海草土壤種子庫:特征、影響因素、研究方法及其在受損海草場恢復(fù)中的作用
2022-05-16 07:53:24邱廣龍權(quán)佳惠蘇治南范航清招禮軍
應(yīng)用海洋學(xué)學(xué)報 2022年2期
關(guān)鍵詞:植物
邱廣龍,權(quán)佳惠,蘇治南,范航清,招禮軍*
(1.廣西科學(xué)院廣西紅樹林研究中心、廣西紅樹林保護(hù)與利用重點實驗室,廣西 北海 536000;2.廣西大學(xué)林學(xué)院,廣西 南寧 530004)
土壤種子庫是指土壤中和凋落物中具有活力的全部種子[1]。豐富的種子庫多樣性在退化生境中為重建多種植被提供了可能性,成為植被周期變化的關(guān)鍵[2]。它是種群更新與拓展的重要基礎(chǔ),作為繁殖體的儲備庫和潛在的植被,既反映了群落的歷史,也預(yù)示著種群的自我恢復(fù)能力,并能減小種群滅絕的幾率[3-4]。基于其重要的生態(tài)學(xué)和遺傳學(xué)意義,土壤種子庫研究日益受到重視。
海草是生長在淺海地區(qū)的單子葉植物。全球海草有72種,除了極少數(shù)種類,絕大部分海草均為水媒傳粉植物,它們在水中完成其生活史。作為克隆植物,海草既能進(jìn)行無性繁殖,也可進(jìn)行有性繁殖,產(chǎn)生種子或胎生苗[5]。當(dāng)具有活力的海草種子進(jìn)入土壤(又稱“沉積物”,下文統(tǒng)稱“土壤”)中,形成了土壤種子庫。本文對海草土壤種子庫的特征、采樣方法、影響因素和生態(tài)學(xué)、遺傳學(xué)意義等作了系統(tǒng)的綜述,為海草的保護(hù)與恢復(fù)提供一些有益的思路。
1 海草土壤種子庫的類型和特征
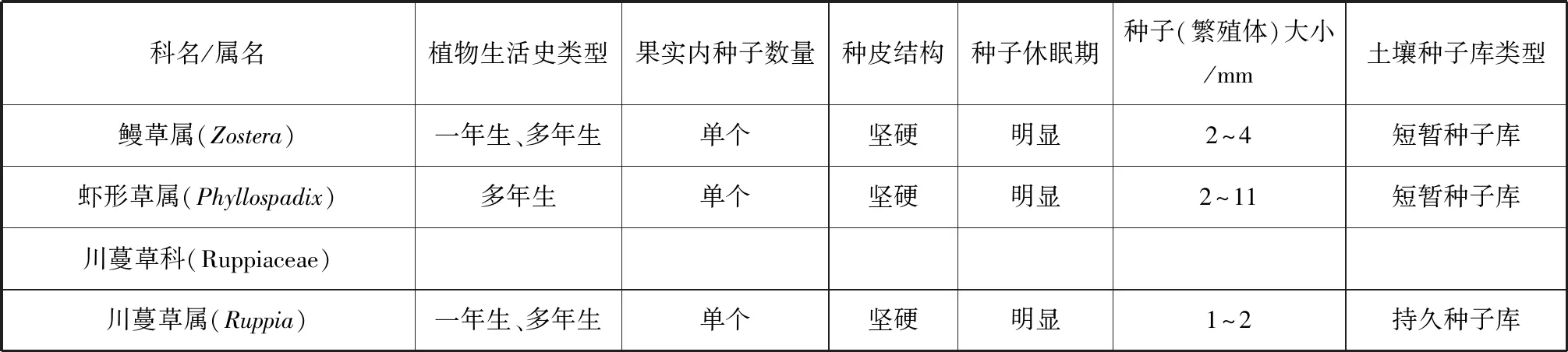
在全球11屬海草中,有6個屬的海草可形成種子庫(表1)。在不能形成種子庫的海草種類中,一部分海草的種子在脫離母體后可快速發(fā)芽生長,例如泰來草屬(Thalassia),其種子在脫離后數(shù)天內(nèi)即可發(fā)芽,若被沉積物覆蓋,則一周內(nèi)失去活力;另外一些海草可形成胎生苗,例如全楔草屬(Thalassodendron)和根枝草屬(Amphibolis)[6-8]。
續(xù)表
具有休眠特性的海草種子形成了海草土壤種子庫。海草種子的休眠特性使物種得以擴(kuò)散并形成種子庫,并在不同的空間和時間尺度再生長。根據(jù)種子休眠時長,可把海草土壤種子庫分為持久種子庫(persistent seed bank,種子壽命≥1 a)或短暫種子庫(transient seed bank,種子壽命<1 a)[7]。持久種子庫的種子壽命較長,例如絲狀針葉草(Syringodiumfiliforme),其種子在沉積物中保持活力可超過4 a[9]。短暫種子庫的種子壽命相對較短,例如鰻草屬(Zostera)和蝦形草屬(Phyllospadix),它們的種子休眠期通常不到1 a。種子的休眠特性是形成土壤種子庫的基礎(chǔ),然而,與陸地植物相比,海草種子的休眠期普遍較短,目前所報道最長的種子休眠期,也僅為數(shù)年,而有些陸地植物種子的休眠期可達(dá)數(shù)百年甚至數(shù)千年[9-10]。
