抗凍蛋白對冷凍蝦夷扇貝閉殼肌持水性及質構特性的影響
2022-06-02 08:42:32史羽瑤王紅麗邱澤慧王錫昌
食品科學 2022年10期
史羽瑤,鄭 堯,王紅麗,邱澤慧,王錫昌
(上海海洋大學食品學院,上海 201306)
蝦夷扇貝()是海水養(yǎng)殖雙殼貝類,是亞洲國家最重要的水產養(yǎng)殖品種之一,廣泛分布于俄羅斯群島、日本北海道和本州島及中國渤海灣附近,具有個體大、生長快、營養(yǎng)價值高等特點,深受國內外消費者的歡迎。據《中國漁業(yè)統(tǒng)計年鑒》,2012—2019年我國扇貝產量基本呈逐年遞增趨勢,2019年扇貝總產量達182.81萬 t,占海水養(yǎng)殖貝類總產量的12.70%。蝦夷扇貝采捕后主要為活品或以閉殼肌為主的加工品(凍品、干制品等)形式流通。活品的經濟價值最高,但存在品質極易劣化、貨架期短等弊端。冷凍通過抑制微生物生長繁殖和酶促反應延長產品貨架期,并在解凍后最大程度保持產品的原有特性。然而,長期凍藏會不可避免引起冰晶生長、蛋白變性及脂質氧化等,最終影響消費者的感官體驗及商品價值。冰晶的生長會對肌纖維造成不可逆的機械損傷,并間接誘導蛋白質變性、脂質氧化,是導致凍藏水產品持水性下降、質構變差的根本原因。
抗凍劑具有降低冰點、抑制冰晶生長、延長凍品貨架期的作用,在水產品中添加抗凍劑是一種有效的凍前品質調控方式。常見的抗凍劑有磷酸鹽類、糖類和蛋白及其衍生物類。磷酸鹽可以提高凍品的持水力,但同時促進了肌動球蛋白解離,加快肌原纖維結構的破壞。且過量磷酸鹽會使凍蝦質地黏稠,呈半透明狀,影響其口感和滋味。海藻糖可以取代蛋白質分子表面的結合水并與之結合,從而抑制蛋白變性;但海藻糖是一種穿透性低溫保護劑,會穿透細胞質,導致核膜收縮,且能量過高,不適于糖尿病患者等人群食用。與普通抗凍劑不同,抗凍蛋白作為最典型的冰活性阻冰劑,是生物為適應極端寒冷環(huán)境、保護生物體免受冷凍損害的特異性糖肽或多肽。抗凍蛋白具備2 大特性:1)熱滯活性:它能夠以非依數的形式降低溶液冰點而不改變其熔點,從而導致溶液的熔點與冰點之間出現差值;2)抑制冰晶生長:抗凍蛋白可通過氫鍵吸附于冰晶表面并積聚在冰-水界面處,利用Kelvin效應抑制胚胎冰晶的形成、生長以及重結晶。抗凍蛋白可以有效降低凍藏過程中冰晶對細胞和組織的損害,即使在低質量濃度下(如0.1 g/100 mL或更低)也有效。與傳統(tǒng)商業(yè)抗凍劑相比,抗凍蛋白具有更好的凍藏穩(wěn)定性,在水產品冷鏈流通過程中有廣泛應用前景。但由于其不易分離純化且分子質量較大,目前針對抗凍蛋白的實用性研究相對較少。
本研究旨在利用真空浸漬的方式將抗凍蛋白添加到蝦夷扇貝閉殼肌中,并與焦磷酸鈉、海藻糖兩種傳統(tǒng)抗凍劑作對比,探究抗凍蛋白相較于傳統(tǒng)抗凍劑對冷凍蝦夷扇貝閉殼肌持水性及質構特性的影響及其機制,以期為冷凍水產品的品質調控提供一定參考。
1 材料與方法
1.1 材料與試劑
蝦夷扇貝由大連獐子島股份有限公司提供,質量(149.14±18.33)g,閉殼肌凈質量(17.36±4.01)g。將鮮活蝦夷扇貝捕撈上岸后,在4 ℃條件下24 h內汽車運輸至上海寶山區(qū)江陽市場,再經2 h冰藏運回上海海洋大學實驗室。快速去除腮、內臟、性腺及外套膜,取出閉殼肌。
抗凍蛋白(純度大于95%,分子質量52 kDa,熱滯值約1.5 ℃) 南京安飛生物有限公司;焦磷酸鈉、氯化鈉、海藻糖、戊二醛固定液、叔丁醇 上海麥克林生化科技有限公司;戴維森固定液、無水乙醇、OCT包埋劑、伊紅染液 北京索萊寶科技有限公司;其他生化試劑均為國產分析純。
