谷子SiSULTR2.1基因的生物信息學(xué)分析及其對 硒、硫的響應(yīng)
2022-05-24 01:28:50尹美強溫銀元李露露高志強
激光生物學(xué)報 2022年2期
關(guān)鍵詞:數(shù)據(jù)庫分析
石 堯,尹美強,溫銀元,李露露,孫 敏,高志強
(1. 山西農(nóng)業(yè)大學(xué)農(nóng)學(xué)院,太谷 030801;2. 黃土高原特色作物優(yōu)質(zhì)高效生產(chǎn)省部共建協(xié)同創(chuàng)新中心, 太谷 030801)
谷子[Setaria italica(L.)Beauv]是起源于我國北方的傳統(tǒng)優(yōu)勢農(nóng)作物,具有耐干旱、耐貧瘠、抗逆、適應(yīng)性高等特點。小米是谷子脫殼后的產(chǎn)物,其營養(yǎng)成分與其他禾谷類糧食作物相比相當(dāng)或更優(yōu)[1]。硒于1817年被瑞典化學(xué)家Berzeliu發(fā)現(xiàn),位于元素周期表第四周期VI主族,化學(xué)性質(zhì)與同一主族的硫和碲相似[2]。其是人和動物體內(nèi)所必需的微量元素,對人體有抗癌、抗氧化防衰老、增強免疫力排毒解毒等作用[3]。人體只有在飲食中攝取有機硒這一條補硒途徑,食源性補硒是改善我國居民健康狀況的必然手段。此外,硒在植物體中也具有非常重要的生理生化作用及功能。
硫轉(zhuǎn)運蛋白是主動轉(zhuǎn)運硫酸鹽時需要的載體蛋白,是植物吸收運輸硫的必需蛋白[4]。硒和硫同屬氧族元素,SeO42-和SO42-競爭細胞膜上的硫轉(zhuǎn)運蛋白[5-6]進入植物體內(nèi)。在模式植物擬南芥中共克隆得到12個硫轉(zhuǎn)運蛋白基因,根據(jù)基因序列相似性,其可以分為4個亞族[7]:SULTR1、SULTR2、SULTR3、SULTR4。SULTR2亞族成員為低親和性硫轉(zhuǎn)運蛋白,負責(zé)將硫酸鹽或硒酸鹽從根皮層轉(zhuǎn)運到木質(zhì)部薄壁細胞,進而運往地上部,是長距離運輸?shù)闹饕D(zhuǎn)運蛋白[8]。所有亞族成員共同完成植物體內(nèi)硫酸鹽的吸收轉(zhuǎn)運工作。
硫轉(zhuǎn)運蛋白對SeO42-和SO42-選擇性吸收的能力與外界植物種類、轉(zhuǎn)運蛋白類型及硫酸鹽水平等因素有關(guān)[9]。White等[10]研究發(fā)現(xiàn),高濃度SO42-會在一定程度上抑制植物對SeO42-的吸收。硫酸鹽會抑制擬南芥根系對硒酸鹽的吸收,二者相互競爭硫轉(zhuǎn)運途徑,因而高濃度硒酸鹽會使植物積累大量的硒元素,從而引發(fā)毒性效應(yīng)。Shibagaki等[11]研究發(fā)現(xiàn),在硒酸鹽脅迫下,缺乏硫轉(zhuǎn)運蛋白的擬南芥突變體對硒酸鹽有耐受性,表明硒酸鹽能通過硫酸鹽轉(zhuǎn)運蛋白進入植物體內(nèi)。Awazuhara等[12]研究發(fā)現(xiàn),AtSULTR2.1參與控制硫酸鹽的轉(zhuǎn)移,并可能調(diào)節(jié)擬南芥種子的硫狀態(tài)。
植物體內(nèi)硒的含量與植物對硒的吸收和轉(zhuǎn)運能力密切相關(guān)。本研究以擬南芥(Arabidopsis thaliana)、水稻(Oryza sativa)硫轉(zhuǎn)運蛋白第二亞族基因序列為模板,在谷子基因組數(shù)據(jù)庫篩選出與硫酸鹽轉(zhuǎn)運蛋白基因高度相似的同源序列,通過改良RNAiso Plus法[13]提取到谷子幼苗的RNA,并反轉(zhuǎn)錄得到谷子硫酸鹽轉(zhuǎn)運蛋白基因SiSULTR2.1的cDNA序列。利用生物信息學(xué)方法對基因的序列、系統(tǒng)發(fā)育、結(jié)構(gòu)以及理化性質(zhì)進行分析,利用實時熒光定量RCR(quantitative real-time PCR,qRT-PCR)技術(shù)研究基因在Na2SeO4、Na2SO4處理后不同時間的表達變化,以求為谷子SiSULTR2.1基因硒、硫轉(zhuǎn)運機制的研究提供理論依據(jù)。
1 材料與方法
1.1 材料及處理
