忍冬屬黃酮合成酶基因FNSⅡ密碼子偏好性及進化分析
2022-05-27 05:03:02何志敏曾子豪朱肖霞余清婷胡珊巍滿威劉素純
中國中醫藥信息雜志 2022年5期
何志敏,曾子豪,朱肖霞,余清婷,胡珊巍,滿威,劉素純
1.長沙學院生物與環境工程學院,湖南 長沙 410003;2.湖南農業大學食品科技學院,湖南 長沙 410128
《中華人民共和國藥典》自2005年版起增加了金銀花中黃酮類化合物木犀草苷含量的測定,但對歸屬山銀花的4個種屬(灰氈毛忍冬、紅腺忍冬、華南忍冬、黃褐毛忍冬)中木犀草苷的含量未做出明確規定,所以木犀草苷成為區分金銀花與山銀花的一個重要化學指標,也是金銀花與山銀花一物一名分列的重要依據。
黃酮合成酶基因(FNSⅠ和FNSⅡ)是植物黃烷酮轉化為黃酮的關鍵酶基因,植物內木犀草苷的含量受黃酮合成酶基因表達的調控,基因表達的高低與其在長期進化過程中形成的密碼子偏好性有關。研究表明,忍冬黃酮合成酶基因FNSⅡ編碼的木犀草苷酶在金銀花和山銀花中的酶催化活性大小是影響木犀草苷生物合成的關鍵因素。
目前已有許多植物的FNSⅡ基因的cDNA被克隆和表征,包括大豆、甘草、苜蓿、擬南芥、非洲菊、高粱等。在長期進化過程中,由于環境壓力及突變選擇,不同物種會傾向性選擇不同的密碼子模式,形成密碼子偏好性。提高FNSⅡ基因表達效率對其功能研究具有重要意義,而其密碼子偏好性與其表達效率有必然聯系。通過密碼子偏好性分析,不僅可以了解不同物種之間的遺傳進化關系,還能對基因的外源表達選擇優先宿主,通過優化密碼子以提高基因的外源表達效率。
本研究對23個不同物種的FNSⅡ基因及本實驗室克隆獲得的29條忍冬屬FNSⅡ基因序列進行密碼子偏好性及進化分析,并進行忍冬FNSⅡ的外源表達系統和遺傳轉化受體的選擇分析,為進一步開展忍冬FNSⅡ基因的功能研究及外源高表達提供依據。
