病發病盛期,分別取品系024、江西短紫山藥、蘇蕷8號、蘇蕷6號和紅河山藥的3~5片展開葉,液氮速凍后置于-80 ℃冰箱中保存。每個品種3個生物學重復,其中,品系024和江西短紫山藥的感病葉和健康葉分別進行取樣保存。
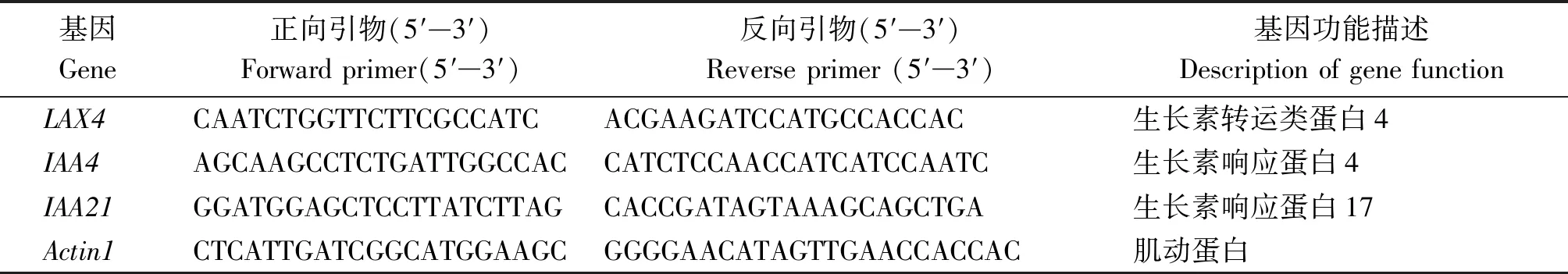
表1 Q-PCR基因及引物Tab.1 The genes and primers used for Q-PCR
2 結果與分析
2.1 轉錄組測序結果質量評估
去除含有接頭的、重復的、質量低的片段,9個樣品獲得Clean reads 在18 538 496~20 504 603條(表2)。各樣品Clean data均達到5 561 548 900 bp及以上,GC含量均高于44%,N堿基含量均為0,Q30堿基百分比均在90%以上,表明各樣品測序所得Clean data的堿基組成基本平衡且大部分reads的堿基質量值大于30,符合質量錯誤率低于1%的要求。轉錄組測序質量較好,數據可靠。

表2 測序數據統計結果Tab.2 The statistics of transcriptome sequencing data
2.2 差異表達基因分析
