紫蘇caleosin基因克隆及表達(dá)分析
2022-05-24 01:28:54史華平董書言周雅莉王計(jì)平李潤(rùn)植
激光生物學(xué)報(bào) 2022年2期
關(guān)鍵詞:分析
王 超,史華平,董書言,邢 志,周雅莉,王計(jì)平*,李潤(rùn)植
(1. 山西農(nóng)業(yè)大學(xué)農(nóng)學(xué)院(作物科學(xué)研究所),太谷 030801; 2. 山西農(nóng)業(yè)大學(xué)(山西省農(nóng)業(yè)科學(xué)院)果樹研究所,太谷 030815)
紫蘇(Perilla frutescensL. Britt.)是唇形科紫蘇屬的一年生草本植物,又名香蘇、紅蘇等[1],原產(chǎn)于中國(guó)中南部地區(qū),在我國(guó)已有2000多年的栽培歷史,現(xiàn)廣泛分布于全國(guó)各地[2]。紫蘇在我國(guó)北方主要用作油料作物,少部分作為藥材來使用;在南方地區(qū),紫蘇以藥用為主,食用為輔[3-4]。紫蘇種子含油率高達(dá)45%~55%,其中不飽和脂肪酸含量豐富,占總含油量的90%以上,是日常食用油中不飽和脂肪酸含量較多的一種新型油料作物。
植物種子中儲(chǔ)存著大量中性脂,主要成分為三酰甘油(triacylglycerol,TAG)。TAG在內(nèi)質(zhì)網(wǎng)膜中裝配后向外凸起,磷脂單分子層結(jié)合在其外部形成大小不等的油體,油體融合成一定大小后,油體蛋白對(duì)其進(jìn)行包裹便形成穩(wěn)定的成熟油體,從內(nèi)質(zhì)網(wǎng)釋放到細(xì)胞液中[5]。TAG主要為種子萌發(fā)及幼苗的生長(zhǎng)提供碳源和能量,而油體是負(fù)責(zé)儲(chǔ)存TAG的一個(gè)特殊細(xì)胞器[6]。油體蛋白主要分為三類:油質(zhì)蛋白(oleosin)、油體鈣蛋白(caleosin)和油體甾醇蛋白(steroleosin)[6]。caleosin和oleosin均能使油體更加穩(wěn)定。Chen等[7]分析推測(cè),在使用相同含量的TAG時(shí),等量caleosin能比oleosin覆蓋更多的油體面積,表明caleosin可以更有效地結(jié)合油體,是一種高效油體蛋白[8]。目前,在藻類植物[9]、蓖麻[10]、油菜[11]、小麥[12]、芍藥[13]、水稻[14]、擬南芥[15]等物種中,caleosin基因均已被克隆并進(jìn)行功能分析。
caleosin作為環(huán)化酶催化不飽和脂肪酸,形成環(huán)氧脂肪酸,而環(huán)氧脂肪酸與植物中氧脂素代謝相關(guān),能調(diào)控植物抗逆性并產(chǎn)生自身免疫[16]。在干旱脅迫時(shí),caleosin N端可以感受內(nèi)質(zhì)網(wǎng)膜腔內(nèi)的Ca2+濃度變化,而C端發(fā)生磷酸化修飾,從而在干旱脅迫信號(hào)轉(zhuǎn)導(dǎo)過程中起重要的調(diào)控作用[14]。在水稻[14]、雨生紅球藻[9]、擬南芥[17]等研究中發(fā)現(xiàn),caleosin基因參與了植物對(duì)環(huán)境脅迫的響應(yīng)。本文從紫蘇轉(zhuǎn)錄組數(shù)據(jù)庫中篩選獲得PfClo1基因的開放閱讀框(open reading frame,ORF)序列,并對(duì)其進(jìn)行分子克隆及生物信息學(xué)分析,通過實(shí)時(shí)熒光定量聚合酶鏈?zhǔn)椒磻?yīng)(quantitative real-time polymerase chain reaction,qRT-PCR)研究PfClo1基因在紫蘇不同組織、不同品種及干旱脅迫處理幼苗中的表達(dá)特性,初步探討PfClo1基因的生物學(xué)功能,為進(jìn)一步深入研究PfClo1基因在紫蘇油脂合成中的功能及對(duì)干旱脅迫的響應(yīng)機(jī)制提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