1 資料
通過美國國家生物技術信息中心(NCBI)數據庫(http://www.ncbi.nlm.nih.gov)下載包括金銀花、灰氈毛忍冬、煙草等23個具有完整編碼區(CDS)植物FNSⅡ基因序列,另外29條序列為本實驗室從已通過鑒定的忍冬屬植物金銀花、灰氈毛忍冬、黃褐毛忍冬、凈花菰腺忍冬、紅腺忍冬中克隆獲得,來源信息見表1。
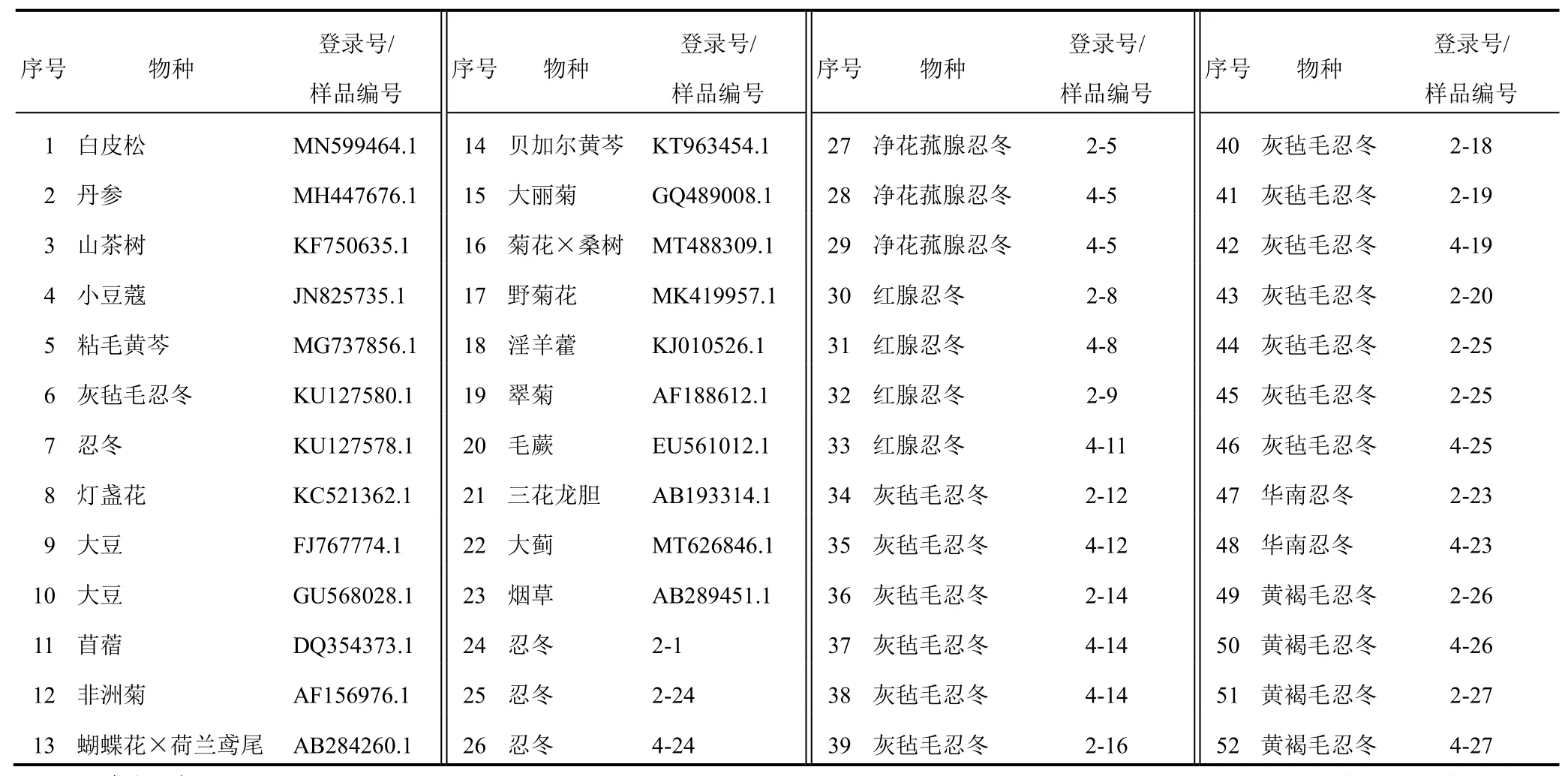
表1 23個物種FNSⅡ基因序列及29條忍冬屬FNSⅡ克隆序列來源信息
2 方法與結果
2.1 密碼子偏性分析
將NCBI下載的23個物種FNSⅡ基因序列去除5’UTR和3’UTR序列,選擇CDS序列,在txt文檔中建立fasta格式,導入Codon W軟件,對52條FNSⅡ基因序列進行分析,獲得密碼子偏好性相關參數,包括相對同義密碼子使用度(RSCU)、密碼子第三位各堿基含量、有效密碼子數(ENC)、密碼子適應指數(CAI)、密碼子偏愛指數(CBI)、最優密碼子使用頻率(FOP)等。
2.1.1 FNSⅡ相對同義密碼子使用度
Codon W分析獲得52條FNSⅡ序列的RSCU值并將RSCU值標準化后,獲得FNSⅡ的RSCU值熱圖。
不同物種FNSⅡ序列的RSCU有一定差異。23個物種的FNSⅡ序列中,最優密碼子為CUC,其均值為1.63。丹參、粘毛黃芩和貝加爾黃芩RSCU>2的密碼子最多,均有7個,見圖1。粘毛黃芩和貝加爾黃芩的密碼子RSCU十分相近,兩者與丹參在多個密碼子上的RSCU值均大于1,在密碼子偏好性上有一定相似性,表明其在進化過程中親緣關系較近。
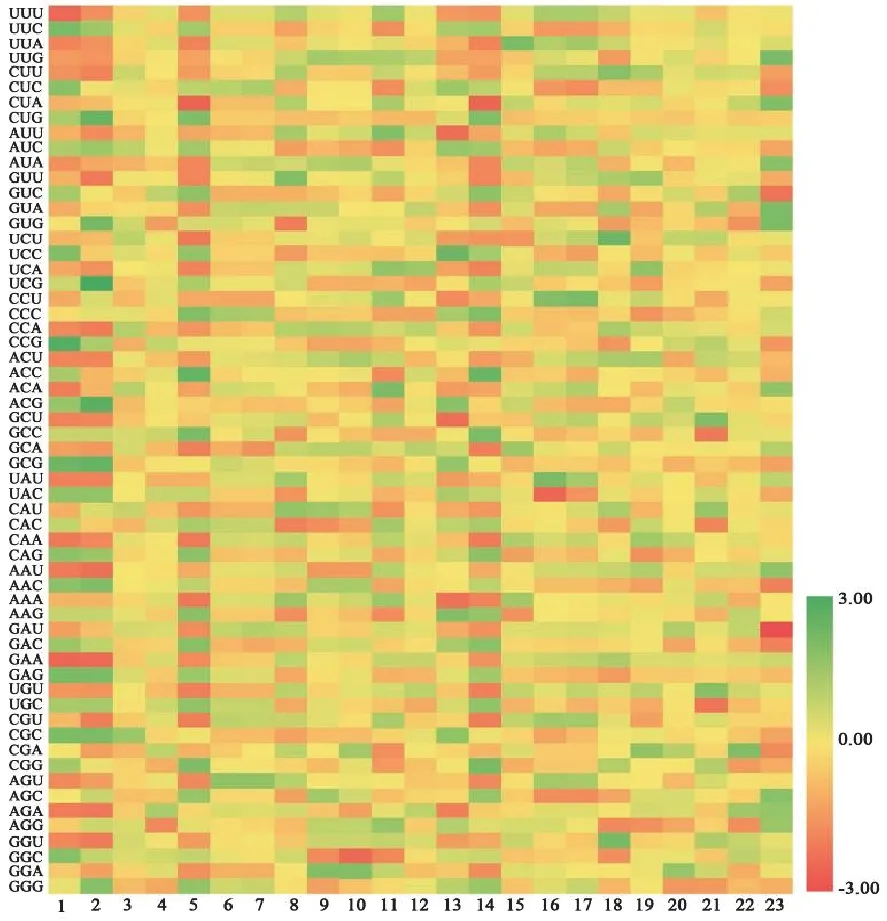