以在山西農(nóng)業(yè)大學(xué)申奉試驗地播種的晉谷21號谷子作為試驗材料。取苗期(播種后25 d)完整無病蟲害根、莖、葉部材料和灌漿期(播種后90 d)根、莖、葉、穗部材料,用液氮速凍后置于-80℃冰箱保存?zhèn)溆茫糜赟iSULTR2.1在不同組織中的表達分析。
以晉谷21號為材料,選用10.0 cm×10.0 cm×8.5 cm栽培盆,品氏泥炭土培養(yǎng),每盆播種12粒完整飽滿的種子,出苗后間苗到每盆9株幼苗。發(fā)芽前均以自來水澆灌,發(fā)芽后在2次Hoagland’s(霍格蘭氏)營養(yǎng)液澆灌之間澆灌2次同體積的自來水。將其培養(yǎng)于寧波東南儀器有限公司的RDN-260E-3D型低溫人工氣候箱中,培養(yǎng)溫度為25℃,濕度為60%/40% RH,光照強度為22000 Lx,光照時間為16 h/8 h。在三葉一心期將其分成三組,一組葉面噴施清水作為對照(130 mL/m2),一組葉面噴施12.5 μmol/L Na2SeO4,一組葉面噴施12.5 μmol/L Na2SO4,后兩組噴施體積與對照組清水體積相同。分別在處理0、12、24、48、96 h后,取完整健康幼苗,液氮速凍后置于-80℃冰箱保存?zhèn)溆茫糜赟iSULTR2.1在Na2SeO4、Na2SO4處理后表達分析。
1.2 試驗方法
1.2.1 總RNA提取及cDNA合成
采用改良RNAiso Plus法提取到谷子幼苗的RNA,并通過BioDrop超微量紫外分光光度計檢測提取到的RNA的濃度和純度,總RNA置于-80℃冰箱保存?zhèn)溆谩⑻崛〉玫降墓茸佑酌缈俁NA參照TaKaRa公司反轉(zhuǎn)錄試劑盒PrimeScript?RT Reagent Kit With gDNA Eraser反轉(zhuǎn)錄說明書,利用BIO-GENER基因擴增儀反轉(zhuǎn)錄合成cDNA。
1.2.2 SiSULTR2.1生物信息學(xué)分析
利用蛋白結(jié)構(gòu)域數(shù)據(jù)庫SMART(https://smart.embl-heidelberg.de/smart/show_motifs.pl)進行蛋白結(jié)構(gòu)域分析。利用ExPASy-Protparam tool數(shù)據(jù)庫(http://web.expasy.org/protparam/)進行氨基酸數(shù)目、相對分子質(zhì)量、理論等電點等理化性質(zhì)分析;利用ExPASy-ProtScale數(shù)據(jù)庫(https://web.expasy.org/protscale/)進行蛋白質(zhì)親疏水性預(yù)測;利用蛋白質(zhì)信號肽在線預(yù)測工具SignalP-5.0(https://services.healthtech.dtu.dk/service.php?SignalP-5.0)進行信號肽預(yù)測;利用TMHMM-2.0數(shù)據(jù)庫(https://services.healthtech.dtu.dk/service.php? TMHMM-2.0)預(yù)測蛋白跨膜結(jié)構(gòu);利用DNAMAN軟件對谷子硫酸鹽轉(zhuǎn)運蛋白基因的氨基酸序列進行多序列比對分析;利用MEGA7.0軟件構(gòu)建系統(tǒng)發(fā)育樹;利用數(shù)據(jù)庫PredictProtein(https://predictprotein.org/)預(yù)測蛋白質(zhì)的二級結(jié)構(gòu);利用SWISS-MODEL數(shù)據(jù)庫(https://swissmodel.expasy.org/)預(yù)測蛋白質(zhì)的三級結(jié)構(gòu);利用Cell-PLoc 2.0數(shù)據(jù)庫(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)、WoLF PSORT(https://wolfpsort.hgc.jp/)進行亞細胞定位預(yù)測。
1.2.3SiSULTR2.1表達模式分析