紫蘇品種為山西農(nóng)業(yè)大學(xué)分子農(nóng)業(yè)與生物能源研究所保存的‘晉紫蘇1號(hào)’和‘并紫蘇1號(hào)’。
1.2 試驗(yàn)方法
1.2.1 材料處理
選取均勻一致、籽粒飽滿的‘晉紫蘇1號(hào)’種子放入清洗干凈的小燒杯中,用自來水沖洗20 min,置于10 mL離心管中,用無菌水沖洗3次(每次3 min),然后用75%的酒精消毒處理40 s,無菌水沖洗3次。準(zhǔn)備好鋪有濾紙的培養(yǎng)皿,加入自來水(浸濕濾紙即可),將處理好的紫蘇種子整齊均勻地?cái)[放到濾紙上常溫暗培養(yǎng)。待紫蘇長(zhǎng)出2片子葉時(shí)光照培養(yǎng)(光照16 h、暗培養(yǎng)8 h、濕度80%),待長(zhǎng)出4片真葉時(shí)選取長(zhǎng)勢(shì)良好的幼苗進(jìn)行20%聚乙二醇(polyethylene glycol,PEG)干旱脅迫處理。分別在脅迫處理后0、3、6、9、12、24 h各取5株幼苗全株放入1.5 mL離心管中,做好標(biāo)記后放入液氮中速凍,-80℃保存?zhèn)溆谩?/p>
在大田中取不同油脂含量的紫蘇品種(‘并紫蘇1號(hào)’的含油率為35.6%,‘晉紫蘇1號(hào)’的含油率為46.8%)的根、莖、葉、花及不同發(fā)育時(shí)期的種子(開花后10、20、30、40 d),液氮速凍,-80℃保存?zhèn)溆谩?/p>
1.2.2PfClo1和PfClo2基因的鑒定及生物信息學(xué)分析
在運(yùn)用高通量測(cè)序獲得的紫蘇轉(zhuǎn)錄組數(shù)據(jù)庫(使用根、莖、葉、花、開花后10、20、30、40 d的種子混樣測(cè)定)中篩選獲得PfClo1和PfClo2兩個(gè)基因片段。
通過NCBI-BLAST(http://www.ncbi.nlm.nih.gov/BLAST/)進(jìn)行同源序列比對(duì)后,使用在線分析軟件對(duì)PfClo基因進(jìn)行生物信息學(xué)分析。利用ProtParam在線分析軟件(https//web.expasy.org/protparam/)對(duì)基因進(jìn)行基本理化性質(zhì)的分析;利用PSORT在線分析軟件(https://Psort.hgc.jp/form2.html)對(duì)基因進(jìn)行亞細(xì)胞定位預(yù)測(cè);利用SOPMA在線分析軟件(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)進(jìn)行蛋白質(zhì)二級(jí)結(jié)構(gòu)的預(yù)測(cè);利用SWISS-MODEL在線分析軟件(https://swissmodel.expasy.org/interactive)構(gòu)建蛋白質(zhì)三級(jí)結(jié)構(gòu)模型;利用NCBI-CDD在線分析軟件(https://www.ncbi.nlmnih.gov/structure/cdd/wrpsb.cgi)分析基因的保守域和功能結(jié)構(gòu)域;使用DNAMAN軟件進(jìn)行多序列比對(duì)分析,使用MEGA 7.0、Clustal W軟件進(jìn)行系統(tǒng)發(fā)育樹的構(gòu)建和分析。
1.2.3PfClo1和PfClo2基因在紫蘇不同組織中的表達(dá)特性分析
