奶山羊ATGL基因啟動子鑒定與轉錄活性分析
2022-05-12 10:58:26閆志浩賈萬里許蒙蒙張景鋒徐秋良韓浩園
華北農學報 2022年2期
李 君,閆志浩,賈萬里,許蒙蒙,張景鋒,徐秋良,韓浩園,權 凱
(1.河南牧業經濟學院 動物科技學院,河南 鄭州 450046;2.河南省反芻動物營養與飼料資源開發國際聯合實驗室,河南 鄭州 450046;3.西峽縣農業農村局,河南 西峽 474550)
脂肪甘油三酯水解酶(Adipose triglyceride lipase,ATGL)是脂肪水解過程中的主要限速酶,它可以特異性地水解甘油三酯為甘油二酯和游離脂肪酸,是調控細胞中甘油三酯和脂肪酸之間平衡的關鍵因子[1]。ATGL在成熟的棕色脂肪組織中表達量最高,主要存在于細胞的胞質內,少量分布于脂滴中。除脂肪組織外,其在肝臟、心臟、腸胃、大腦和腎臟中也有少量表達[2]。在COS-7細胞中過表達ATGL,檢測到培養基中游離脂肪酸含量增加,細胞內甘油三酯儲存減少,表明ATGL在甘油三酯水解過程中發揮重要作用[3]。在妊娠和泌乳這種特殊的生理階段,乳腺具有強大的短期合成和分解脂質的能力,是機體最活躍的代謝器官。研究發現,ATGL在奶山羊的各組織中均有表達,且在脂肪組織、肺臟和乳腺組織中表達量高于其他組織;同時還發現,ATGL在泌乳的乳腺組織中表達量高于干奶期乳腺組織,且干擾ATGL顯著增加乳腺上皮細胞中甘油三酯含量[4]。因此推測,ATGL在奶山羊乳腺泌乳過程中發揮重要的生物學功能。
ATGL作為脂肪第一步水解過程中的主要限速酶,控制著整個水解過程的進展速率。ATGL的表達量及活性高低對甘油三酯和脂肪酸含量的影響具有關鍵作用。啟動子是控制基因轉錄活性的重要元件,研究表明,ATGL的轉錄活性受到營養和激素水平的調控,饑餓會激活ATGL轉錄、喂食后則降低其轉錄水平[5]。多個轉錄因子可調控ATGL轉錄,Roy等[6]研究發現,過氧化物酶體增殖物激活受體(Peroxisome proliferators-activated receptors,PPARG)與特異性蛋白1(Specificity protein 1,Sp1)相互作用共同調控ATGL的轉錄。Kaltenecker等[7]在小鼠脂肪細胞中研究發現,生長激素通過信號轉導與轉錄活化蛋白5(Signal transducers and activators of transcription 5,STAT5)促進ATGL的轉錄,從而促進脂解。P53上調ATGL轉錄活性并且在脂肪細胞中促進脂解[8]。研究還表明,ATGL基因的轉錄受到干擾素調節因子4(Interferon regulatory factor 4,IRF4)和叉頭蛋白O1(Forkhead box protein O1,FOXO1)轉錄因子的調節,生物信息學分析發現,ATGL的啟動子區域含有至少2個FOXO1結合位點[9-10]。因此,加強對ATGL基因啟動子的深入研究,將有助于掌控ATGL的轉錄表達。山羊奶脂肪酸含量豐富,乳脂滴小,利于消化吸收。
為了進一步提高羊乳品質,本研究在對奶山羊ATGL基因功能研究的基礎上,通過對其啟動子進行克隆與轉錄活性分析,對ATGL轉錄調控機制進行分析,旨在為進一步研究該基因的表達調控機制以及改善羊乳品質提供基礎資料。
1 材料和方法
1.1 試驗材料與試劑