圖1 23個物種FNSⅡ序列的RSCU熱圖
31條忍冬屬FNSⅡ序列RSCU熱圖見圖2。忍冬屬的最優密碼子也為CUC,其均值為2.70,表明大多數密碼子在FNSⅡ中的相對使用頻率較高。凈花菰腺忍冬(2-5)和紅腺忍冬(2-8)的最優密碼子CUC的RSCU值達3.29,表明兩者的FNSⅡ序列在表達時對CUC的偏好性較強。
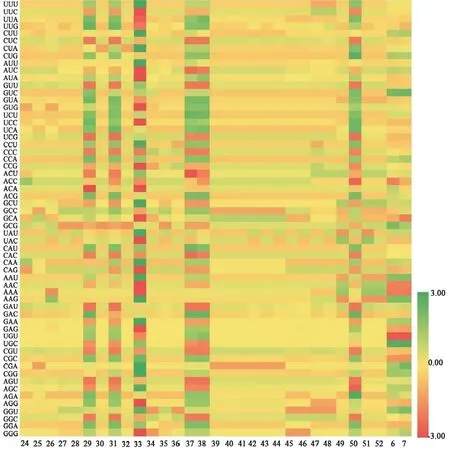
圖2 31條忍冬屬FNSⅡ序列的RSCU熱圖
2.1.2 FNSⅡ特性分析
同義密碼子的差異通常體現在密碼子第3位。在23個不同物種的FNSⅡ序列中,單堿基U3s、C3s、A3s、G3s平均含量分別為0.306、0.350、0.295、0.312,GC3s和GC平均含量分別為0.514和0.469。但白皮松、丹參、粘毛黃芩、蝴蝶花×荷蘭鳶尾、貝加爾黃芩GC3s含量均高于70%,GC含量達56%以上,而其他物種的GC含量與GC3s含量相當,大部分在45%~50%,少部分如燈盞花、苜蓿、淫羊藿等的GC和GC3s含量低于40%。忍冬屬FNSⅡ序列GC3s和GC平均含量分別為0.521和0.459,與其他物種的平均含量相當,結果見表2。表明大部分植物的FNSⅡ序列第3位堿基對G/C和A/T結尾的偏好程度相似。
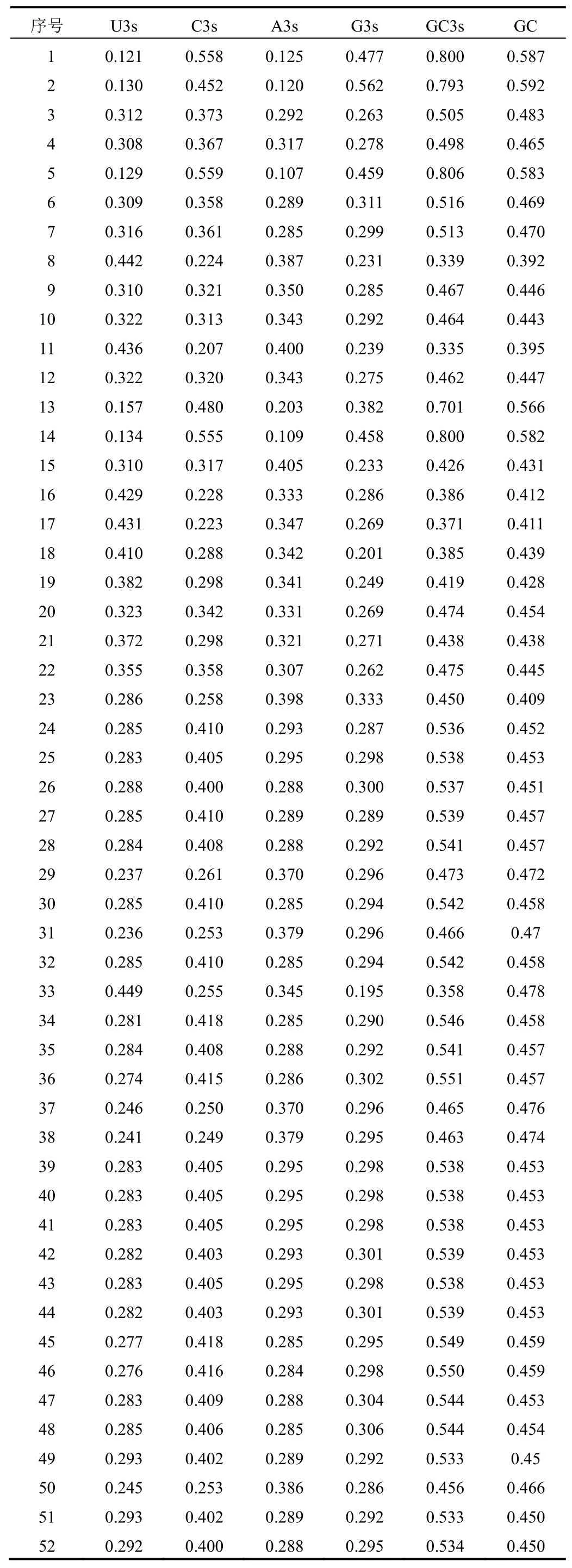
表2 52條FNSⅡ基因密碼子堿基組成特性分析
為系統評估FNSⅡ基因的密碼子使用偏好性程度,對不同物種的FNSⅡ基因密碼子偏好性參數進行分析。從整體來看,各物種的ENC介于39~58,平均值為52.29,忍冬屬FNSⅡ的ENC介于50~57,平均值為52.66。