使用艾德萊生物公司的EASYspin植物RNA快速提取試劑盒提取-80℃保存的紫蘇材料的RNA,使用ABM反轉(zhuǎn)錄試劑盒進(jìn)行反轉(zhuǎn)錄得到cDNA,稀釋至100 ng/μL后-20℃保存?zhèn)溆谩@肞rimer 6.0設(shè)計(jì)熒光定量引物(表1),以Actin2作為內(nèi)參基因。使用2×RealStar Green Power Mixture試劑盒(購自Genstar公司),通過熒光定量分析儀進(jìn)行qRT-PCR擴(kuò)增。qRT-PCR反應(yīng)體系(20.0 μL):2×RealStar Green Power Mixture 10.0 μL、PfClo1/2F 0.5 μL、Pf-Clo1/2R 0.5 μL、cDNA模板 1.0 μL、ddH2O 8.0 μL。擴(kuò)增程序:94℃ 10 min,95℃ 15 s,55℃ 1 min,40個(gè)循環(huán)。每個(gè)樣品重復(fù)3次。通過表達(dá)譜分析,選取PfClo1基因進(jìn)行后續(xù)研究。
1.2.4 引物設(shè)計(jì)
使用Primer 6.0軟件設(shè)計(jì)PCR引物(表1)。
1.2.5PfClo1基因的克隆
通過表達(dá)譜分析,選取PfClo1基因進(jìn)行克隆,利用Primer 6.0設(shè)計(jì)全長(zhǎng)引物(表1),以紫蘇葉片cDNA為模板,按照2íTS-INGKE Master Mix(北京擎科生物公司)試劑盒體系和程序進(jìn)行PCR擴(kuò)增,將PCR產(chǎn)物經(jīng)瓊脂糖凝膠電泳后使用SanPrep柱式DNA回收試劑盒(上海生工生物工程股份有限公司)回收,測(cè)定回收產(chǎn)物的質(zhì)量濃度,連接到Pmd19-T載體中,轉(zhuǎn)化大腸桿菌DH5α,涂板挑取單克隆,菌檢為陽性后送至上海生工生物工程股份有限公司測(cè)序。
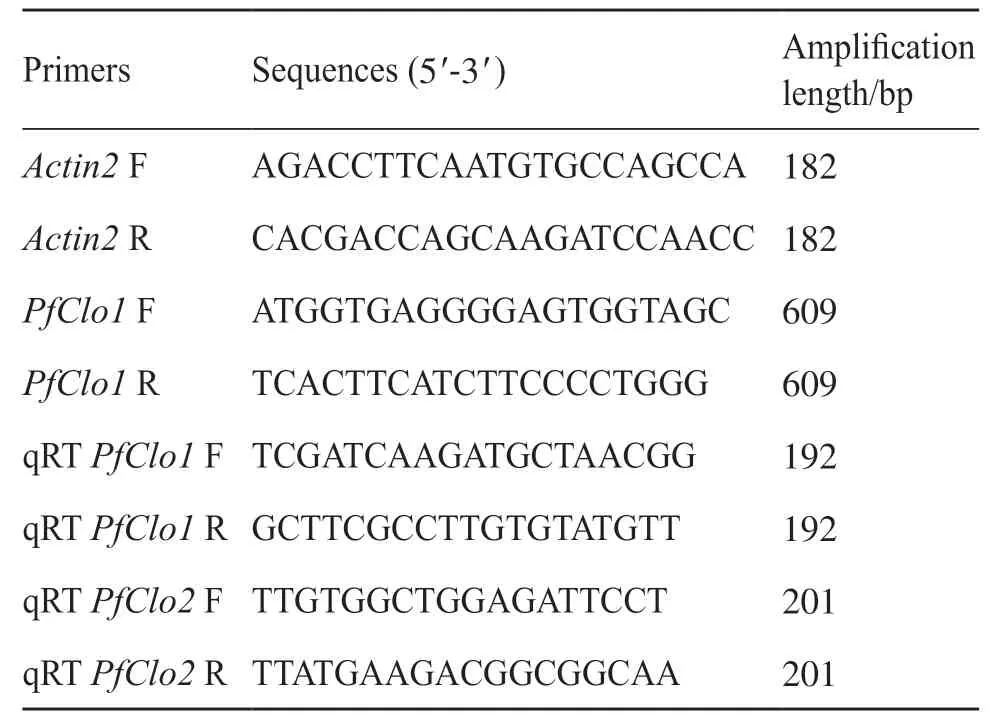
表1 引物信息Tab. 1 Primer information
1.2.6PfClo1基因在不同紫蘇品種中的差異表達(dá)及對(duì)干旱脅迫響應(yīng)分析
提取‘晉紫蘇1號(hào)’和‘并紫蘇1號(hào)’不同組織的RNA及‘晉紫蘇1號(hào)’干旱脅迫處理后的葉片RNA,反轉(zhuǎn)錄成cDNA后通過熒光定量分析儀進(jìn)行qRT-PCR擴(kuò)增。
2 結(jié)果與分析
2.1 PfClo蛋白的理化性質(zhì)分析
PfClo1蛋白的分子式為C1841H3075N609O780S121,相對(duì)分子質(zhì)量為50100.54,理論等電點(diǎn)為5.19(呈酸性),不穩(wěn)定系數(shù)為40.76,屬于不穩(wěn)定蛋白,脂肪酸系數(shù)為26.60,親水性系數(shù)為0.678,屬于疏水性蛋白。PSORT軟件預(yù)測(cè)PfClo1蛋白定位于細(xì)胞質(zhì),屬于胞內(nèi)蛋白。PfClo2蛋白的分子式為C1755H2930N582O732S120,相對(duì)分子質(zhì)量為47743.23,理論等電點(diǎn)為5.18(呈酸性),不穩(wěn)定系數(shù)為40.28,屬于不穩(wěn)定蛋白,脂肪酸系數(shù)為29.73,親水性系數(shù)為0.775,屬于疏水性蛋白。PSORT軟件預(yù)測(cè)PfClo2蛋白定位于細(xì)胞質(zhì),屬于胞內(nèi)蛋白。
2.2 PfClo蛋白的結(jié)構(gòu)分析
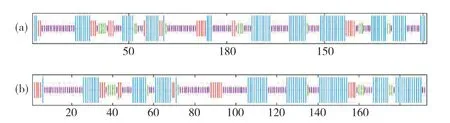
圖1 紫蘇PfClo蛋白二級(jí)結(jié)構(gòu)Fig. 1 The secondary structure of PfClo protein in perilla藍(lán)色線條:α-螺旋;紅色線條:延伸鏈;綠色線條:β-折疊;紫色線條:無規(guī)則卷曲。Blue line: α-helix; Red line: Extended strand; Green line: β-sheet; Purple line: Random coil.