海草的土壤種子庫具有巨大的種間差異和空間差異。不同海草種子庫密度數(shù)量級為100~105,例如小絲粉草(Cymodoceanodosa)、諾氏鰻草(Zosteranoltii)、魔蝎鰻草(Zosteracapricorni)的土壤種子庫密度分別為0~220、367~488、177粒/m2[11-13],而三脈喜鹽草(Halophilatricostata)、單脈二藥草(Halo-duleuninervis)、毛葉喜鹽草(Halophiladecipiens)的土壤種子庫密度可分別高達(dá)70 000、80 000、176 880粒/m2[14-16]。即使是同一種海草,不同種群之間差別也較大。例如馬來西亞登嘉樓州甘馬挽的貝克喜鹽草(Halophilabeccarii)種群土壤種子庫密度僅156粒/m2[17],而我國華南地區(qū)貝克喜鹽草土壤種子庫密度為2 105~5 749粒/m2[18-19],局部區(qū)域可高達(dá)24 685粒/m2[20]。
土壤種子庫的空間分布包括種子的水平分布和垂直分布。在水平分布上,不同空間尺度(cm~km)的海草土壤種子庫密度均表現(xiàn)出巨大的異質(zhì)性[12,21-22]。而在垂直分布上,種子庫密度常具有自上而下遞減的特點[19,23]。這一特性使種子庫在空間上具有立體結(jié)構(gòu),影響著種子庫種子的留存和萌發(fā),進(jìn)而影響原有植被的恢復(fù)與重建。垂直分布的特性使小部分種子處于下層土壤中,發(fā)芽率一般較低,但由于下層水熱環(huán)境相對穩(wěn)定,種子可以存活較長時間,形成了植物種群的天然基因庫,對物種保護(hù)及維護(hù)生物多樣性具有重要意義[24-25]。此外,對同一種海草而言,種子越重,被埋藏的深度通常越深,但也意味這些重的種子有更多的代謝儲備,因此種子有更長的壽命,由此發(fā)育的幼苗有更高的生物量[26]。
海草土壤種子庫同時具有高度的時間動態(tài)性[22,27]。Harrison(1993)對荷蘭西南部一年生鰻草的研究表明,不同季節(jié)土壤種子庫密度差異顯著,約有80%埋藏的種子在次年2月前消失[22]。對貝克喜鹽草的研究也表現(xiàn)出了土壤種子庫的時間動態(tài)性[20]。
2 影響海草土壤種子庫的因素
2.1 植被/植物特性
長期以來,人們認(rèn)為土壤種子庫與地上植被并沒有必然的聯(lián)系,陸地植物和海草皆如此[7,24,31,33]。然而,隨著研究的深入,人們發(fā)現(xiàn)地上植被與土壤種子庫的關(guān)系比之前發(fā)現(xiàn)的更復(fù)雜,因為兩者之間的關(guān)系受植被的演替階段、植物生物學(xué)、生理學(xué)特征等的影響。一般認(rèn)為在演替早期或經(jīng)常受干擾的生境中,其種子細(xì)小且具休眠性,易于被埋藏在土壤庫中。因此,在土壤種子庫中,常見的是次生演替早期種類的種子[33]。但隨著植被的演替,形態(tài)較大、密度較小、缺乏休眠性、適應(yīng)于穩(wěn)定環(huán)境的種子逐漸出現(xiàn)[33-34]。在多年生植物占優(yōu)勢的生境,成年植物的密度與其種子庫密度無關(guān),而在一年生或短命植物占優(yōu)勢的生境中則有較強(qiáng)的相關(guān)性[34]。例如,韋梅球等(2017)對短命海草貝克喜鹽草的研究表明,其土壤種子庫密度與其植被雌花密度、直立莖密度存在極顯著正相關(guān)關(guān)系[19]。