建立抗病品種接種不同時間差異基因集,同時建立接種相同時間的抗感病品種差異基因集,選擇共同差異表達的基因,結果表明(圖1):抗感品種間與抗病品種內接種72,96 h差異表達的基因數最多,分別為3 814,1 630個。差異表達基因韋恩圖結果表明(圖2),接種24,48,72,96 h差異表達的基因個數分別為197,132,187,313個,去除各接種時間點共同差異表達的基因數,共獲得711個差異表達的基因。
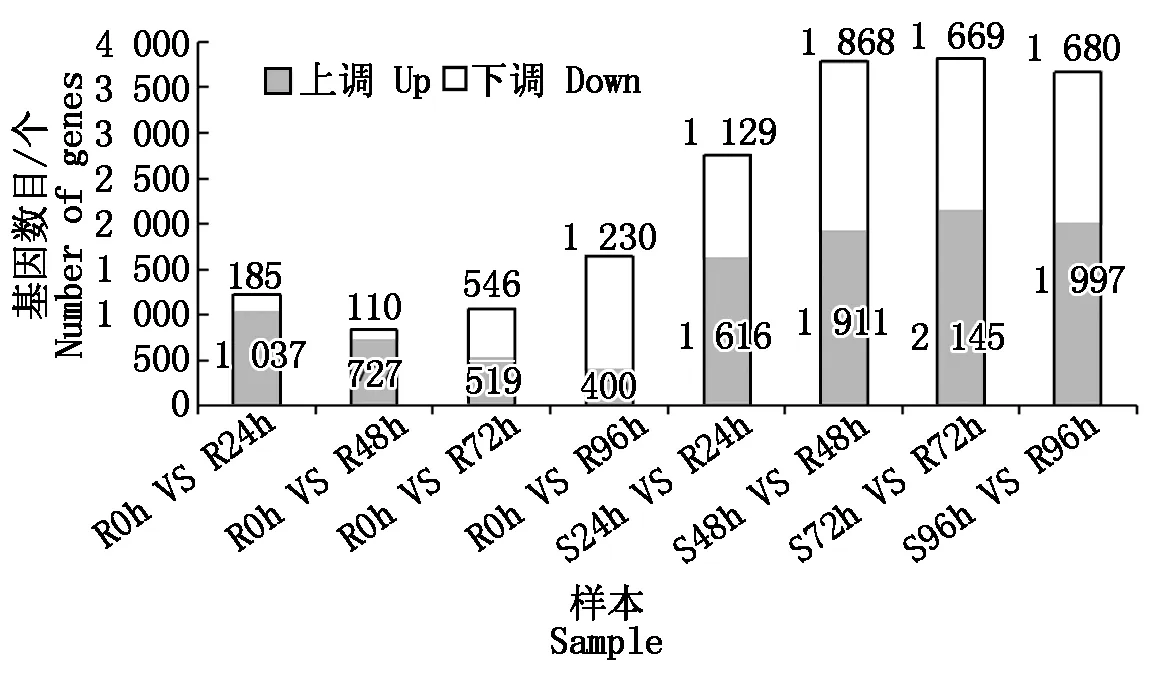
圖1 各樣本間差異表達基因數Fig.1 The statistics of differentially expressed genes between samples
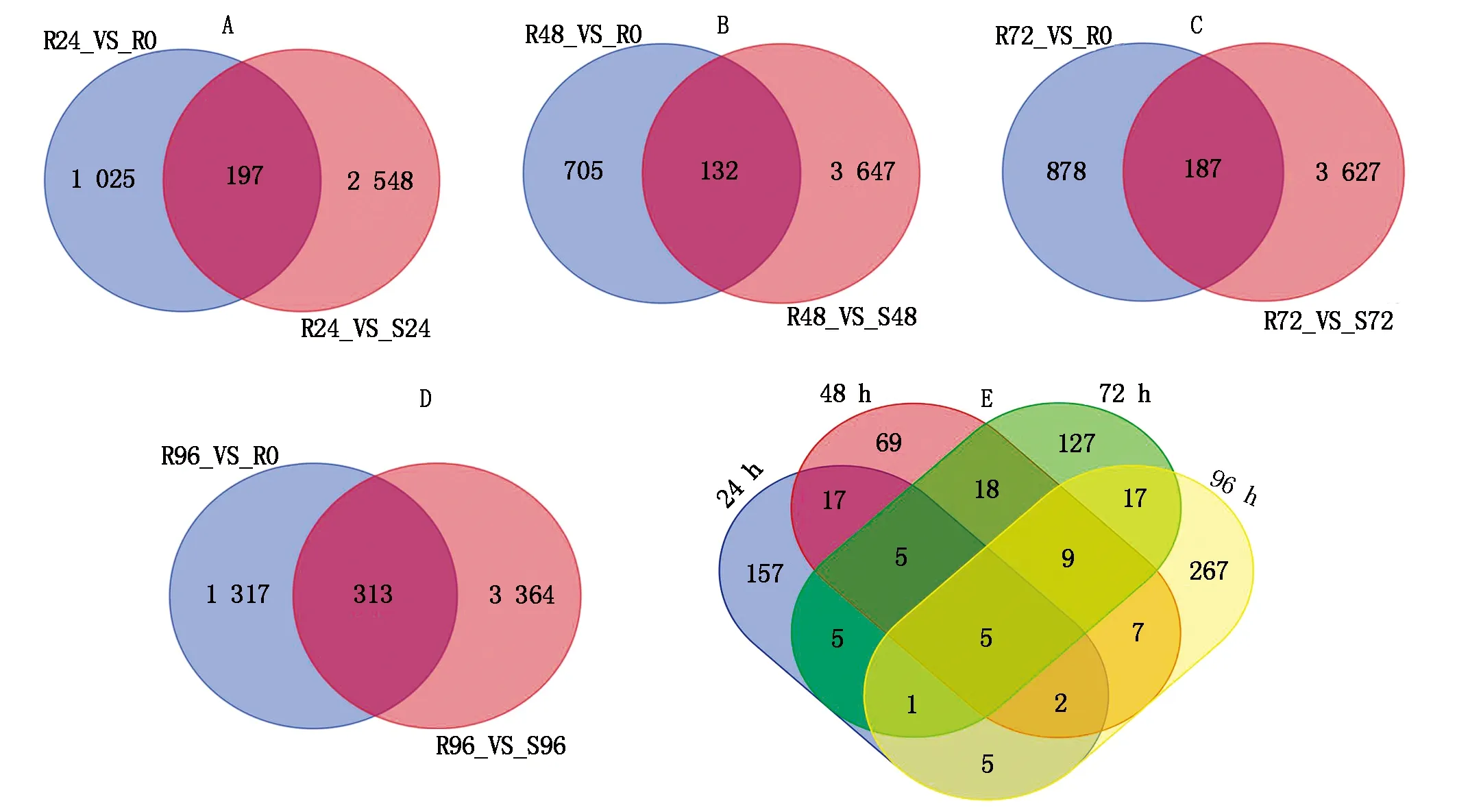
A—D.分別為接種24,48,72,96 h差異表達基因韋恩圖;E.接種24,48,72,96 h共同差異表達基因韋恩圖。A—D.Venn map of differentially expressed genes after inoculation 24,48,72,96 h,respectively;E.Venn map of common differentially expressed genes after inoculation 24,48,72,96 h.