FOP值介于0.326~0.528,平均值為0.392,表明大部分FNSⅡ基因的密碼子偏好性較弱。同時,不同植物FNSⅡ的CAI較小(0.117~0.257),且大部分植物FNSⅡ的ENC值較大。52條FNSⅡ基因的密碼子偏好性參數見表3。分析結果表明,FNSⅡ基因密碼子偏好性弱,且在不同植物中的表達水平可能較低。此外,不同植物FNSⅡ的ENC值存在一定差異,表明FNSⅡ基因在不同植物中的表達水平不同。
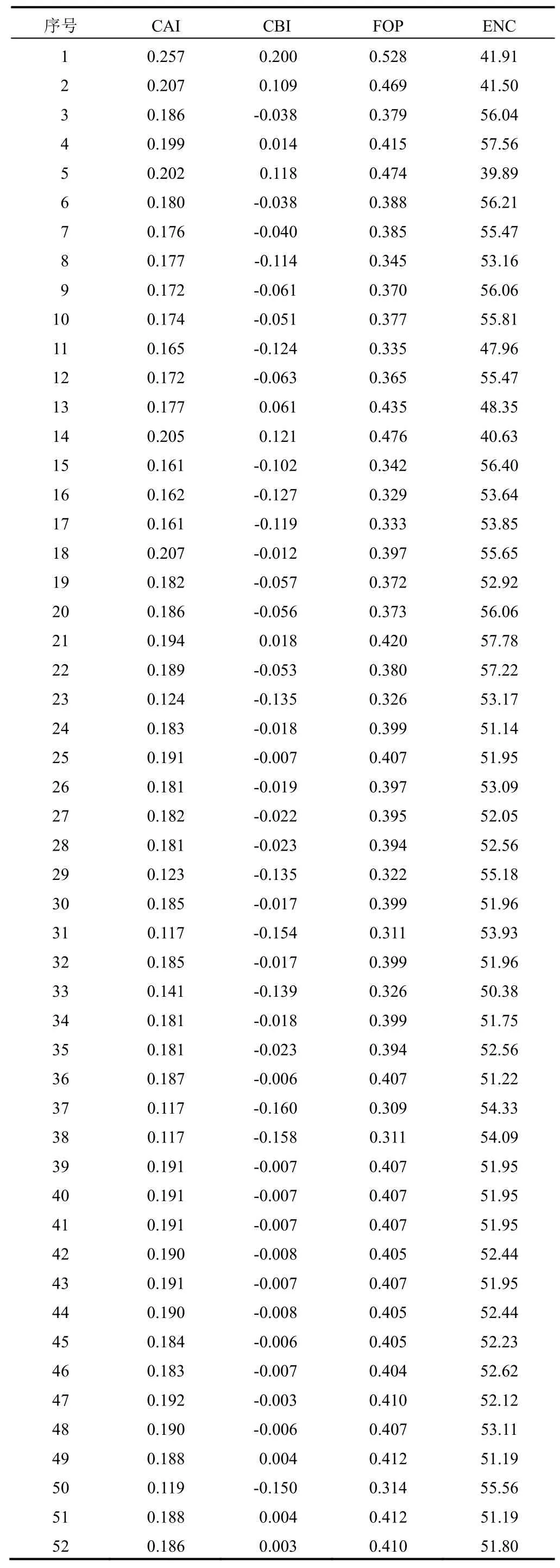
表3 52條FNSⅡ基因密碼子偏好性參數
2.2 ENC-Plot分析
以GC3s為自變量,以ENC為因變量,按ENC=2+GC3s+29/[GC3s+(1-GC3s)]建立期望曲線。將所得各序列以GC3s為橫坐標、ENC為縱坐標繪制散點圖,不同物種的FNSⅡ基因ENC-Plot圖見圖3。可以看出,在期望曲線附近分布著大部分植物FNSⅡ基因,距離期望曲線最近的是燈盞花、淫羊藿、三花龍膽,該結果表明,突變壓力是這些植物FNSⅡ基因密碼子偏好性形成的主要原因。而自然選擇主導煙草、忍冬、翠菊、苜蓿的FNSⅡ基因密碼子偏好性的形成,其分布于離期望曲線較遠的位置。
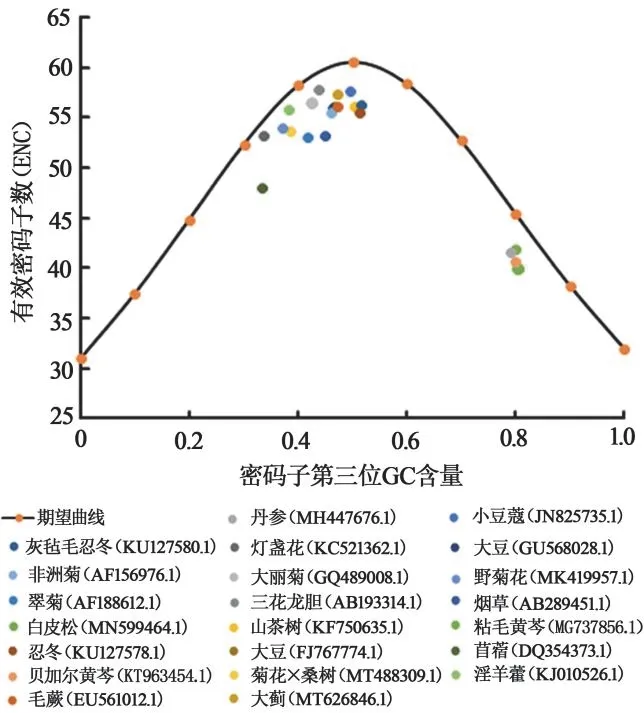
圖3 不同物種FNSⅡ基因ENC-Plot分析
忍冬屬FNSⅡ基因普遍分布于離期望曲線稍遠的位置,尤其是灰氈毛忍冬、華南忍冬、黃褐毛忍冬,結果見圖4,表明忍冬屬植物FNSⅡ基因密碼子偏好性形成主要受自然選擇的影響。
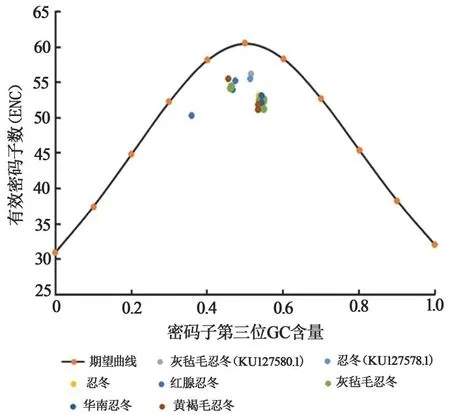
圖4 忍冬屬FNSⅡ基因ENC-Plot分析
2.3 FNSⅡ系統進化分析