種子庫的規(guī)模與種子的形態(tài)大小有關(guān),通常種子數(shù)量多且個體小的植物具有較大的土壤種子庫,這一規(guī)律與陸地植物相似[7,23,33,35]。此外,海草植物的生活史類型也對土壤種子庫動態(tài)產(chǎn)生影響。一般而言,一年生海草種群的種子庫比多年生海草種群更具規(guī)模,且時間波動性更大[5,17,36]。
繁殖體擴(kuò)散能力較弱的海草(例如喜鹽草屬和二藥草屬)更有可能形成持久的種子庫,并迅速成為定殖草場內(nèi)的定居者。相反,果實較大且有浮力的屬(例如泰來草屬和波喜蕩草屬)有更強(qiáng)的擴(kuò)散能力,較少在原草場進(jìn)行補(bǔ)充更新[15]。
2.2 動物因素
一些海洋動物,例如沙蠶、蟹類、儒艮(Dugongdugon)等的取食行為或擾動可能對種子庫的動態(tài)造成較大的影響。動物的取食行為會減少進(jìn)入到土壤種子庫中的種子,還可造成種子的水平移動,或促進(jìn)種子的垂直移動,影響種子的埋藏[37-39]。據(jù)報道,藍(lán)蟹(Callinectessapidus)最高可取食65%的鰻草(Zosteramarina)種子[38]。
動物既是海草種子的捕食者,同時也是某些海草種子散播的重要媒介[24]。據(jù)報道,在儒艮糞便中曾發(fā)現(xiàn)大量具有活力的海草種子,例如卵葉喜鹽草(Halophilaovalis)、毛葉喜鹽草、單脈二藥草、牟氏鰻草(Zosteramuelleri)等的種子[23],由此可見海洋動物的行為能影響土壤種子庫的空間分布。
一些底內(nèi)無脊椎動物(例如環(huán)節(jié)動物)的生物擾動,也會促進(jìn)海草種子在土壤中的移動,從而影響種子的埋藏。然而,埋藏的深度對某些海草種子的存活具有決定性的影響。例如,適度的埋藏有利于鰻草種子的萌發(fā),但埋藏過深則會影響種子的活力,從而又降低了萌發(fā)率[39]。
2.3 環(huán)境因素
環(huán)境條件可通過影響海草植物的有性繁殖產(chǎn)出,從而影響到土壤種子庫的規(guī)模。在很多情況下,影響種子產(chǎn)量的環(huán)境因素可能比直接影響土壤種子庫的環(huán)境因素更能決定種子庫的分布與規(guī)模[7,21]。
大規(guī)模的干擾例如颶風(fēng)、風(fēng)暴潮等可顯著改變沉積物的空間位置,從而改變了海草土壤種子庫在空間上的水平分布和垂直分布。對潮下帶海草毛葉喜鹽草的一項研究表明,颶風(fēng)造成了沉積物和種子庫的空間再分配,導(dǎo)致土壤種子庫的整體移動[40]。
盡管大部分的海草種子密度比海水大,從母體中釋放后常會沉降到母體植物附近的沉積物中,但是一些海草的佛焰苞、果實或花序在海水中具有較好的浮力[5],從而加強(qiáng)了這些海草的種子擴(kuò)散,使土壤種子庫在空間上分布范圍更廣。因此,區(qū)域或局部的水文條件會對土壤種子庫的空間分布產(chǎn)生重要的影響。
微生境的差異(例如洼地、土壤裂縫)是造成土壤種子庫空間異質(zhì)性的重要原因。例如,對單脈二藥草的研究表明,在被定向水流或儒艮取食行為去除了植被的洼地,比周邊有更高的土壤種子庫密度[21]。