血液基因組DNA提取試劑盒和DNA回收試劑盒購自北京艾德萊生物科技有限公司;LATaqDNA聚合酶、克隆載體pMD19-T、限制性內切酶BglⅡ和MluⅠ均購自TaKaRa公司;DNA Marker、感受態細胞TOP10和高純度質粒小提中量試劑盒等購自北京天根生化科技有限公司;雙熒光素酶報告系統購自Promega公司;X-tremeGENE HP DNA轉染試劑購自羅氏公司;DMEM/F-12培養基、表皮生長因子、氫化可的松、胰島素、催乳素、胎牛血清等購自Invitrogen公司。奶山羊血液和奶山羊乳腺上皮細胞由河南牧業經濟學院羊生物育種實驗室提供。
1.2 ATGL基因啟動子的克隆
利用血液基因組DNA提取試劑盒提取奶山羊基因組DNA。參照GenBank中牛(Bostaurus)的基因組序列,利用Primer Premier 5.0設計合成山羊(Caprahircus)ATGL基因啟動子克隆引物APF和APR,啟動子缺失片段引物APS1、APS2、APS3、APS4、APS5、APA(表1)。以奶山羊基因組DNA為模板,PCR擴增ATGL基因啟動子序列,反應體系:DNA模板(100 ng/μL)0.5 μL,上、下游引物(10 μmol/L)各1 μL,2×GC Buffer Ⅱ 10 μL,dNTP Mix(100 μmol/L)2 μL,LATaq酶0.2 μL,ddH2O補齊至20 μL;反應程序:95 ℃預變性4 min;95 ℃變性 30 s,56 ℃退火30 s,72 ℃延伸 2 min,33 個循環;72 ℃ 再次延伸10 min,12 ℃保存。利用1%瓊脂糖凝膠電泳對PCR產物進行檢測,將預期目的條帶進行凝膠回收。將回收產物與pMD19-T于16 ℃連接4 h,然后轉化入感受態細胞TOP10,挑取單克隆菌落于LB液體培養基內培養12~16 h,擴繁后用高純度質粒提取試劑盒提取質粒,經BglⅡ和MluⅠ雙酶切鑒定后篩選陽性克隆,送至上海生工生物工程有限公司測序。
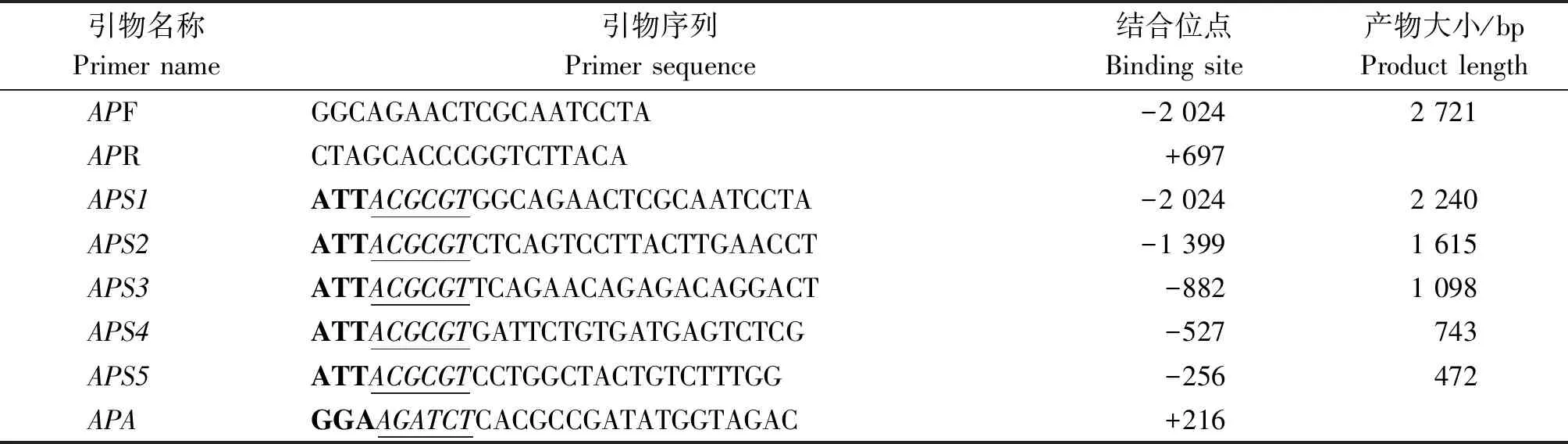
表1 奶山羊ATGL基因啟動子克隆引物序列Tab.1 The sequences of primers used for cloning ATGL gene promoter of dairy goat
1.3 ATGL基因啟動子的生物信息學分析
應用Promoter 2.0預測ATGL基因的啟動子核心區域;ATGL基因啟動子CpG島采用CpG Islands(2004版,http://www.ebi.ac.uk/emboss/cpgplot/)在線軟件進行分析;ATGL基因啟動子上潛在的轉錄因子結合位點應用Gene Regulation(2015版,http://www.gene regulation.coml)和JASPAR(2020版,http://jaspar.genereg.net/)數據庫進行預測,綜合比較預測分值較高的結合位點,以確保其具有較高的特異性[11]。
1.4 ATGL啟動子報告基因載體構建
根據設計的啟動子缺失片段引物(表1),以含有ATGL啟動子的陽性質粒為模板進行PCR擴增,得到一系列帶有BglⅡ和MluⅠ酶切位點的啟動子缺失片段,分別將其與pGL3-Basic載體(Promega)經雙酶切后進行連接,最后得到含有ATGL啟動子不同缺失片段的報告基因重組質粒,對酶切鑒定正確的質粒進一步測序以鑒定其準確性,用于后續試驗。
1.5 細胞培養與轉染