將列入所有序列的fasta文件導入Clustal X軟件,進行序列比對后,選擇bootstrap N-J tree聚類分析(bootstrap=1 000),輸出.phd的結果文件,將該文件導入MEGA4.0軟件,構建系統發育樹,結果見圖5。通過鄰接法構建的基于52條FNSⅡ CDS序列系統進化樹主要分為忍冬屬和非忍冬屬兩大分支,符合一般植物分類的規律。當中可以發現忍冬屬植物中大部分聚為一個大類,各FNSⅡ按植物親緣關系遠近聚為一類,但除黃褐毛忍冬和華南忍冬外,其他忍冬屬植物之間有混合交錯分布而聚為一類的現象,而華南忍冬甚至與非忍冬屬植物聚為一類。同比之下,非忍冬屬植物較符合分類規律,同種屬的2種大豆,粘毛黃芩和貝加爾黃芩,同為菊科的野菊花、燈盞花、大薊等分別聚為一類,親緣關系較遠的三花龍膽、山茶樹、苜蓿則獨自分為一類。
2.4 聚類分析
利用SPSS18.0軟件,以52條FNSⅡ序列RSCU為變量,采用組間連接法,進行密碼子偏好性聚類分析。基于FNSⅡ基因RSCU的聚類分析結果(見圖5)與基于CDS序列的系統進化分析結果(見圖6)基本一致。在RSCU聚類中,絕大多數忍冬屬植物聚為一個大類,但在另一小分支,灰氈毛忍冬、凈花菰腺忍冬、黃褐毛忍冬、紅腺忍冬及煙草聚為一類,燈盞花、非洲菊等菊科植物聚成一類,但夾雜了小豆蔻和3個忍冬屬樣本;黃芩屬植物、丹參、蝴蝶花×荷蘭鳶尾、白皮松聚為一類;大豆、山茶樹、淫羊藿、三花龍膽聚為一類。表明其存在核酸序列一致性較高的FNSⅡ基因。同為忍冬屬植物的FNSⅡ基因分布于多個類別,其中14個灰氈毛忍冬的FNSⅡ基因分布于多個大類,與其他植物聚成一小支,與系統進化樹的歸類情況相似但存在一定差異。同種植物的FNSⅡ基因被聚類到不同分支上,說明其FNSⅡ基因具有密碼子偏好性差異。以上結果表明,FNSⅡ基因的密碼子偏好性差異大小與物種親緣關系遠近有一定關系,反映了物種的進化關系,但基于RSCU的聚類在反映物種進化關系準確度方面遜于基于CDS序列的系統進化樹。