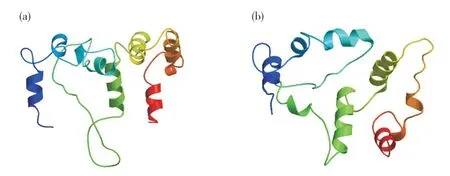
圖2 紫蘇PfClo蛋白三級(jí)結(jié)構(gòu)Fig. 2 The tertiary structure prediction of PfClo protein in perilla(a)PfClo1蛋白;(b)PfClo2蛋白。(a) Pfclo1 protein; (b) Pfclo2 protein.
2.3 PfClo蛋白的多序列比對(duì)分析
利用在線軟件進(jìn)行紫蘇PfClo基因編碼氨基酸的多序列比對(duì)及功能結(jié)構(gòu)域分析,結(jié)果如圖3所示,PfClo蛋白與芝麻(Sesamum indicum,NM_001304394.1)、丹參(Salvia miltiorrhizaXM_042171389.1)、煙草(Nicotiana tabacum,XM_016615461.1)和擬南芥(Arabidopsis,AT4G26740.1)的caleosin家族成員具有相似結(jié)構(gòu),具有鈣結(jié)合基序、酪蛋白激酶Ⅱ磷酸化位點(diǎn)等保守結(jié)構(gòu)域。

圖3 紫蘇與其他物種caleosin的氨基酸序列比對(duì)Fig. 3 Amino acid sequence alignment of caleosin from perilla and other species鈣結(jié)合基序和酪蛋白激酶II磷酸化位點(diǎn)的片段用紅線標(biāo)出,名稱在底部標(biāo)出。Pf:紫蘇;Si:芝麻;Sm:丹參;Nt:煙草;At:擬南芥。The fragments of calcium binding motif and casein kinase II phosphorylation site are marked with red horizontal line and the names are marked at the bottom. Pf: Perilla frutescens; Si: Sesamum indicum; Sm: Salvia miltiorrhiza; Nt: Nicotiana tabacum; At: Arabidopsis thaliana.
2.4 PfClo蛋白的系統(tǒng)進(jìn)化樹分析
在NCBI數(shù)據(jù)庫中使用Clustal W軟件將紫蘇PfClo蛋白與其他8種植物[芝麻(Sesamum indicum,NM_001304394.1)、丹參(Salvia miltiorrhiza,XM_042171389.1)、煙草(Nicotiana tabacumXM_016615461.1)、擬南芥(Arabidopsis,AT4G26740.1)、蓖麻(RicinuscommunisXM_002528321.3)、向日葵(Helianthus annuus,XM_022185293.2)、甘藍(lán)型油菜(Brassica napus,NM_001315834.1)、花生(Arachis hypogaea,XM_025820922.2)]的caleosin序列進(jìn)行多序列比對(duì),并用MEGA7.0軟件構(gòu)建系統(tǒng)進(jìn)化樹(圖4)。可以看出,PfClo1與丹參SmClo聚為一支,均屬于唇形科植物,親緣關(guān)系更為接近;PfClo2與蓖麻RcClo聚為一支,親緣關(guān)系更為接近。

圖4 紫蘇與其他物種caleosin的系統(tǒng)進(jìn)化樹Fig. 4 Phylogenetic tree of caleosin from perilla and other species
2.5 PfClo1和PfClo2基因在‘晉紫蘇1號(hào)’不同 組織中的表達(dá)特性分析
紫蘇PfClo基因在不同組織中的表達(dá)譜分析表明(圖5):PfClo1基因在根、莖、葉中基本不表達(dá),在花中表達(dá)量較高,該基因主要在種子中表達(dá),且隨著種子的發(fā)育表達(dá)量不斷增加,在開花后30 d表達(dá)量達(dá)到最高,是開花后10 d表達(dá)量的22.08倍;PfClo2基因僅在葉和花中微量表達(dá),在根、莖、種子中均不表達(dá)。因此,本研究選取PfClo1基因進(jìn)行后續(xù)試驗(yàn)。

圖5 ‘晉紫蘇1號(hào)’不同組織中PfClo1(a)和PfClo2(b)基因的表達(dá)特性Fig. 5 Expression characteristics of PfClo1 (a) and PfClo2 (b) genes in different tissues of ‘Jinzisu 1’不同小寫字母代表P<0.05水平上的差異顯著。R:根;S:莖;L:葉;F:花;10 d:開花后10 d的種子;20 d:開花后20 d的種子;30 d:開花后30 d的種子;40 d:開花后40 d的種子。下同。Different lowercase letters represent significant differences at the level of P<0.05 . R: Root; S: Stem; L: Leaf; F: Flower; 10 d: Seeds 10 days after flowering; 20 d: Seeds 20 days after flowering; 30 d: Seeds 30 days after flowering; 40 d: Seeds 40 days after flowering. The same below.
2.6 紫蘇PfClo1基因的克隆
從紫蘇轉(zhuǎn)錄組數(shù)據(jù)庫中篩選并克隆獲得Pf-Clo1基因(圖6)。紫蘇PfClo1基因的cDNA全長(zhǎng)序列為609 bp,編碼202個(gè)氨基酸殘基(圖7)。

圖6 PfClo1基因克隆電泳圖Fig. 6 PfClo1 gene cloning electrophoresisM:Marker DL2000。M: Marker DL2000.