3 海草土壤種子庫的研究方法
3.1 采樣方法
海草土壤種子庫可通過一定容積的柱狀采樣器進(jìn)行樣品采集,采樣方法與海草生物量的采集基本相同[31,41]。考慮到較深的沉積物中幾乎不存在有活力的種子,采集沉積物時不必過深,通常以10~20 cm深度即可。
土壤種子庫取樣量的大小無統(tǒng)一標(biāo)準(zhǔn),常用方法有3種:小樣方大樣本量法、大樣方小樣本量法、大單位內(nèi)子樣方再分亞單位小樣方法。考慮到海草持久性土壤種子庫的種子形態(tài)普遍較小以及在微生境中的高度異質(zhì)性,建議采用小樣方大樣本量的方法,結(jié)果更具代表性和可靠性[31,42]。
由于海草土壤種子庫的高度時空異質(zhì)性,因此土壤種子庫需較高的時空采樣精度[7,43]。每季度/月的采樣可精確描述土壤種子庫大小的季節(jié)/月際變化。然而,土壤種子庫研究耗時耗力,為節(jié)約人力物力,可根據(jù)海草生活史(一年生/多年生)、土壤種子庫類型(短暫種子庫/持久種子庫)以及物候期(如開花結(jié)實的月份)等特點,僅在每年的固定時間開展若干次采樣。此外,如需揭示持久種子庫大小的年際變化,則要開展連續(xù)多年的定位取樣[2]。
為減輕帶回實驗室的樣品重量,通常在野外使用合適孔徑(比目標(biāo)種子的直徑稍小)的網(wǎng)篩或網(wǎng)袋將沉積物樣先篩洗一遍,剩余含種子的沉積物帶回實驗室作進(jìn)一步的處理分析。
3.2 種子庫密度的分析方法
海草土壤種子庫密度的研究方法可分為直接計數(shù)法和幼苗萌發(fā)法兩種,采用何種方法取決于種子的大小以及破除休眠的可控性。兩種方法各有優(yōu)缺點,直接計數(shù)法簡單、相對萌發(fā)法更快捷,但由于有些時候部分種子外觀很完整卻不具備活力,導(dǎo)致結(jié)果偏高(如果不進(jìn)行活力測試的情況下)。直接計數(shù)法是指在實驗室的環(huán)境下,借助于篩網(wǎng)或某些溶液對含種子的沉積物進(jìn)行分選,以肉眼直接觀察或利用放大鏡、顯微鏡等對沉積物中的種子進(jìn)行鑒定和計數(shù)。網(wǎng)篩孔徑的大小或分選溶液的濃度是該項技術(shù)效率的關(guān)鍵[31]。直接計數(shù)法適用于種子個體較大或難于破除休眠的海草種子。
直接計數(shù)法可借助某些溶液,使種子脫離沉積物漂浮到所使用的溶液中,便于種子的鑒定和統(tǒng)計,以提高分析處理的效率。據(jù)報道,采用硅溶膠(Ludox)方法在海草種子庫的分析中已取得了較好的效果,尤其是在研究喜鹽草屬的土壤種子庫時[44-45]。
幼苗萌發(fā)法是指將樣品置于人工氣候箱或植物生長室,控制適宜的光照、溫度、濕度等條件,或者輔助一些物理措施(例如去除種皮),使種子盡可能全部萌發(fā),定時記錄和統(tǒng)計幼苗數(shù),據(jù)此來評估土壤中具有活力的種子數(shù)量。此法對于個體特別小、難以進(jìn)行直接計數(shù),且其休眠機(jī)理已被熟知的海草種子較為適用。但幼苗萌發(fā)法需時較長,因此,休眠機(jī)理尚不清楚,無法破除休眠的海草種子并不適用。此外,若土壤種子庫中存在2種或超過2種海草的種子,但萌發(fā)條件較難同時滿足不同海草種子的萌發(fā)要求,加之有些海草種子的發(fā)芽同步性較低,因此,在觀察期間并非所有具活力的種子都能萌發(fā),以上因素常會造成幼苗萌發(fā)法結(jié)果的低估。直接計數(shù)法和萌發(fā)法的綜合使用可給土壤種子庫密度分析帶來更可靠的評估結(jié)果[31]。