奶山羊乳腺上皮細胞培養采用含10% FBS、100 U/mL青霉素、100 μg/mL鏈霉素、10 ng/mL表皮生長因子、5 μg/mL氫化可的松和5 μg/mL胰島素的DMEM/F12細胞培養基,于37 ℃,5% CO2培養箱中培養24 h,更換新鮮培養基,倒置顯微鏡下觀察細胞匯合度達90%以上時,即可進行傳代培養。
細胞于轉染前一天鋪48孔板,待細胞密度達到70%~80%時,參照轉染試劑操作說明進行DNA轉染。48孔板每孔轉染復合物體系為:X-tremeGENE HP 1 μL,質粒 0.3 μg(pGL3-ATGL:pRL-TK =25∶1),DMEM/F12補齊至30 μL。每個處理重復3次。
將羅格列酮和T0901317溶解于DMSO中,分別配制成10 mol/L的處理液。DNA轉染細胞12 h后,將配制好的羅格列酮或T0901317加入細胞中至最終的試驗濃度分別為50,1 μmol/L,同時對照組用相同濃度的DMSO進行處理。培養36 h后收集細胞,用于后續雙熒光素酶活性檢測。
1.6 雙熒光素酶活性檢測
乳腺上皮細胞轉染48 h后,收集細胞,按照雙熒光素酶報告基因檢測試劑盒操作說明測定其活性。先棄去培養基,用PBS清洗細胞2次以去除殘留培養基,之后加入細胞裂解液于搖床上孵育30 min以充分裂解細胞,最后于多功能酶標儀上檢測螢火蟲熒光素酶活性(F值)與海腎熒光素酶活性(R值),以螢火蟲與海腎的比值代表ATGL啟動子的相對活性。每孔進行3次測定,取平均值。
1.7 數據分析
對于啟動子活性檢測結果,利用數據分析軟件SPSS 22.0進行統計學分析,每個處理設置3個生物學重復,采用t檢驗和單因素方差分析進行顯著性檢驗,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結果與分析
2.1 奶山羊ATGL啟動子的克隆與序列鑒定
利用引物APF和APR進行PCR擴增,成功得到約2 700 bp的ATGL基因5′側翼序列(圖1)。連接克隆載體,將鑒定為陽性的重組質粒進行測序,結果顯示,成功克隆得到奶山羊ATGL基因5′側翼序列2 721 bp。

圖1 ATGL基因啟動子的PCR擴增產物Fig.1 The PCR amplification products of ATGL gene promoter
2.2 ATGL啟動子的生物信息學分析
經分析發現,擴增得到的ATGL基因5′側翼序列,轉錄起始位點(+1)上游2 024 bp及下游697 bp,包含外顯子1和部分內含子1(圖2-A)。CpG島分析結果顯示,其序列中存在1個CpG島,位置為-253—+1位,且ATGL啟動子具有典型的TATA-box。經在線軟件預測發現,ATGL啟動子上存在多個重要的調控元件,包括FOXO1、FOXO3、SREBF1、PPARA、PPARG、C/EBPα和E2F1等轉錄因子結合位點(圖 2-B)。

A.ATGL基因5′側翼序列圖示;B.ATGL啟動子潛在的轉錄因子結合位點分析(+1為轉錄起始位點)。A.Schematic representation of ATGL 5′flanking region;B.Analysis of the putative transcription factor binding sites of the ATGL promoter(+1 is the first base at transcription start site).