2.3 差異表達基因的GO功能富集分析
差異表達的基因GO功能分類結果表明:有666個基因被注釋,分為生物過程、細胞組分和分子功能三大類和50個亞類(圖3),其中生物過程包含20個亞類,基因數最多的5個亞類依次是細胞過程、代謝過程、單組織過程、刺激響應和生物調節;細胞組分包含15個亞類,基因數最多的2個亞類是細胞和細胞部分;分子功能包含15個亞類,基因數最多的2個亞類是結合和催化活性。差異表達基因GO功能分類基因數占比與背景基因GO功能分類基因數占比趨勢基本一致。

BP.生物過程:BP1.細胞過程;BP2.代謝過程;BP3.單組織過程;BP4.刺激響應;BP5.生物調節;BP6.定位;BP7.細胞組分組織或生物發生;BP8.發展過程;BP9.信號;BP10.多細胞生物過程;BP11.生殖;BP12.生殖過程;BP13.多組織過程;BP14.免疫系統過程;BP15.生長;BP16.脫毒;BP17.節律過程;BP18.運動;BP19.細胞殺傷;BP20.生物附著。CC.細胞組分:CC1.細胞;CC2.細胞部分;CC3.細胞器;CC4.膜;CC5.細胞器部分;CC6.膜部分;CC7.大分子復合物;CC8.胞外區;CC9.膜包圍腔;CC10.細胞連接;CC11.超分子復合物;CC12.胞外區部分;CC13.擬核;CC14.其他生物體;CC15.其他生物體部分。MF.分子功能:MF1.結合;MF2.催化活性;MF3.轉運活性;MF4.結構分子活性;MF5.信號轉導活性;MF6.核酸結合轉錄因子活性;MF7.分子轉導活性;MF8.分子功能調節器;MF9.電子載體活性;MF10.抗氧化活性;MF11.營養庫活性;MF12.轉錄因子活性、蛋白質結合;MF13.金屬伴侶蛋白活性;MF14.翻譯調節活;MF15.蛋白標簽。
GO功能顯著性富集分析表明(圖4),富集于生物過程途徑的基因數最多,為116個,其中顯著富集于響應生物刺激(GO:0050896)和應激反應(GO:00006950)terms的基因數分別為63,44個;其次是富集于分子功能途徑的基因,為79個;富集于細胞組分的基因數最少,為31個。前20條顯著富集的terms中(圖5),4個差異基因富集于生物刺激響應term,且富集程度最高;防衛反應term富集基因數最多,為13個;此外,有些基因富集于細胞壁修飾、氧化還原過程和催化酶活性,這些基因可能均與植物抗病有關。
2.4 差異表達基因的KEGG富集分析
利用KEGG數據庫對篩選出的差異基因進行了功能富集分類和Pathway注釋(圖6):有40個基因富集于31條信號通路,其中23條參與代謝途徑,3條參與遺傳信息處理,分別有2條參與環境信息處理和細胞進程,1條參與有機體系統。糖酵解/糖異生代謝途徑富集的基因數最多,為6個,其次是植物激素信號轉導途徑、戊糖和葡萄糖醛酸相互轉化途徑,均為5個。

圖6 差異表達基因KEGG富集分類Fig.6 KEGG enrichment classification of the differentially expressed genes