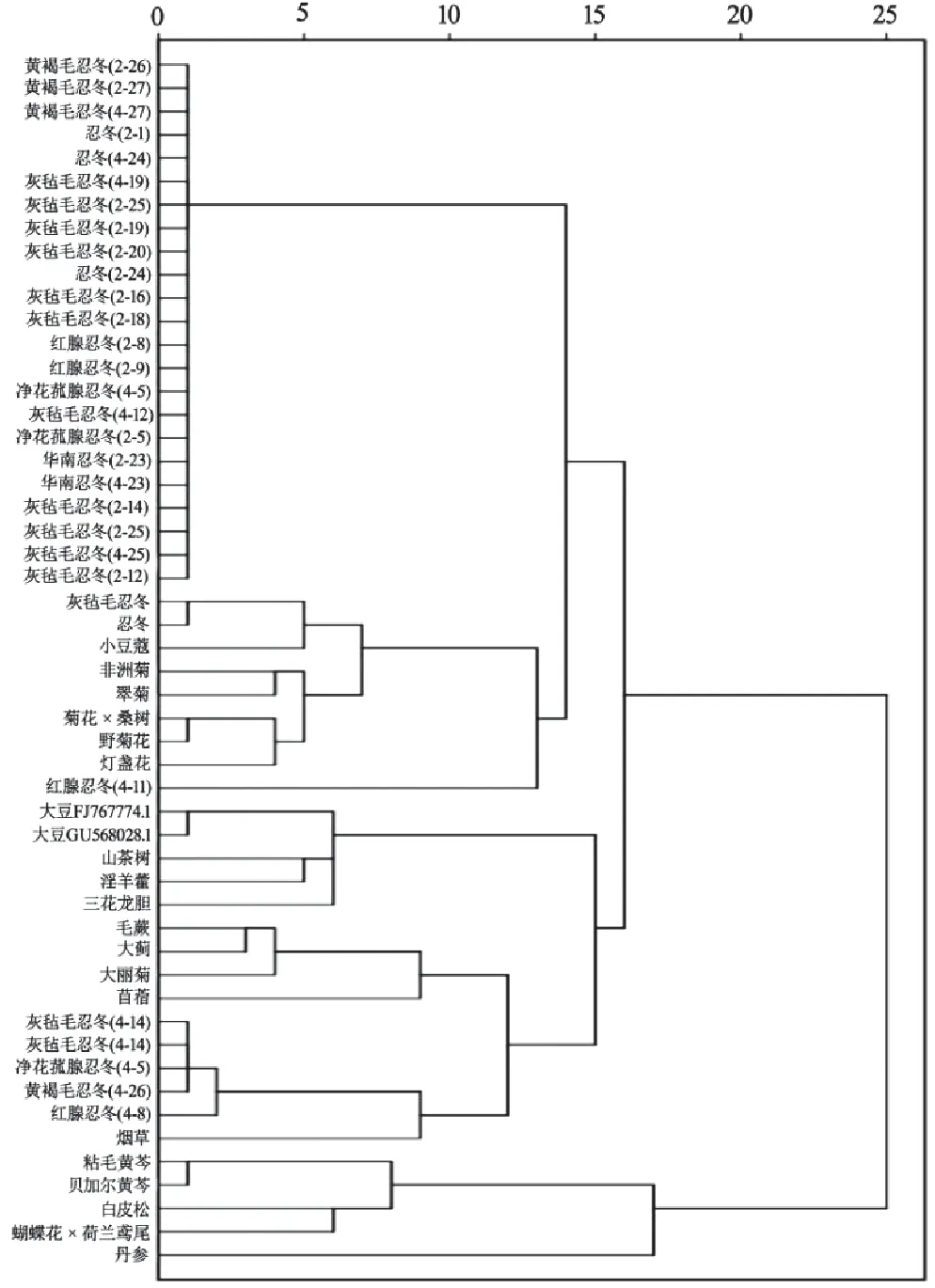
圖5 FNSⅡ的RSCU聚類分析樹狀圖
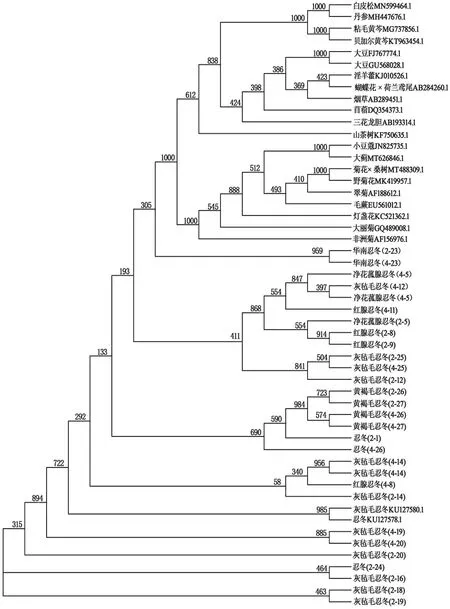
圖6 FNSⅡ系統進化分析
2.5 忍冬FNSⅡ與模式生物密碼子偏好性比較
通過比較忍冬()與模式生物擬南芥()、煙草L.()、大腸桿菌()和酵母()FNSⅡ基因的密碼子使用頻率,分析頻率差異性較大(頻率比值<0.5或>2)的密碼子個數,以確定最佳遺傳轉化受體和外源表達系統,結果見表4。大腸桿菌、酵母與忍冬FNSⅡ基因差異較大的密碼子個數分別為34、31,說明忍冬FNSⅡ的最佳外源表達是酵母,但實際應用時需要對外源表達的FNSⅡ密碼子進行優化以提高表達效率。擬南芥、煙草與忍冬FNSⅡ的差異性密碼子數分別為26、27,說明兩者均可作為忍冬FNSⅡ的遺傳轉化受體。
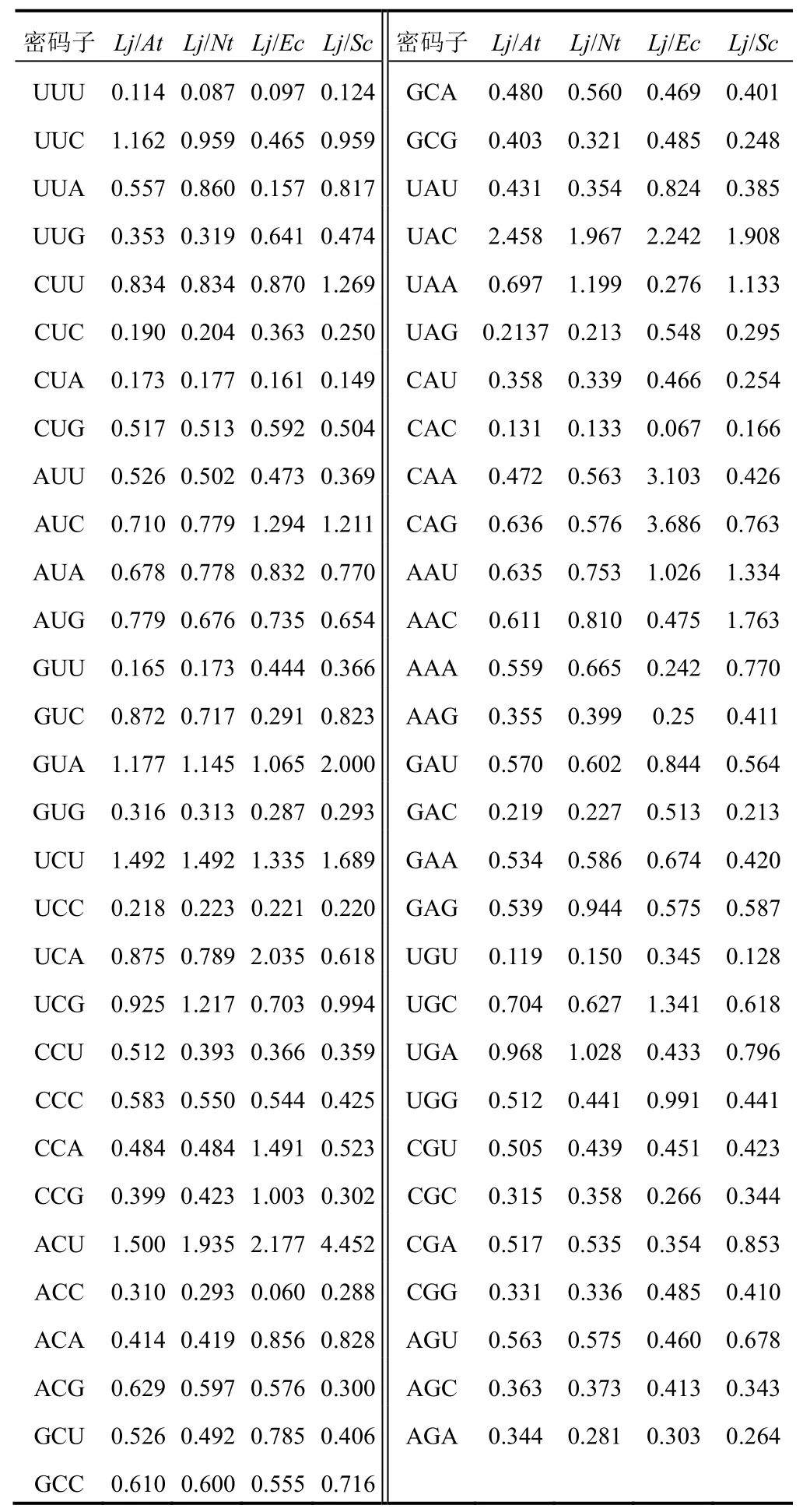
表4 忍冬FNSⅡ與模式生物基因組密碼子使用頻率比較
3 討論
本研究首先對23條不同種植物及29條本實驗室克隆的忍冬屬FNSⅡ基因進行密碼子偏好性整體分析,發現不同種植物FNSⅡ及忍冬屬FNSⅡ的最優密碼子為CUC。通過分析不同物種RSCU>2的偏好性密碼子發現,丹參、粘毛黃芩和貝加爾黃芩中均有7個,而忍冬屬植物除紅腺忍冬(4-11)有5個外,其余均為2個,表明忍冬屬植物FNSⅡ基因對優勢密碼子的使用偏好性較弱。金銀花和山銀花FNSⅡ的密碼子RSCU都具有很高的相似性。