2.3 ATGL啟動子缺失片段載體構建
以ATGL基因5′側翼序列全長為模板,利用一系列缺失引物進行PCR擴增,得到5個不同長度的啟動子截短片段(圖3-A)。通過連接pGL3-Basic報告基因載體,得到啟動子缺失片段報告基因重組質粒,通過酶切鑒定(圖3-B)和測序,結果證實重組質粒構建成功。
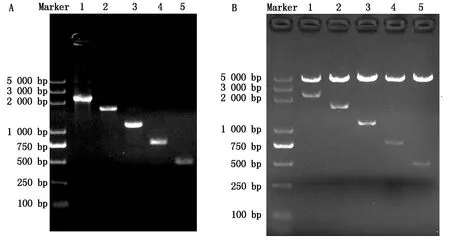
A.不同缺失片段PCR擴增產物:1.-2 024—+216位;2.-1 399—+216位;3.-882—+216位;4.-527—+216位;5.-256—+216位;B.不同缺失片段重組質粒酶切鑒定:1.pGL-APS1(-2 024—+216位);2.pGL-APS2(-1 399—+216 位);3.pGL-APS3(-882—+216位);4.pGL-APS4(-527—+216位);5.pGL-APS5(-256—+216位)。
2.4 ATGL基因啟動子核心區域確定
將ATGL缺失片段報告基因載體和內參質粒(海腎熒光素酶載體)共轉染奶山羊乳腺上皮細胞,48 h后收集細胞檢測相對熒光素酶活性。結果表明,ATGL啟動子區域由-2 024位缺失到-882位時,啟動子活性無顯著變化(P>0.05);由-882位缺失到-527位時,其活性顯著降低(P<0.05);由-527位缺失到-256位時,啟動子活性又顯著上升(P<0.05)(圖4)。說明啟動子區域-527—-256位可能存在抑制元件,推測轉錄起始位點上游-256 —+1位為ATGL基因啟動子轉錄核心區域。

pGL3-Basic.熒光素酶報告基因空載體;LUC.不同缺失片段的熒光素酶報告基因載體;不同字母表示差異顯著(P<0.05)。圖5—6同。pGL3-Basic.Luciferase reporter gene empty vector;LUC.Luciferase reporter gene vector with various deletion fragments;Different letters indicate significant differences(P<0.05).The same as Fig.5—6.
2.5 羅格列酮對ATGL啟動子活性的影響
對乳腺上皮細胞轉染pGL-APS1重組質粒,用PPARG特異性激動劑羅格列酮(ROSI)處理細胞,結果發現,羅格列酮顯著上調ATGL啟動子活性(P<0.05)(圖5-A)。接著將活性變化較為顯著的pGL-APS3、pGL-APS4和pGL-APS5重組質粒轉染奶山羊乳腺上皮細胞,并用羅格列酮處理,結果發現,APS3和APS4均能響應羅格列酮信號(圖5-B),表明PPARG調控奶山羊ATGL啟動子活性,且作用區域位于-527—-256位。

A.羅格列酮對ATGL啟動子全長活性的影響;B.羅格列酮對ATGL啟動子不同片段的影響。A.Effect of ROSI on the activity of ATGL promoter in full length;B.Effect of ROSI on the activity of various deletion fragments of ATGL promoter.
2.6 T0901317對ATGL啟動子活性的影響
以同樣的方法在乳腺上皮細胞中轉染pGL-APS1重組質粒,用SREBP1激動劑T0901317處理細胞發現,T0901317顯著上調ATGL啟動子活性(P<0.05)(圖6-A);而在不同缺失片段中,T0901317僅對APS3有顯著上調作用(P<0.05),對APS4和APS5沒有顯著影響(P>0.05)(圖6-B),表明SREBP1參與調控奶山羊ATGL基因轉錄,且其作用區域位于-882—-527位。生物信息學分析發現,該片段中-710—-703位和-600—-592位存在預測的SREBP1結合位點,T0901317可能通過這2個位點發揮調控作用。

A.T0901317對ATGL啟動子全長活性的影響;B.T0901317對ATGL啟動子不同片段的影響。A.Effect of T0901317 on the activity of ATGL promoter in full length;B.Effect of T0901317 on the activity of various deletion fragments of ATGL promoter.