對所有富集的KEGG Pathway進行篩選,共獲得10條顯著富集的 Pathway(表3)。其中5條涉及糖代謝途徑,分別是戊糖和葡萄糖醛酸相互轉化、糖酵解/糖異生、半乳糖代謝、淀粉與蔗糖代謝和磷酸戊糖途徑;2條涉及次生代謝產物途徑,分別是類固醇生物合成途徑和托烷、哌啶、吡啶生物堿合成途徑;分別有1條涉及植物激素信號轉導途徑、甘油酯代謝和生物素代謝途徑。
2.5 山藥響應炭疽菌侵染差異表達基因分類
結合KEGG富集分析對差異表達基因進一步按功能進行分類,主要集中在糖代謝、植物與病原互作、激素代謝、次生代謝等途徑。
2.5.1 淀粉和蔗糖代謝相關基因表達分析 在炭疽菌侵染24 h,與蔗糖分解有關的β-呋喃果糖苷酶(INV)和α-磷酸海藻糖合酶(TPS)、與淀粉合成有關的果糖1,6-二磷酸醛縮酶(FBA-Ⅱ)和磷酸葡糖變位酶(PGM)均上調表達;與淀粉分解有關的α-淀粉酶(Ams)下調表達(圖7-A)。
2.5.2 植物與病原互作途徑相關基因表達分析 在炭疽菌侵染24 h,多個病程相關蛋白下調表達(圖7-B),包括PR1(AOPR1、ACPR1)、PR2(β-1-3-葡聚糖酶,ACPR2、MCPR2、SIPR2)、PR4(幾丁質酶,DBPR4)、PR6(蛋白酶抑制劑,PARR-6、CCPR-6)和PDPR10(海棗核糖核酸酶),而PR5(甜味蛋白)和ACPR-10(菠蘿核糖核酸酶)上調表達;NB-ARC類抗病基因(At1g61310)和NBS-LRR類抗病基因(At4g27220、TW65_03605、TW65_04322、BRADI_1g47610v3、At4g36180、FLS2、EIX2)在炭疽菌侵染24 h均上調表達,其中EIX2在炭疽菌侵染96 h時,表達量最高。參與植物防衛反應的4個熱擊蛋白(HSP12、HSP88、HSP90、HSP70)在炭疽菌侵染24 h均上調表達,其中HSP12和HSP88在炭疽菌侵染24 h表達量最高,而HSP90和HSP70在炭疽菌侵染48 h表達量最高;過氧化氫酶(CAT2)和線粒體超氧物歧化酶(MSD1)在炭疽菌侵染24 h上調表達且表達量最高;谷胱甘肽轉移酶(AOGST)在炭疽菌侵染24 h下調表達。

表3 差異表達基因顯著富集的KEGG通路Tab.3 KEGG pathways significantly enriched of differentially expressed genes
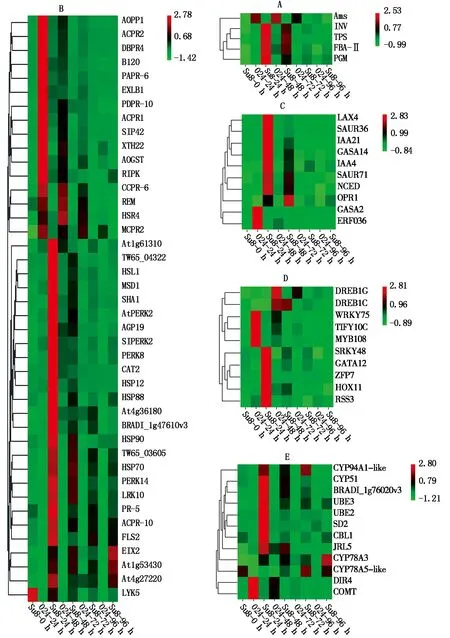
A.植物與病原互作;B.淀粉和蔗糖代謝;C.激素類;D.轉錄因子;E.次生代謝。A.Plant-phytopathogen interaction;B.Starch and sucrose metabolism;C.Hormones;D.Transcription factors;E.Secondary metabolism.