圖7 紫蘇PfClo1的核苷酸序列和氨基酸序列Fig. 7 Nucleotide sequence and amino acid sequence of PfClo1起始密碼子用紅色框框出,終止密碼子以星號(hào)指示。The start codon is framed in red, the end codon is indicated by an asterisk.
2.7 PfClo1基因在不同紫蘇品種中的表達(dá)特性分析
紫蘇PfClo1基因在不同紫蘇品種中的表達(dá)特性分析表明(圖8),PfClo1基因在‘并紫蘇1號(hào)’種子中特異性表達(dá),且開花后40 d表達(dá)量最高,但總體來說,相比高油品種‘晉紫蘇1號(hào)’,其相對(duì)表達(dá)水平較低,可見編碼caleosin的PfClo1基因在不同油脂含量的紫蘇材料中存在正相關(guān)關(guān)系。
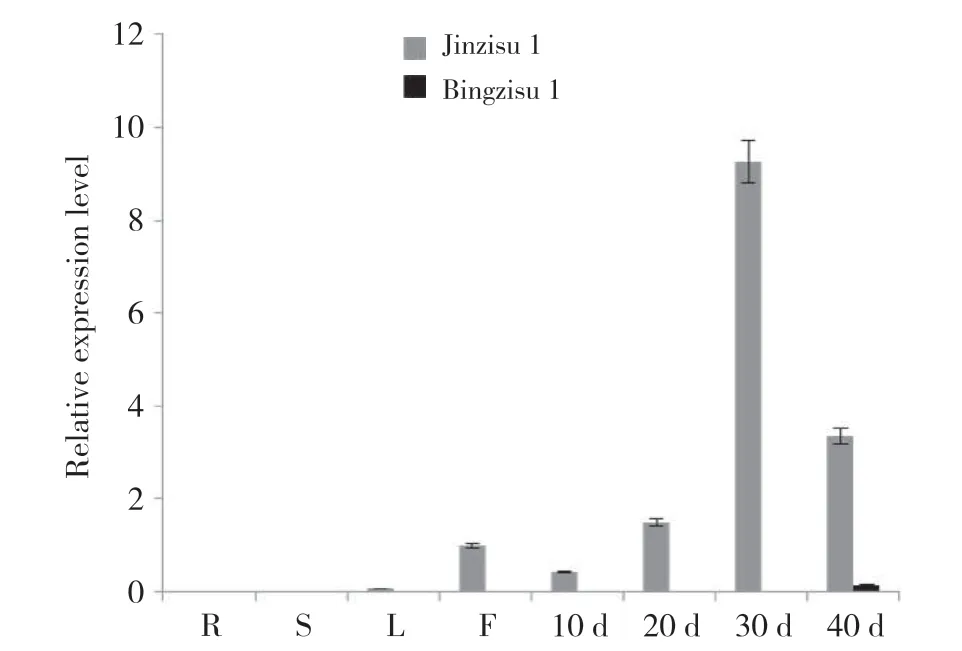
圖8 PfClo1基因在‘晉紫蘇1號(hào)’和‘并紫蘇1號(hào)’中的表達(dá)特性Fig. 8 Expression characteristics of PfClo1 gene in ‘Jinzisu 1’ and ‘Bingzisu 1’ varieties
2.8 干旱脅迫條件下 PfClo1基因在‘晉紫蘇1號(hào)’中的表達(dá)特性分析
PfClo1基因在20% PEG脅迫后的表達(dá)情況分析如圖9所示:隨著脅迫處理時(shí)間的延長(zhǎng),Pf-Clo1基因的表達(dá)量急劇上升,脅迫6 h時(shí)表達(dá)量是3 h表達(dá)量的9.92倍;隨后表達(dá)量繼續(xù)升高,12 h時(shí)PfClo1基因的表達(dá)量達(dá)到最高,為0 h表達(dá)量的6.78倍,3 h表達(dá)量的23.56倍,脅迫處理24 h時(shí)表達(dá)量開始下降。
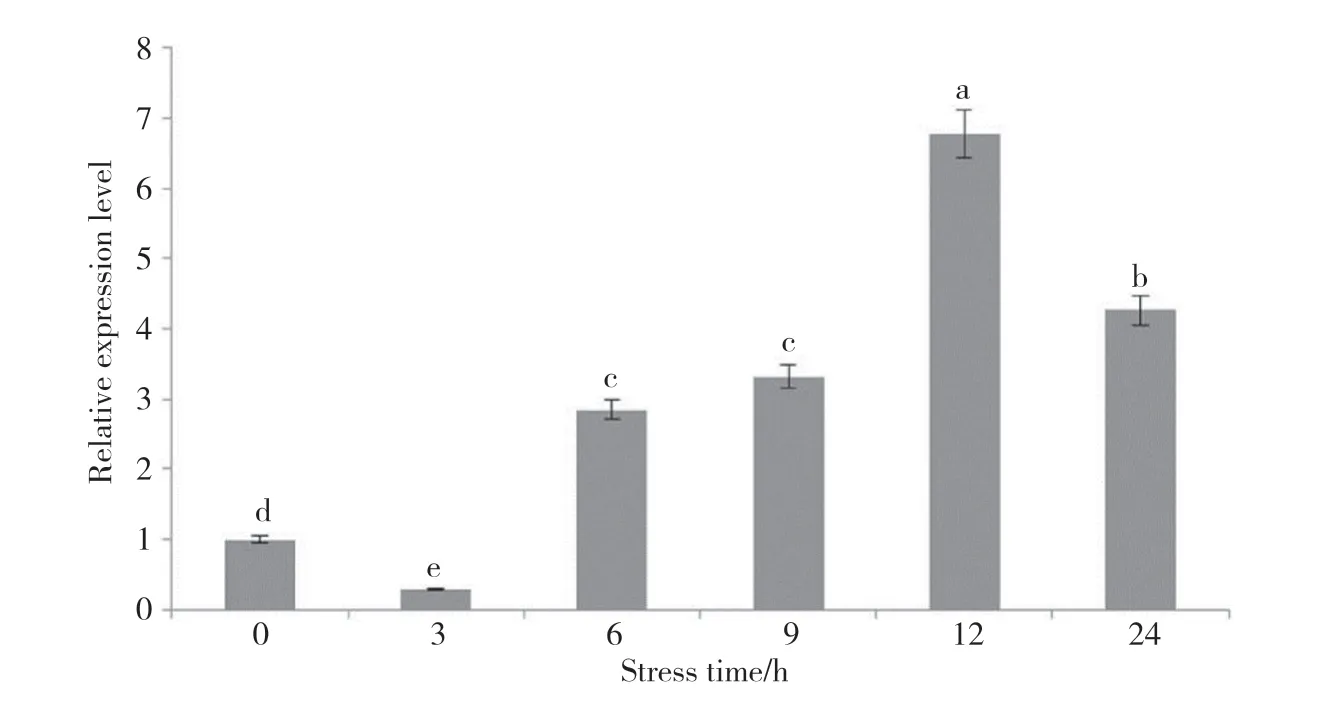
圖9 PfClo1基因干旱條件下在‘晉紫蘇1號(hào)’中的表達(dá)特性Fig. 9 Analysis of the expression characteristics of PfClo1 gene in ‘Jinzisu 1’ under drought conditions不同小寫字母代表P<0.05水平上的差異顯著。Different lowercase letters represent significant differences at the level of P<0.05.
3 討論
油體鈣蛋白基因caleosin在油體形成過程中起重要作用。Khalil等[18]、Marine等[19]研究發(fā)現(xiàn),過表達(dá)caleosin基因有利于合成更多數(shù)量的油體。在酵母中過表達(dá)caleosin基因會(huì)使酵母油脂積累量、油體數(shù)量增加,且油體體積增大[10],表明caleosin基因超表達(dá)可以誘導(dǎo)油脂的合成積累。丁勇等[11]、魏征等[14]的研究結(jié)果表明:油體蛋白N端存在能夠結(jié)合Ca2+的EF-hand區(qū)域,可能發(fā)揮調(diào)控信號(hào)轉(zhuǎn)導(dǎo)途徑的功能;中間部分存在疏水區(qū)域,能增加油體蛋白的疏水性,提高油體的穩(wěn)定性;C端帶有保守蛋白激酶磷酸化位點(diǎn),能啟動(dòng)油體合成途徑。