3 結論與討論
ATGL作為甘油三酯水解的限速酶,可以催化動物脂肪組織和非脂肪組織的甘油三酯水解[12],前期的研究發現,ATGL在奶山羊乳腺組織中高表達,且在乳腺上皮細胞中發揮重要作用,ATGL影響細胞中甘油三酯含量和游離脂肪酸含量,同時影響脂肪酸代謝相關基因表達水平[4]。基于前期的這些研究成果,對奶山羊ATGL基因啟動子的結構與轉錄活性進行分析,有助于闡明ATGL基因在乳腺細胞中的轉錄調控機制,對于研究羊奶中乳脂和乳脂肪酸的調控作用具有重要意義。
不同的組織和細胞含有相同的DNA序列,而組織和細胞功能的多樣性則是由基因的差異表達實現的,因此,研究基因的轉錄調控對理解生命活動具有重要意義。轉錄調控主要是通過轉錄因子結合靶基因啟動子上特定DNA序列來實現,啟動子上存在多種調控元件,這些元件的數量、種類以及它們之間的距離與順序都有可能影響基因的轉錄,它們通過激活或抑制基因轉錄,對基因的表達差異發揮作用[13]。ATGL基因在脂肪細胞中受到多種轉錄因子調控,其5′調控區含有E-box、TATA box、CAAT box等結合位點[14],多種轉錄因子可以識別基因5′調控區的特異位點并與之作用調控ATGL基因的表達[15]。本研究以河南奶山羊基因組DNA為模板,擴增出ATGL基因5′側翼序列2 721 bp,參考其他物種的ATGL啟動子序列和生物信息學分析發現,轉錄起始位點上游序列包含典型的啟動子元件(TATA-box),這些結果與在豬上的研究結果一致[16]。在線軟件預測發現,奶山羊ATGL啟動子中還含有FOXO1、FOXO3、SREBF1、PPAR和CEBPα等轉錄因子結合位點,這些結合位點和豬上ATGL啟動子預測結果相一致[17]。通過缺失分析發現,從-882位缺失到-527位時啟動子活性顯著下降,表明此區域含有正調控元件,而從-527位缺失到-256位時,啟動子活性反而上調,表明此區域含有抑制其活性的元件。經過分析確定其核心區域在-256—+1位,這與孫健[16]在草魚上和王小宇[17]在豬上的研究報道結果基本一致,但是二者報道的ATGL啟動子活性區域更小。
PPARG已被證實在奶牛和奶山羊的乳腺脂肪酸調控網絡中發揮重要作用,其影響脂肪酸的從頭合成、甘油三酯的合成和分泌相關基因的表達[18-20],PPARG通過結合在PPAR 反應元件上對下游基因表達進行調控[21]。Shi等[22]研究發現,在奶山羊乳腺上皮細胞中,羅格列酮和PPARG過表達影響奶山羊ATGL基因mRNA水平,在多個物種的ATGL啟動子上均存在PPARγ結合位點[23]。有研究發現,在脂肪細胞中PPARG抑制ATGL基因的轉錄,但是在脂肪細胞分化后期,PPARG促進ATGL基因的轉錄,這種促進作用是通過與轉錄因子Sp1 的共同作用完成,說明PPARG對ATGL的調控具有時空特異性[6]。本試驗中,通過在線軟件預測到在ATGL啟動子的-282—-267位存在潛在的PPARG結合位點,通過PPARG特異性激動劑處理細胞發現,羅格列酮在截短片段的-527—-256位發揮作用,表明PPARG調控ATGL基因轉錄,這與前人的研究結果一致。
SREBP1是調控脂肪合成的主要轉錄因子,SREBP1核蛋白在運輸進入細胞核后與靶基因啟動子上的位點結合從而激活靶基因的轉錄[24-25]。研究證實,LXRα可直接結合在SREBP1的啟動子元件上,對其進行轉錄調控[26]。而LXRα的人工合成激動劑T0901317又被廣泛地作為SREBP1的激動劑應用于SREBP1的功能研究中[27]。Xu等[28]研究發現,過表達SREBP1影響ATGL基因mRNA表達。本研究通過T0901317處理細胞,發現其顯著上調ATGL啟動子活性,且作用區域位于-882—-527位;生物信息學分析發現,奶山羊ATGL啟動子-710—-703位和-600—-592位存在預測的SREBP1結合位點。表明SREBP1可能是ATGL基因的轉錄因子,促進ATGL基因轉錄,但是關于SREBP1對ATGL啟動子的調控作用及其作用位點目前還未見報道,在奶山羊上還需進一步的試驗驗證。
本研究成功克隆奶山羊ATGL啟動子序列,利用生物信息學分析了ATGL啟動子序列中的調控元件結合位點,構建了不同片段長度的ATGL啟動子報告基因載體,啟動子活性分析表明其核心區域位于-256—+1位,且PPARG和SREBP1能夠促進ATGL基因的啟動子活性,且作用區域分別位于-527—-256位和-882—-527位。