富含脯氨酸的伸展素類受體蛋白激酶(AtPERK2、SIPERK2、PERK8、PERK14),LRR類受體蛋白激酶(At1g53430、LRK10),凝集素受體激酶(HSL1)在炭疽菌侵染24 h上調表達且表達量最高,受體相互作用蛋白激酶(RIPK),溶解素基序型激酶(LYK5),凝集素受體激酶(B120)在炭疽菌侵染24 h下調表達;細胞壁蛋白阿拉伯半乳聚糖蛋白(AGP19)和鈣調素結合蛋白(SHA1)在炭疽菌侵染24 h上調表達,細胞壁修飾的擴展蛋白(EXLB1)、細胞壁降解相關木葡聚糖內糖基轉移酶(XTH22),超敏相關蛋白(HSR4)和植物Remorin蛋白(REM)在炭疽菌侵染24 h下調表達。
2.5.3 激素相關基因的表達分析 在炭疽菌侵染過程中,多個與防御相關的植物激素信號轉導途徑中基因表現出差異,其中生長素信號轉導起主導作用。1個生長素極性運輸載體(LAX4)、4個生長素早期響應蛋白(IAA4、IAA21、SAUR36、SAUR71)在炭疽菌侵染24 h均上調表達且表達量最高;赤霉素調節蛋白(GAGA14、GASA2)在炭疽菌侵染24 h分別上調或下調表達;茉莉酸合成關鍵酶基因12-氧代植二烯酸還原酶(OPR1)和脫落酸合成關鍵酶基因9-順式-環氧類胡蘿卜素雙加氧酶(NCED)在炭疽菌侵染24 h均上調表達,乙烯響應因子(ERF036)在炭疽菌侵染24 h下調表達(圖7-C)。
2.5.4 轉錄因子 轉錄因子在抗病信號轉導途徑中起重要調控作用。在炭疽菌侵染24 h,WRKY類轉錄因子(WRKY48)、GATA類轉錄因子(GATA12)、HD-ZIP類轉錄因子(HOX11)、bHLH-MYC(RSS3)和鋅指蛋白基因(ZFP7)均上調表達,DREB類轉錄因子(DREB1C)在炭疽菌侵染48 h上調表達。DREB類轉錄因子(DREB1G)、WRKY類轉錄因子(WRKY75)、MYB類轉錄因子(MYB108)和TIFY類轉錄因子(TIFY10C)在炭疽菌侵染24 h下調表達(圖7-D)。
2.5.5 次生代謝產物相關基因的表達分析 次生代謝產物作為生活壁壘可以直接參與植物的防御反應,也可以作為信號物質參與植物抗病反應。細胞色素P450基因(CYP94A1-like、CYP78A3、CYP51)、泛素化連接酶(UBE2、UBE3)、防御素(SD2)、幾丁質結合凝集素(CBL1)、木菠蘿凝集素(JRL5)、甾醇C-24甲基轉移酶(BRADI_1g76020v3)在炭疽菌侵染24 h上調表達,其中,CYP94A1-like、CYP78A3分別在炭疽菌侵染72,96 h表達量最高,細胞色素P450基因(CYP78A5-like)在炭疽菌侵染72 h上調表達;參與木質素合成的導體蛋白(DIR4)和咖啡酸3-O-甲基轉移酶(COMT)受炭疽菌侵染后下調表達(圖7-E)。
2.6 差異表達基因的Q-PCR驗證
根據差異基因富集情況,選擇了植物激素信號轉導途徑中與生長素信號相關的基因,通過Q-PCR檢測其表達水平。結果表明(圖8):LAX4、IAA4、IAA21在接種24 h上調表達,與轉錄組測序基因表達變化趨勢一致,表明轉錄組測序結果是可靠的。
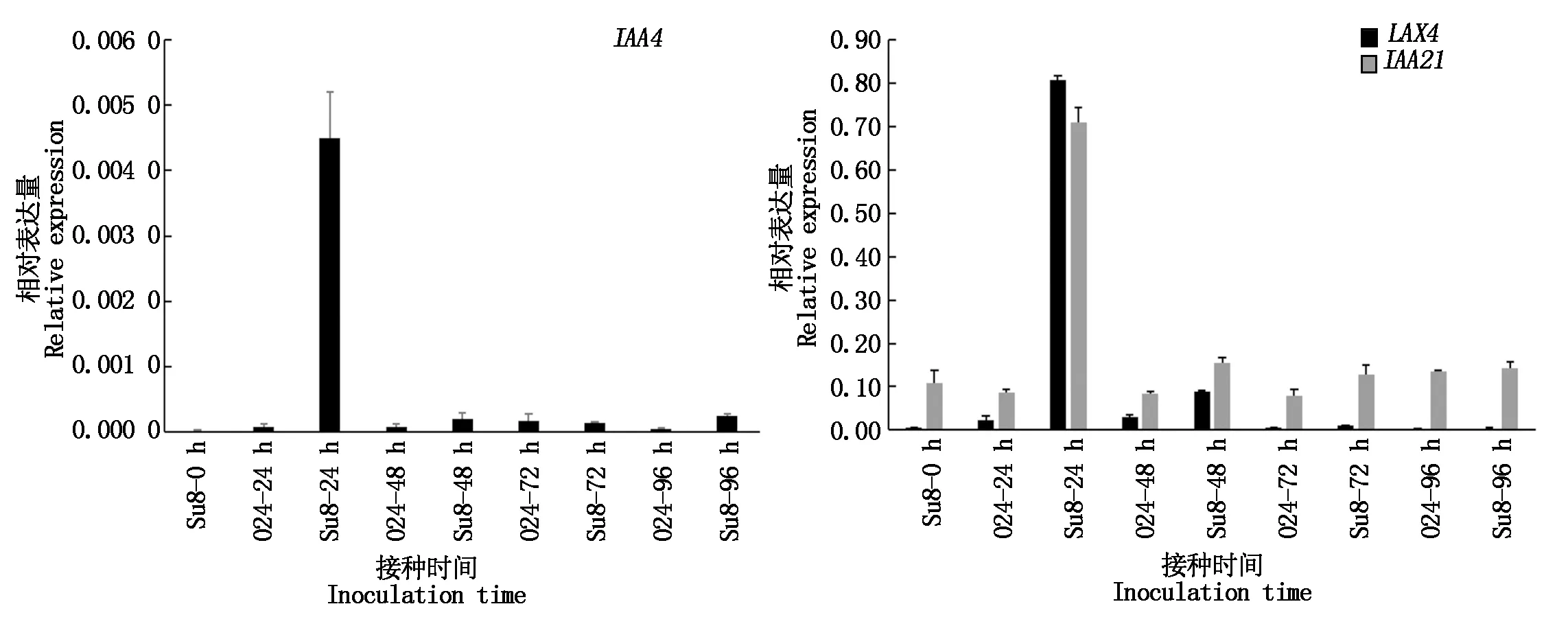
圖8 差異表達基因Q-PCR驗證Fig.8 Q-PCR verification of differentially expressed genes