將1.2.1處反轉(zhuǎn)錄合成的cDNA稀釋5倍,-20℃保存,用于后續(xù)qRT-PCR分析。使用軟件Primer Premier 5.0設(shè)計引物(表1),引物由上海生工生物工程股份有限公司合成。qRT-PCR反應(yīng)體系:SYBR Premix Ex Taq 5.0 μL;上、下游引物(10.0 μmol/L)各0.4 μL;ROX Reference DyeⅡ 0.2 μL;cDNA 1.0 μL;加水至總體積10.0 μL;重復(fù)3次。qRT-PCR反應(yīng)程序:95℃預(yù)變性3 min;95℃變性10 s,52℃退火30 s,42個循環(huán);從65℃以每隔5 s上升0.5℃的速度上升至95℃。以Actin為內(nèi)參基因,采用2-△△Ct法分析結(jié)果。

表1 引物序列Tab. 1 Sequence of primers
1.3 數(shù)據(jù)分析
所有試驗均設(shè)3次生物學(xué)重復(fù)。使用SPSS25軟件進行數(shù)據(jù)處理及多重比較,采用Microsoft Excel 2010軟件制表作圖。
2 結(jié)果與分析
2.1 SiSULTR2.1的基因鑒定
查閱文獻并在美國國立生物技術(shù)信息中心(National Center for Biotechnology Information,NCBI)搜索下載獲得擬南芥AtSULTR2.1、水稻OsSULTR2.1基因序列。以獲得的擬南芥、水稻硫轉(zhuǎn)運蛋白基因序列為參考,在Phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)和NCBI(https://www.ncbi.nlm.nih.gov/)數(shù)據(jù)庫中進行Blast檢索和比對,獲得谷子硫轉(zhuǎn)運蛋白基因序列,并將其命名為SiSULTR2.1。
2.2 SiSULTR2.1蛋白的理化性質(zhì)分析
2.2.1 SiSULTR2.1蛋白的保守結(jié)構(gòu)域分析
理化性質(zhì)分析結(jié)果顯示:SiSULTR2.1的蛋白質(zhì)相對分子質(zhì)量為70374.55 Da,氨基酸數(shù)目為656個;理論等電點為8.81,富含堿性氨基酸;帶負電荷的殘基總數(shù)(Asp+Glu)有47個,帶正電荷的殘基總數(shù)(Arg+Lys)有56個;SiSULTR2.1蛋白的分子式為C3195H5112N828O893S30,脂肪指數(shù)為110.61,不穩(wěn)定蛋白指數(shù)為36.88,此類蛋白為穩(wěn)定性蛋白;SiSULTR2.1蛋白中丙氨酸含量最高,占總氨基酸數(shù)的10.8%。
蛋白結(jié)構(gòu)域分析數(shù)據(jù)庫SMART顯示,SiSULTR2.1蛋白具有硫轉(zhuǎn)運蛋白所特有的Sulfate-transp結(jié)構(gòu)域(位置:89~471)和STAS結(jié)構(gòu)域(位置:522~641)(圖1)。

圖1 SiSULTR2.1蛋白的保守結(jié)構(gòu)域Fig. 1 Conservative domains of SiSULTR2.1 protein
2.2.2 SiSULTR2.1蛋白的親疏水性、信號肽及跨膜結(jié)構(gòu)域預(yù)測
利用ExPASy-ProtScale數(shù)據(jù)庫對蛋白的親疏水性進行預(yù)測。如圖2所示,SiSULTR2.1蛋白親水性區(qū)域和疏水性區(qū)域是交叉排列的。Score>0為疏水區(qū),Score<0為親水區(qū),我們可以看到Score>0區(qū)域比Score<0區(qū)域密集。由此可表明,SiSULTR2.1編碼的蛋白是疏水性蛋白。蛋白信號肽預(yù)測結(jié)果顯示(圖3),SiSULTR2.1無信號肽位點,屬于非分泌蛋白。跨膜結(jié)構(gòu)域預(yù)測顯示,SiSULTR2.1有9個跨膜結(jié)構(gòu)域(圖4),表明SiSULTR2.1蛋白屬于跨膜蛋白。

圖2 SiSULTR2.1蛋白的親疏水性預(yù)測Fig. 2 Prediction of the hydrophobicity of SiSULTR2.1 protein
圖3 SiSULTR2.1蛋白的信號肽預(yù)測Fig. 3 Prediction of SiSULTR2.1 protein signal peptide