進一步分析ENC和FOP,發現23個不同種植物FNSⅡ基因的ENC均值為52.29,忍冬屬植物FNSⅡ基因的ENC均值為52.66,52個樣本的FOP均值接近0.36,表明所選52個樣本FNSⅡ基因的密碼子偏好性普遍較弱。上述ENC和RSCU分析結果均表明植物FNSⅡ密碼子偏好性較弱。CAI與基因表達水平通常呈正相關,而ENC則相反。本研究52條FNSⅡ序列的CAI均小于0.3,而ENC均大于50,表明FNSⅡ基因表達水平較低,而低水平表達的基因往往對密碼子的偏好性也較低,與前述分析結論一致。ENC-plot分析結果表明,突變壓力是大部分植物FNSⅡ基因偏好性形成的主因,僅有少數如煙草、忍冬、翠菊、苜蓿的FNSⅡ基因主要受自然選擇影響或其他因素的影響,如基因長度、結構等。
親緣關系較近的物種在密碼子使用偏好性方面趨向于相同或相近。本實驗室克隆的忍冬屬FNSⅡ基因在各種屬間的同源性高達95%以上,在密碼子使用偏好性方面具有很高的相似性。基于FNSⅡ序列的進化分析與傳統的植物分類結果基本一致,表明FNSⅡ基因與植物的進化過程密切相關。同為忍冬屬的不同種有混合交錯聚為一支的現象,如忍冬與灰氈毛忍冬、黃褐毛忍冬都有聚為一支的現象。其原因除與FNSⅡ序列的高同源性有關外,可能還與該基因影響的木犀草苷在忍冬、灰氈毛忍冬及黃褐毛忍冬中的含量有關,目前在大部分山銀花中未檢測到木犀草苷,但通過超高效液相色譜和高效液相色譜-串聯質譜能在灰氈毛忍冬和黃褐毛忍冬中檢測到少量木犀草苷。以主成分含量為變量對5個忍冬屬種質資源進行的系統聚類分析結果顯示,灰氈毛忍冬和黃褐毛忍冬與忍冬種質差異較其他山銀花種屬小。另外,華南忍冬與非忍冬屬植物歸為一個大類,存在與傳統的植物分類規律沖突的現象。已有研究表明,華南忍冬野生居群具有較高水平的遺傳多樣性,居群間遺傳分化程度較高,地理距離與遺傳距離之間具有顯著相關性,地理隔離效應是導致居群間遺傳分化的重要因素。所以,推測其可能受到生長的地理環境影響,導致華南忍冬的FNSⅡ基因在分化過程中發生了一定程度的變異。