2.7 差異表達基因的Q-PCR表達分析
為進一步驗證差異基因炭疽病抗性,選擇炭疽病高感品系024和江西短紫山藥、高抗品種蘇蕷8號和蘇蕷6號、中抗品種紅河山藥,進行Q-PCR分析,其中品系024和江西短紫山藥分別對感病葉和健康葉進行表達分析。結果表明(圖9):IAA4、LAX4和IAA21抗病品種表達量顯著高于感病品種,其中,LAX4和IAA21在蘇蕷8號的表達量顯著高于蘇蕷6號和紅河山藥;IAA4在蘇蕷6號的表達量顯著高于其他品種,在蘇蕷8號和紅河山藥表達量無顯著差異。除IAA21江西短紫山藥健康葉表達量顯著高于感病葉外,IAA4和LAX4在感病葉和健康葉表達量無顯著差異,即健康葉不一定健康。

024-S、 024-R、 JXD-S、JXD-R、Su8-R、Su6-R、HH-R分別表示品系024感病葉、健康葉,江西短紫山藥感病葉、健康葉,蘇蕷8號、蘇蕷6號和紅河山藥。024-S, 024-R, JXD-S, JXD-R, Su8-R, Su6-R, HH-R show strain 024 susceptible leaf and healthy leaf,Jiangxi yam susceptible leaf and healthy leaf, Suyu 8, Suyu 6 and Honghe yam.
3 結論與討論
3.1 淀粉代謝參與山藥炭疽病抗性
淀粉代謝異常是植物染病后的主要生理變化之一,通常與淀粉合成有關的酶基因上調表達,地上部出現淀粉積累,地下部發生淀粉欠缺問題[10-11]。蔗糖合酶催化蔗糖降解,是蔗糖向淀粉轉化的第一步[12]。炭疽菌侵染24 h,蘇蕷8號葉片與蔗糖分解有關的β-呋喃果糖苷酶(INV)和α-磷酸海藻糖合酶(TPS)、與淀粉合成有關的果糖1,6-二磷酸醛縮酶(FBA-Ⅱ)和磷酸葡糖變位酶(PGM)均上調表達,而與淀粉分解有關的α-淀粉酶下調表達(Ams),即山藥受炭疽菌侵染后,蘇蕷8號葉片較品系024出現更多的淀粉積累可能。
3.2 防衛基因介導山藥炭疽病抗性
植物在受到病原物侵染時通過誘導產生的防衛基因表達來拮抗,防衛基因按功能劃分,主要分為4類[13]:①病程相關蛋白基因,編碼PR蛋白;②細胞壁修飾有關基因,編碼細胞壁中富含羥脯氨酸的糖蛋白,過氧化物酶及硫堇等;③清除活性氧(ROS)的防衛酶基因,包括超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)等;④次生物質合成基因,編碼植保素及木質素等生物合成所需的酶。不同類別的病程相關蛋白在炭疽病侵染后表達情況不同。NB-ARC類抗病基因、NBS-LRR類抗病基因和熱擊蛋白在炭疽菌侵染24 h全部上調表達,而在11個病程蛋白中,只有PR5和ACPR10上調表達、9個受體蛋白激酶中,6個上調表達,3個下調表達、1個谷胱甘肽轉移酶基因下調表達,這些結果表明,病程相關蛋白僅是山藥抵抗炭疽菌侵染的部分因子,它們與其他諸多因子共同起作用,且在不同程度或種類的抗病品種(系)中他們的生物學功能也有差異[14]。同樣,6個細胞壁修飾蛋白中,阿拉伯半乳聚糖蛋白(AGP19)和鈣調素結合蛋白(SHA1)在炭疽菌侵染24 h上調表達,而導致細胞壁松弛的擴展蛋白(EXLB1)和細胞壁降解相關木葡聚糖內糖基轉移酶(XTH22)下調表達,可阻止細胞壁的松弛,增強蘇蕷8號對炭疽菌的免疫功能。