圖4 SiSULTR2.1蛋白跨膜結(jié)構(gòu)域預(yù)測Fig. 4 Prediction of the transmembrane domain of SiSULTR2.1 protein
2.3 SiSULTR2.1蛋白的系統(tǒng)發(fā)育分析
利用DNAMAN軟件將谷子硫轉(zhuǎn)運蛋白第二亞族基因與水稻、玉米(Zea mays)、大豆(Glycine max)、擬南芥硫轉(zhuǎn)運蛋白第二亞族基因進行多序列比對,發(fā)現(xiàn)其序列比對一致性達到71.79%。將谷子硫轉(zhuǎn)運蛋白氨基酸序列與水稻、玉米、擬南芥硫轉(zhuǎn)運蛋白氨基酸序列分別進行雙序列比對,分析發(fā)現(xiàn)其與水稻、玉米同源序列相似性較高,分別為85.45%、88.91%,與擬南芥同源序列相似性不高,為55.99%。
根據(jù)4個物種的10個硫轉(zhuǎn)運蛋白基因氨基酸序列,利用MEGA7.0軟件構(gòu)建進化樹。由圖5可知,谷子與玉米、水稻聚為一大類,其與玉米親緣關(guān)系最近。谷子、玉米、水稻均為禾本科作物,親緣關(guān)系較近;谷子和玉米均為C4作物,而水稻為C3作物,因而谷子玉米親緣關(guān)系最近。

圖5 SiSULTR2.1蛋白與其他植物SULTR2蛋白的系統(tǒng)進化樹分析Fig. 5 Phylogenetic analysis of protein between SiSULTR2.1 and SULTR2 of other plantsZm:玉米;Os:水稻;Gm:大豆;At:擬南芥。Zm: Zea mays; Os: Oryza sativa; Gm: Glycine max; At: Arabidopsis thaliana.
2.4 SiSULTR2.1蛋白的結(jié)構(gòu)分析
2.4.1 SiSULTR2.1蛋白的二級結(jié)構(gòu)預(yù)測
數(shù)據(jù)庫PredictProtein預(yù)測顯示,谷子硫酸鹽轉(zhuǎn)運蛋白二級結(jié)構(gòu)主要由α-螺旋、β-折疊和無規(guī)則卷曲構(gòu)成。SiSULTR2.1蛋白二級結(jié)構(gòu)α-螺旋、β-折疊和無規(guī)則卷曲占比分別為25.30%、8.54%、66.16%。SiSULTR2.1蛋白含有豐富的α-螺旋和無規(guī)則卷曲結(jié)構(gòu),其中α-螺旋可以形成多個跨膜結(jié)構(gòu)域。
2.4.2 SiSULTR2.1蛋白的三級結(jié)構(gòu)預(yù)測
利用SWISS-MODEL數(shù)據(jù)庫在線構(gòu)建SiSULTR2.1的三維結(jié)構(gòu)模型。結(jié)果顯示,SiSULTR2.1可以較明顯地區(qū)分出STAS結(jié)構(gòu)域和Sulfate-transp結(jié)構(gòu)域(圖6a)。其中STAS結(jié)構(gòu)域(圖6b)位于羧基末端(C端),含有核苷三磷酸(nucleotide triphosphate,NTP)結(jié)合位點[9],通過磷酸化和去磷酸化過程[19]調(diào)控SeO42-的轉(zhuǎn)運活力;Sulfate-transp結(jié)構(gòu)域(圖6c)位于氨基末端(N端),與α-螺旋相互纏繞形成了一個通道結(jié)構(gòu),有利于硒酸鹽的跨膜轉(zhuǎn)運。

圖6 SiSULTR2.1蛋白的三級結(jié)構(gòu)及STAT結(jié)構(gòu)域、Sulfate-transp結(jié)構(gòu)域預(yù)測Fig. 6 Prediction of tertiary structure of SiSULTR2.1 protein, STAS domain and Sulfate-transp domain (a)SiSULTR2.1蛋白的三級結(jié)構(gòu)預(yù)測;(b)STAS結(jié)構(gòu)域;(c)Sulfate-transp結(jié)構(gòu)域。(a) Prediction of tertiary structure of SiSULTR2.1 protein; (b) STAS domain; (c) Sulfate-transp domain.