紫蘇PfClo1屬于典型的caleosin家族,其C末端結(jié)構(gòu)域含有酪蛋白激酶II(casein kinase II,CKII)磷酸化位點(diǎn),可推測(cè)其在油體成熟時(shí)參與Ca2+信號(hào)轉(zhuǎn)導(dǎo),這與上述研究結(jié)果一致。多序列比對(duì)及系統(tǒng)進(jìn)化分析驗(yàn)證了PfClo1與芝麻、丹參、擬南芥等植物的caleosin具有相似的結(jié)構(gòu)域及較近的親緣關(guān)系,推測(cè)該基因在紫蘇中編碼caleosin蛋白,調(diào)控油體合成,增加油體的穩(wěn)定性。
caleosin基因在不同植物中的表達(dá)特性不同,在水稻不同組織中均有表達(dá)[14],在芍藥[13]和芝麻[20]種子中其表達(dá)量呈先升高后降低的趨勢(shì)。本研究通過分析紫蘇不同組織及種子發(fā)育不同時(shí)期caleosin基因的表達(dá)特性,發(fā)現(xiàn)其在花和種子中均有表達(dá),在種子發(fā)育后期表達(dá)量達(dá)到高峰,與油體形成時(shí)期相符,推測(cè)該基因高量表達(dá)與油體形成有關(guān);通過分析不同紫蘇品種的差異表達(dá),發(fā)現(xiàn)低油品種‘并紫蘇1號(hào)’的PfClo1基因表達(dá)量極低,推測(cè)編碼油體鈣蛋白的PfClo1基因的表達(dá)與紫蘇油脂的形成存在一定的正相關(guān)關(guān)系。
為豐富滁河左岸堤頂路防浪墻景觀效果,左岸景觀中的防浪墻造型采用異形流線手法,蜿蜒在滁河岸邊,而景墻則采用山水畫卷圖案刻畫在異型混凝土上來顯現(xiàn)。該材料在生產(chǎn)過程中一次成型,后期無需其他工序。所有工作均在工廠完成,只需運(yùn)輸至現(xiàn)場(chǎng)安裝即可,整個(gè)過程實(shí)現(xiàn)PC化,即計(jì)算機(jī)雕刻模型,先做好路徑,然后采用機(jī)器雕刻。
向蘭舟等[17]在擬南芥中研究發(fā)現(xiàn),在高鹽、高溫脅迫及脫落酸(abscisic acid,ABA)誘導(dǎo)條件下,AtCLO3基因表達(dá)均上調(diào),其中高溫脅迫下表達(dá)顯著上調(diào)。魏征等[14]在水稻研究中發(fā)現(xiàn),PEG6000模擬干旱脅迫時(shí),OsCLO-3和OsCLO-6在各個(gè)組織中的表達(dá)水平均有所上調(diào)。本研究通過分析紫蘇在干旱脅迫下caleosin基因的表達(dá)特性,可以看出在干旱脅迫6 h時(shí)表達(dá)量已明顯升高,脅迫12 h時(shí)達(dá)到高峰,表明PfClo1基因具有干旱應(yīng)答功能,能夠快速響應(yīng)干旱脅迫。
參考文獻(xiàn)(References):
[1]回瑞華, 刁全平, 侯冬巖, 等. 紫蘇籽中脂肪酸及主成分α-亞麻酸的分析[J]. 鞍山師范學(xué)院學(xué)報(bào), 2016, 18(4): 24-27.
HUI Ruihua, DIAO Quanping, HOU Dongyan,et al. Fatty acids in perilla seed and principal component analysis of alpha linolenic acid[J]. Journal of Anshan Normal University, 2016, 18(4): 24-27.