正常條件下活性氧(ROS)的產生與清除處于動態平衡,當植物在遭受逆境或脅迫時,植物體內ROS平衡會被打破,誘導體內積累ROS使植物發生抗病反應,過多的ROS積累會使膜脂過氧化而使膜系統受損,對植物產生傷害[15]。為了維持平衡,一方面植物通過抗氧化酶系統(SOD、POD、CAT等)將活性氧分解;另一方面依靠抗氧化次生代謝產物,直接將ROS清除,例如抗壞血酸、類胡蘿卜素、谷胱甘肽、多酚和類黃酮等[16]。蘇蕷8號受炭疽菌侵染后,過氧化氫酶(CAT2)和線粒體超氧物歧化酶(MSD1)上調表達,MSD1可以將O2-歧化為 H2O2,CAT2通過直接將H2O2分解生成無毒害H2O和O2[17]。
CYP450s是一種廣譜性生物催化酶,參與多種代謝反應,如脂肪酸代謝、植物激素的生物合成與降解、次生代謝產物的合成[18]。CYP94A1-like參與葉片角質的合成[19]、CYP51編碼甾醇14α-去甲基化酶[20],與甾醇C-24甲基轉移酶(BRADI_1g76020v3)共同參與生物甾醇的合成[21],葉片角質和作為質膜組分的生物甾醇合成相關基因在蘇蕷8號的上調表達能有效防止病原物侵染植物組織。CYP78A家族成員(CYP78A3、CYP78A5-like)可能成為一類應用于作物改良的基因,參與種子和器官的發育[22],它們上調表達是否與抗病有關,尚未見報道。防御素(SD2)、幾丁質結合凝集素(CBL1)和木菠蘿凝集素(JRL5)基因在蘇蕷8號的上調表達,可與真菌細胞表面的不同單糖,多糖支鏈或異源多糖結合,從而干擾真菌細胞壁的合成,抑制其生長[23-24]。
植物泛素化系統是在一系列酶的催化作用下,泛素共價結合靶標蛋白對其進行泛素化修飾,其泛素化靶標蛋白可介導抗病信號,甚至一些病原物效應蛋白也在其宿主細胞中起泛素連接酶的作用,并以此來干擾植物抗病反應[25]。在水稻中,E2受稻瘟病菌和植物激素誘導表達,在激素信號傳導中發揮重要的作用[26]。同樣,受稻瘟菌侵染時,編碼RING型的E3連接OsBBI1,過表達后植株的細胞壁明顯增厚,而且感染部位的H2O2和酚類化合物大量積累,即OsBBI1參與由細胞壁介導的水稻抗病機制[27]。蘇蕷8號受炭疽菌后,泛素化連接酶(UBE2、UBE3)上調表達,可能有利于增強蘇蕷8號的抗性。
3.3 轉錄因子調控山藥炭疽病抗性
轉錄因子可作為防御基因表達中的正或負調節因子,通過其保守結構域與靶基因啟動子上的特異結構特異性結合,來實現抗逆調控功能[28]。WRKY48參與擬南芥丁香假單胞菌侵染后的防衛反應,過表達WRKY48,丁香假單胞菌的生長的敏感性增強與防御相關PR基因表達減少有關,WRKY48負調節PR基因的表達[29]。棉花GhWRKY48基因對棉花枯萎病、黃萎病、水楊酸、茉莉酸均有一定的響應,GhWRKY48基因正向調節棉花抗枯萎病反應,負向調節棉花對黃萎病的抗性[30-31],WRKY75正向調節茉莉酸鹽介導的植物對壞死型真菌病原菌的防御[32]。本試驗中WRKY48上調表達,WRKY75下調表達,可能分別正向和負向調控山藥對炭疽病的響應。表明,WRKY家族同一個基因不僅在不同的作物調控功能不盡相同,即使在同一作物的調控功能也不同。棉花中MYB108通過與鈣離子和鈣調素樣蛋白GhCML11協同作用正向調節棉花黃萎病響應[33]。TIFY10C是水稻中一個茉莉酸信號途徑的抑制子,通過與OsCOI1形成復合體或與其他蛋白質形成異源二聚體抑制茉莉酸信號途徑,JA處理導致TIFY10C降解,從而對白葉枯病信號作出響應[34]。蘇蕷8號下調表達MYB108和TIFY10C,可能負向調節山藥對炭疽病的響應。GATA12、HOX11、RSS3、ZFP7、DREB類轉錄因子在植物生長發育,激素信號途徑、次生代謝產物合成、抗逆反應等方面發揮重要作用[35-40],在植物病害調控方面未見相關報道。