在進化過程中,相同物種或相近種屬會形成相似的密碼子偏好性,密碼子偏好性能在一定程度上反映不同物種間親緣關系的遠近。基于RSCU的聚類分析結果顯示,大部分忍冬屬植物的FNSⅡ基因聚為一支,表明親緣關系較近的物種有相似的密碼子偏好性。少部分忍冬屬植物被聚為其他大類的分支。14個灰氈毛忍冬與4個紅腺忍冬的FNSⅡ基因距離較遠,分別與其他多種植物聚為一支,說明其FNSⅡ基因的密碼子偏好性差異較大。同種植物的FNSⅡ基因被聚類到不同分支上,說明在進化過程中由于突變或自然選擇等壓力導致地域差異,同種屬FNSⅡ基因之間的密碼子偏好性不同。RSCU聚類在一定程度上能夠反映出物種進化的關系,但容易受到更多因素的影響,比較適合在較小的分類單元里提供較為可靠的分類依據,因此,與RSCU聚類相比,基于CDS序列的系統進化樹分析的準確度更高。
通過對比分析忍冬與模式生物的密碼子使用頻率,發現酵母為忍冬的最佳外源表達系統,但為達到較高水平的蛋白表達需要對密碼子進行優化,而擬南芥和煙草均可作為其遺傳轉化受體。本研究可為FNSⅡ基因的分類與演化表達調控及植物遺傳育種等研究提供科學依據,對優化金銀花和山銀花鑒別方法的研究提供指導。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
電子制作(2018年18期)2018-11-14 01:48:24
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