[2]譚美蓮, 嚴(yán)明芳, 汪磊, 等. 國(guó)內(nèi)外紫蘇研究進(jìn)展概述[J]. 中國(guó)油料作物學(xué)報(bào), 2012, 34(2): 225-231.
TAN Meilian, YAN Mingfang, WANG Lei,et al. Research progress onPerilla frutescens[J]. Chinese Journal of Oil Crop Sciences, 2012, 34(2): 225-231.
[3]王嵐, 夏繼成. 紫蘇油的研究現(xiàn)狀及應(yīng)用[J]. 黑龍江科技信息, 2011(35): 1.
WANG Lan, XIA Jicheng. Research status and application of perilla oil[J]. Heilongjiang Science and Technology Information, 2011(35): 1.
[4]李鵬, 朱建飛, 唐春紅. 紫蘇的研究動(dòng)態(tài)[J]. 重慶工商大學(xué)學(xué)報(bào) (自然科學(xué)版), 2010, 27(3): 271-275.
LI Peng, ZHU Jianfei, TANG Chunhong. Research progress ofPerilla frutescensand perilla meal[J]. Journal of Chongqing Technology and Business University (Natural Science Edition), 2010, 27(3): 271-275.
[5]SCHMUTZ J, CANNON S B, SCHLUETER J,et al. Genome sequence of the palaeopolyploid soybean[J]. Nature, 2010, 463(7294): 178-183.
[6]沈玥. 油料作物酰基脂類代謝基因和轉(zhuǎn)錄因子及油體鈣蛋白的基因組學(xué)分析[D]. 咸陽: 西北農(nóng)林科技大學(xué), 2016.
SHENG Yue. Genome-wide investigation of acyl-lipid metabolism and caleosin in oilcrops[D]. Xianyang: Northwest A&F University, 2016.
[7]CHEN M, CHYAN C L, LEE T,et al. Constitution of stable artificial oil bodies with triacylglycerol, phospholipid, and caleosin[J]. Journal of Agricultural and Food Chemistry, 2004, 52(12): 3982-3987.
[8]JIANG P L, TZEN J T C. Caleosin serves as the major structural protein as efficient as oleosin on the surface of seed oil bodies[J]. Plant Signaling & Behavior, 2010, 5(4): 447-449.
[9]朱琴, 張宏江, 張春輝, 等. 雨生紅球藻caleosin基因的克隆、 序列分析及表達(dá)特性[J]. 激光生物學(xué)報(bào), 2020, 29(6): 529-537.