3.4 植物激素信號轉導與山藥炭疽病抗性
生長素信號傳導過程涉及了2類轉錄因子,一類是生長素響應因子AFR家族,另一類是Aux/IAA轉錄因子家族,AFR能和生長素早期誘導基因如 Aux/IAA,SAUR,GH3等家族的啟動子響應元件特異結合形成二聚體,激活或者抑制基因表達。當生長素濃度較高時,生長素與泛素連接酶復合體SCFTIR1結合后,會促使Aux/IAA抑制因子的泛素化與降解,產生眾多ARF蛋白并激活生長素信號途徑[41]。病原菌侵染導致生長素水平失衡,進而改變其信號途徑相關基因的表達。水稻GH3-2過量表達后能增強水稻對白葉枯病菌、細菌性條斑病菌和稻瘟病的抗性[42]。擬南芥生長素信號缺失突變體axr2(Auxin resistant 2) 對丁香假單胞桿菌的抗性增強,表明生長素信號途徑不利于擬南芥對丁香假單胞桿菌的免疫反應[43]。本研究中,LAX4、IAA4、IAA21、SAUR36、SAUR71在炭疽菌侵染24 h均上調表達,且LAX4和IAA21在抗病品種表達量顯著高于感病品種,推測生長素信號途徑有利于山藥對炭疽病的免疫反應。
茉莉酸信號途徑中茉莉酸合成關鍵酶基因OPR1和脫落酸合成關鍵酶基因NCED上調表達,有利于增強蘇蕷8號炭疽病抗性。JA在馬鈴薯晚疫病菌侵染期間,會識別Pep-13致病因子,激活茉莉酸信號途徑,誘導ROS的產生、PR基因的表達及細胞超敏死亡,增強對馬鈴薯晚疫病的抗性[44]。ABA可以破壞煙草中由水楊酸介導的對煙草花葉病毒(TMV)和葉斑病的抵抗[45],亦可控制植物氣孔關閉,促進胼胝質沉積阻止病菌侵染,正向調控植物的抗病性[46]。油桐在尖孢鐮孢菌侵染后,VmAP2/ERF036和其同源基因VfAP2/ERF036表現出相反的表達模式,VmAP2/ERF036的表達受到抑制,表明二者的抗病調節模式不同[47]。ERF036在蘇蕷8號下調表達,可能負向調控山藥炭疽菌的侵染。GASA(Gibberellic Acid-stimulated Arabidopsis)蛋白是一類CRP蛋白,大多數成員受赤霉素調控。目前只在馬鈴薯、辣椒和法國菜豆中發現GASA蛋白參與植物防御反應[48-50]。但已有證據表明,GASA蛋白可通過促進水楊酸SA合成基因的表達和植物體內SA的水平來抵抗非生物脅迫[51]。本研究中GASA14在炭疽菌侵染24 h上調表達,GASA2下調表達,它們如何參與蘇蕷8號抗病性還需要進一步研究。
猜你喜歡
語數外學習·高中版中旬(2023年2期)2023-05-10 13:26:53
語數外學習·高中版中旬(2022年5期)2022-07-13 20:47:51
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中學生百科·大語文(2017年10期)2017-11-04 06:56:38
中國衛生(2016年3期)2016-11-12 13:23:26
公民與法治(2016年22期)2016-05-17 04:20:13
中國衛生(2014年12期)2014-11-12 13:12:52
癌變·畸變·突變(2014年6期)2014-02-27 06:15:03