2.4.3 SiSULTR2.1蛋白的亞細胞定位預(yù)測
利用Cell-PLoc和WoLF PSORT數(shù)據(jù)庫對SiSULTR2.1進行亞細胞定位預(yù)測,結(jié)果略有不同。Cell-PLoc預(yù)測結(jié)果顯示,該蛋白定位于線粒體、葉綠體的可能性不大,定位于細胞膜上的可能性較大。WoLF PSORT預(yù)測結(jié)果顯示,該蛋白定位于液泡膜上。上述結(jié)果表明,SiSULTR2.1可能以一種膜結(jié)合蛋白形式參與SeO42-在細胞的跨膜轉(zhuǎn)運。
2.5 SiSULTR2.1的表達模式分析
2.5.1SiSULTR2.1的組織表達特異性分析
谷子苗期根、莖、葉(圖7a)和灌漿期根、莖、葉、穗(圖7b)的qRT-PCR結(jié)果顯示,SiSULTR2.1在谷子不同部位均有表達,且在不同時期表達量有差異,表現(xiàn)出時空特異性。SiSULTR2.1在苗期莖中的表達量最高,在灌漿期莖、葉片中的表達量均較高。SiSULTR2.1在不同時期根中的表達量均最低,在灌漿期穗中的表達量也較低。
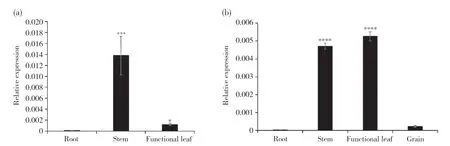
圖7 SiSULTR2.1在谷子不同時期不同組織中的表達Fig. 7 Expression of SiSULTR2.1 in different tissues of millet in different periods(a)SiSULTR2.1在谷子苗期不同組織中的表達;(b)SiSULTR2.1在谷子灌漿期不同組織中的表達。差異顯著性分析采用LSD法。*P<0.01;***P<0.001;****P<0.0001。(a) Expression of SiSULTR2.1 in various tissues of millet at seedling stage; (b) Expression of SiSULTR2.1 in various tissues of millet at filling stag. LSD method was used to analyze the difference significance. *P<0.01; ***P<0.001; ****P<0.0001.
2.5.2SiSULTR2.1在Na2SeO4、Na2SO4處理后的表達分析
由圖8可知,SiSULTR 2.1對Na2SeO4、Na2SO4處理均有表達響應(yīng),但其對Na2SeO4和Na2SO4處理表達響應(yīng)的模式不一致。Na2SeO4處理12 h時,谷子幼苗SiSULTR 2.1的相對表達量與對照組相近,但處理24、48、96 h時幼苗中的相對表達量均顯著高于對照組,且96 h時表達量達到峰值,是對照組的6.75倍。Na2SO4處理12 h時,谷子幼苗SiSULTR 2.1的相對表達量達到峰值,是對照組的17.75倍,處理24、48 h時幼苗中的相對表達量均顯著高于對照組,但與12 h相比表達量呈下降趨勢,且96 h表達量低于對照組。

圖8 SiSULTR2.1在Na2SeO4、Na2SO4處理下的表達分析Fig. 8 Expression analysis of SiSULTR2.1 under various Na2SeO4 and Na2SO4 treatment conditions(a)SiSULTR2.1在Na2SeO4處理下的表達;(b)SiSULTR2.1在Na2SO4處理下的表達。差異顯著性分析采用LSD法。***P<0.001;****P<0.0001。(a) Expression of SiSULTR2.1 in Na2SeO4 treatment conditions; (b) Expression of SiSULTR2.1 in Na2SO4 treatment conditions. LSD method was used to analyze the difference significance. ***P<0.001; ****P<0.0001.