3.3 種子活力測定方法
如果在土壤種子庫密度分析時采用的是直接計數(shù)法,還需要對種子活力進(jìn)行測定,以評估土壤中“真正具有活力”的種子。確定海草種子活力有四唑染色法、直接萌發(fā)法和直接檢驗胚法[31]。四唑染色法在海草種子庫研究中最常用,在鰻草屬和二藥草屬的種子活力測定中得到了較好的應(yīng)用[31,46]。染色前需去除種子外種皮,室溫條件下以5%四唑溶液浸泡24 h,然后對著色種子(被認(rèn)為是有活力的種子)進(jìn)行統(tǒng)計[31]。采用染色法需保證足夠的樣品量(每一重復(fù)種子數(shù)至少25粒)和重復(fù)數(shù)(每一處理至少3個重復(fù)),以獲得準(zhǔn)確的評估結(jié)果[31]。
4 海草土壤種子庫的生態(tài)學(xué)和遺傳學(xué)意義
在評估人為干擾或自然災(zāi)害對植物群落的影響時,往往側(cè)重于現(xiàn)有種群成熟個體的結(jié)構(gòu),而忽略了植物生活史的土壤種子庫階段和種苗補(bǔ)充階段[47]。在傳統(tǒng)的海草生態(tài)監(jiān)測中,也多聚焦于海草的覆蓋度、生物量和密度[48-49],對海草的土壤種子庫并未加以重視。然而,大量的研究表明,與陸生植物一樣,海草的土壤種子庫,尤其是持久種子庫,在植被受干擾后(例如臺風(fēng))的恢復(fù)中常能起到關(guān)鍵作用[8,50]。具有豐富土壤種子庫的海草生態(tài)系統(tǒng)常意味著其具備更大的恢復(fù)力,能從大規(guī)模的干擾中恢復(fù)[51-54]。
種子庫對種群恢復(fù)的重要性因物種、棲息地和生活史特征的差異而有所不同。在空間或時間上受不可預(yù)測干擾的海草生境,持久土壤種子庫具有巨大的恢復(fù)潛能[35]。在所有的海草中,絲粉草屬(Cymodocea)、喜鹽草屬、針葉草屬(Syringodium)和川蔓草屬(Ruppia)可以形成持久性土壤種子庫,而其他屬的海草只能形成短暫種子庫或不能形成種子庫。對能形成持久種子庫的海草而言,種子持續(xù)存在并保持活力直到下一個繁殖周期,在種群受到重大干擾的情況下,它們可以作為一個重要的恢復(fù)源。
海草土壤種子庫對于一年生的海草種群的續(xù)存必不可少,因為這些種群的恢復(fù)和更新幾乎完全依賴于土壤種子庫,缺乏持續(xù)存在的種子庫會降低這些海草種群應(yīng)對脅迫的恢復(fù)力[23,54-56]。例如,喜鹽草屬棲息地的種子庫為昆士蘭的海草恢復(fù)提供了一個重要的物質(zhì)基礎(chǔ);在外界環(huán)境條件不利時,毛葉喜鹽草的恢復(fù)力依賴于土壤種子庫的續(xù)存以及后續(xù)的迅速萌發(fā)生長和繁殖[57]。