ZHU Qing, ZHANG Hongjiang, ZHANG Chunhui,et al. Molecular cloning, sequencing analysis and expression characteristics ofcaleosinfromHaematococcus pluvialis[J]. Acta Laser Biology Sinica, 2020, 29(6): 529-537.
[10]李傳寶. 蓖麻Caleosin基因的克隆及功能驗(yàn)證[D]. 晉中: 山西農(nóng)業(yè)大學(xué), 2016.
LI Chuanbao. Cloning and function analysis ofCaleosingene fromRicinus communis[D]. Jinzhong: Shanxi Agricultural University, 2016.
[11]丁勇, 陳慶波, 徐春雷, 等. 油菜油體鈣蛋白基因BnClo1的克隆和表達(dá)[J]. 作物學(xué)報(bào), 2008,34(11): 1921-1928.
DING Yong, CHENG Qingbo, XU Chunlei,et al. Cloning and expression ofBnClo1gene fromBrassica napus[J]. Acta Agronomica Sinica, 2008, 34(11): 1921-1928.
[12]JAMME F, VINDIGNI J D, MECHIN V,et al. Single cell synchrotron FT-IR microspectroscopy reveals a link between neutral lipid and storage carbohydrate fluxes inS. cerevisiae[J]. PLoS One, 2013, 8(9): e74421.
[13]孟家松, 張夏燕, 方紫雯, 等. 芍藥油體鈣蛋白基因PlClo克隆及表達(dá)分析[J]. 植物生理學(xué)報(bào), 2019, 55(3): 274-280.
MENG Jiasong, ZHANG Xiayan, FANG Ziwen,et al. Cloning and expression analysis of herbaceous peonycaleosingenePlClo[J]. Plant Physiology Journal, 2019, 55(3): 274-280.
[14]魏征, 馬紅, 葛曉春. 水稻油體鈣蛋白家族的進(jìn)化及對(duì)干旱脅迫的響應(yīng)性分析[J]. 科學(xué)通報(bào), 2011, 56(20): 1612-1621.
WEI Zheng, MA Hong, GE Xiaochun. Phylogenetic analysis and drought-responsive expression of the ricecaleosingene family[J]. Chinese Science Bulletin, 2011, 56(20): 1612-1621.
[15]SHEN Y, XIE J, LIU R D,et al. Genomic analysis and expression investigation ofcaleosingene family inArabidopsis[J]. Biochemical and Biophysical Research Communications, 2014, 448(4): 365-371.
[16]MOSBLECH A, FEUSSNER I, HEILMANN I. Oxylipins: structurally diverse metabolites from fatty acid oxidation[J]. Plant Physiology and Biochemistry, 2008, 47(6): 511-517.
[17]向蘭舟, 胡婭晴, 謝涵, 等. 擬南芥油體鈣蛋白基因CALEOSIN 3對(duì)脅迫環(huán)境及ABA誘導(dǎo)的響應(yīng)[J/OL]. 分子植物育種: 1-12[2021-10-20]. http://kns.cnki.net/kcms/detail/46.1068.S.20210122.1218.004.html.
XIANG Lanzhou, HU Yaqing, XIE Han,et al. Response ofArabidopsisoil body protein geneCALEOSIN 3to stress environment and ABA induction[J/OL]. Molecular Plant Breeding: 1-12[2021-10-20]. http://kns.cnki.net/kcms/detail/46.1068.S.20210122.1218.004.html.
[18]KHALIL H B, BRUNETTI S C, PHAM U M,et al. Characterization of thecaleosingene family in the triticeae[J]. BMC Genomics, 2014, 15(1): 239-239.
[19]MARINE F, SABINE D, CELINE B,et al. Heterologous expression ofAtClo1, a plant oil body protein, induces lipid accumulation in yeast[J]. FEMS Yeast Research, 2009, 9(3): 428-438.
[20]CHEN J C F, TSAI C C Y, TZEN J T C. Cloning and secondary structure analysis of caleosin, a unique calcium-biding protein in oil bodies of plant seeds[J]. Plant & Cell Physiology, 1999, 40(10): 1079-1086.
[21]FRANDSEN G I, MUNDY J, TZEN J T C. Oil bodies and their associated proteins, oleosin and caleosin[J]. Physiologia Plantarum, 2001, 112(3): 301-307.
[22]NAESTED H, FRANDSEN G, JAUH G Y,et al. Caleosins: Ca2+-binding proteins associated with lipid bodies[J]. Plant Molecular Biology, 2000, 44(4): 463-476.
[23]周雅莉, 安茜, 任文燕, 等. 紫蘇PfPDAT1基因序列及表達(dá)特性分析[J]. 山西農(nóng)業(yè)大學(xué)學(xué)報(bào) (自然科學(xué)版), 2018, 38(12): 44-49.
ZHOU Yali, AN Xi, REN Wenyan,et al. Analysis ofPfPDAT1gene sequence and expression characteristics inPerilla frutescens[J]. Journal of Shanxi Agricultural University (Natural Science Edition), 2018, 38(12): 44-49.
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