3 討論
SiSULTR2.1編碼的谷子硫轉(zhuǎn)運蛋白含有硫轉(zhuǎn)運蛋白所特有的STAS結(jié)構(gòu)域和Sulfate-transp結(jié)構(gòu)域[14],與前人在茶樹(Camellia sinensis)[9]、馬鈴薯(SolanumtuberosumL.)[15]、短柄草[Brachypodiumdistachyon(L.)Beauv.][16]上的研究結(jié)果一致。該基因編碼的蛋白質(zhì)含有多個跨膜結(jié)構(gòu)域,是一種跨膜轉(zhuǎn)運蛋白。根據(jù)序列同源性,可將SiSULTR2.1歸屬于硫轉(zhuǎn)運蛋白基因家族第二亞族基因,主要負責(zé)硒、硫的遷移轉(zhuǎn)運[17]。SiSULTR2.1是疏水性蛋白,無信號肽序列,屬非分泌蛋白,這些特征與玉米、水稻、薄殼山核桃[Carya illinoinensis(Wangenh.)K. Koch][18]的硫轉(zhuǎn)運蛋白具有高度的相似性。
組織特異性表達結(jié)果揭示,SiSULTR2.1在谷子根、莖、葉、穗中均有表達,且表達模式與張晶晶等[9]在茶樹上的研究結(jié)果相近,具有明顯的組織特異性。SiSULTR2.1在谷子莖、葉片中的表達量較高,但在根、穗中表達量很低。SiSULTR2.1在根中表達量很低,張晶晶[19]的研究中也有類似發(fā)現(xiàn)。SiSULTR2.1在穗中較低的表達量與前人研究結(jié)果有差異,可能與品種的特異性有關(guān)。
本研究發(fā)現(xiàn),SiSULTR2.1對硒、硫處理有不同的響應(yīng)模式。Na2SeO4處理結(jié)果顯示,處理后12 h內(nèi)SiSULTR2.1表達變化量不大,24 h后表達量顯著增加,且在處理后96 h達到最高。SiSULTR2.1在Na2SeO4處理下隨著處理時間的延長,相對表達量總體呈上升趨勢,倪鐘濤等[18]和胡玉榮等[20]也有類似發(fā)現(xiàn)。Na2SO4處理結(jié)果顯示,處理后12 hSi-SULTR2.1的表達就能被顯著誘導(dǎo),處理后24、48 h 其表達量有所下降,處理后96 h表達量恢復(fù)到原來的水平。這可能是因為硫是谷子生長發(fā)育所必須的大量元素,谷子幼苗噴施Na2SO4后,硫轉(zhuǎn)運蛋白基因受SO42-誘導(dǎo)短時間內(nèi)大量表達用以轉(zhuǎn)運硫元素。由此可見,SiSULTR2.1能在短時間內(nèi)迅速響應(yīng)Na2SO4的誘導(dǎo),而響應(yīng)Na2SeO4誘導(dǎo)需要較長時間。Kataoka等[21]研究發(fā)現(xiàn),SULTR2;1與SULTR3;5均表達于根系維管組織中,主要負責(zé)硫酸鹽向地上部運輸,但是二者是否同樣負責(zé)硒酸鹽從根部向地上部的運輸還未可知。
谷子硫轉(zhuǎn)運蛋白基因家族成員眾多,它們均可能在吸收轉(zhuǎn)運Na2SeO4或Na2SO4的某環(huán)節(jié)中有特定的作用[22]。本研究初步探討了谷子SiSULTR2.1對硒、硫的響應(yīng),但該基因在硒、硫的吸收、轉(zhuǎn)運方面具體的功能仍需進一步研究探討。
猜你喜歡
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
財經(jīng)(2017年15期)2017-07-03 22:40:49
財經(jīng)(2017年2期)2017-03-10 14:35:35
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
財經(jīng)(2016年15期)2016-06-03 07:38:02
財經(jīng)(2016年3期)2016-03-07 07:44:46
財經(jīng)(2016年6期)2016-02-24 07:41:51
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