中國的海草主要分布于南海區(qū)的海南、廣西、廣東以及黃渤海區(qū)的山東、河北和遼寧。南海區(qū)的海草以卵葉喜鹽草、日本鰻草(Zosterajaponica)和貝克喜鹽草分布較為廣泛,而黃渤海區(qū)則以鰻草及日本鰻草分布較廣泛[58-59]。這些廣泛分布的海草種類均具有土壤種子庫,而且同時具有多年生和一年生的生長形式。盡管到目前為止,未見有人為干擾與海草土壤種子庫相互關(guān)系的報道,但是普遍認(rèn)為種子比植物成株具有更強(qiáng)的脅迫忍耐能力,因此埋藏在土壤中的種子更能逃避干擾、疾病和捕食的損害[35,60]。中國海草場退化嚴(yán)重,人為干擾是導(dǎo)致其退化的主要原因,突出表現(xiàn)為在海草床海域破壞性的挖捕和養(yǎng)殖活動,以及在海草生境和周邊的圍填海活動[58]。這些頻繁的人為干擾活動對海草植被的群落結(jié)構(gòu)造成了負(fù)面影響[61],而土壤種子庫往往比海草植株有更強(qiáng)的生態(tài)恢復(fù)力,因此,保護(hù)好這些海草場的土壤種子庫成為日后這些受損草場恢復(fù)的關(guān)鍵。
此外,土壤種子庫被認(rèn)為是植物種群基因多樣性的潛在提供者,所以土壤種子庫在維持群落生物多樣性和遺傳多樣性方面具有重要意義[34,62]。持久續(xù)存的種子庫是由不同世代、不同年月和不同基因型的種子構(gòu)成,因此一個剛剛萌發(fā)形成的幼苗種群會包含有不同的基因型,構(gòu)成一個“進(jìn)化的記憶”。如果環(huán)境受到干擾而種子混合體被帶到土壤表層,所萌發(fā)長成的植物,將是存在時限很不相同的母株的后代。在這些植物之間,通過開花授粉結(jié)籽過程所進(jìn)行的基因交換,將會對種群的遺傳變異、環(huán)境適應(yīng)和種群進(jìn)化等產(chǎn)生重要的影響[33,63]。種子壽命和休眠越長,生態(tài)學(xué)上積累的變異便越多,遺傳變異潛力就越大[2,64]。因此,可充分利用種子庫的基因庫和進(jìn)化記憶功能,對一些具有長命種子的海草,例如針葉草屬、二藥草屬和喜鹽草屬,開展保護(hù)和進(jìn)化方面的研究。
5 展望與結(jié)論
盡管國內(nèi)外對于海草土壤種子庫給予了越來越多的關(guān)注,但是對于很多基礎(chǔ)性的問題仍然存在較大的不確定性或研究空白。例如,(1)很多海草種子的休眠機(jī)理和壽命仍不清楚,這將影響我們對海草土壤種子庫持續(xù)性的認(rèn)知;(2)在研究方法上,以往多數(shù)的研究均存在樣本量過少以及采樣周期過短的問題,這會對研究結(jié)果的可靠性以及長期動態(tài)變化規(guī)律的闡明產(chǎn)生影響;(3)我國很多海草場都面臨人為干擾的影響,但到目前為止還沒有人為干擾與海草土壤種子庫相互關(guān)系的系統(tǒng)性研究;(4)過去的相關(guān)研究多側(cè)重于海草土壤種子庫本身的定量描述,而沒有將研究跟海草種群的恢復(fù)與重建、生物多樣性保護(hù)等生態(tài)學(xué)、生物學(xué)問題結(jié)合在一起,致使研究的可應(yīng)用性大打折扣。鑒于此,未來的海草土壤種子庫研究,需在種子休眠的機(jī)理和調(diào)控、人為干擾影響、研究方法的改進(jìn)(基于長期定位觀測的小樣方大樣本量的方法)、在受損海草場恢復(fù)與重建中的作用等方面開展更廣泛而深入的研究。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