1.2 儀器與設備
DW-40L508超低溫冰箱 艾本德(上海)國際貿易有限公司;TA.XT Plus質構儀 英國SMS公司;GM-0.33A隔膜真空泵 天津津騰儀器有限公司;H2050R高速冷凍離心機 長沙湘儀檢測設備有限公司;CR-400型色差計 日本Knoica Minotla Sensing公司;FJ200-SH型數顯高速分散均質機 上海標本模型廠;BT-224S電子分析天平 賽多利斯科學儀器(北京)有限公司;Eclipse E100正置光學顯微鏡 日本Nikon公司;MesoMR23-060H-1型低場核磁共振儀 上海紐邁電子科技有限公司;SU5000熱場發(fā)射掃描電子顯微鏡 日本日立公司。
真空浸漬裝置由真空干燥器、抽濾瓶和真空泵組成,如圖1所示。
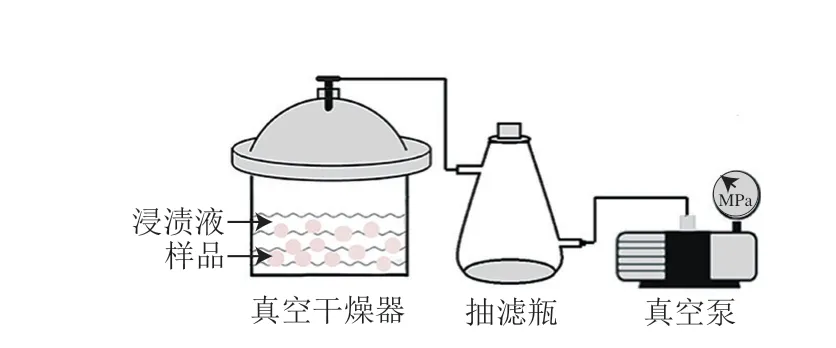
圖1 真空浸漬裝置示意圖Fig.1 Schematic of vacuum impregnation equipment
1.3 方法
1.3.1 樣品前處理
將120 個新鮮閉殼肌隨機分成4 組,按照料液比1∶1.5加入以下浸漬液中:空白組:0.9 g/100 mL NaCl溶液;焦磷酸鈉組:3.0 g/100 mL NaPO溶液+0.9 g/100 mL NaCl溶液;海藻糖組:3.0 g/100 mL海藻糖溶液+0.9 g/100 mL NaCl溶液;抗凍蛋白組:0.2 g/100 mL抗凍蛋白溶液+0.9 g/100 mL NaCl溶液。在4 ℃、0.07 MPa真空度下浸漬1 h,瀝干稱質量,使用鼓風凍結機將其冷凍到-18 ℃后,放入-18 ℃冰箱凍藏7 d,取樣,于4 ℃冰箱解凍3.5 h,使其中心溫度達到0~4 ℃。
1.3.2 得率測定
參考Jiang Qingqing等的方法稱量浸漬前和解凍后的閉殼肌質量,分別記為、,得率即為新鮮閉殼肌質量與冷凍-解凍后閉殼肌質量之比。按式(1)計算得率:

1.3.3 水分含量測定
按照GB 5009.3—2016《食品中水分的測定》中的直接干燥法測定,每組至少重復3 次。
1.3.4 離心損失率的測定
將解凍后的閉殼肌沿肌纖維方向切下質量(2.00±0.02)g樣品,記為/g,用雙層濾紙包裹后放于50 mL離心管底部,以6 000 r/min離心10 min,再次稱樣品質量,記為/g。按式(2)計算離心損失率:
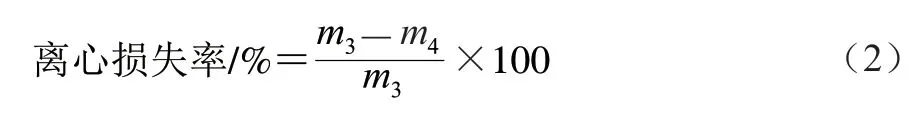
1.3.5 色差測定
參照Tironi等的方法并稍作改動,每組取5 個樣品,采用色差儀測定閉殼肌亮度值(*)、紅度值(*)、黃度值(*)。將色差儀鏡頭垂直緊貼于閉殼肌表面,確保鏡口不漏光。按式(3)計算():
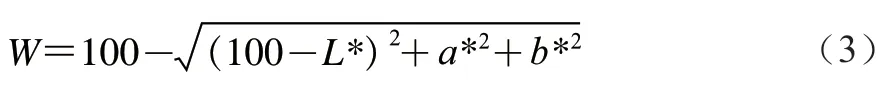
1.3.6 質構特性及剪切力的測定
參考馬海建的方法,略作修改。將貝柱切成20 mm×20 mm×15 mm的長方體,進行2 次軸向壓縮測定。測試條件如下:采用P/50平底柱形探頭,測前速率1.00 m/s,測試速率1.00 mm/s,返回速率1.00 mm/s,壓縮程度50%,兩次壓縮間停留時間5 s,觸發(fā)力5 g,數據采集速率200 points/s。每組樣品測定6 次。
使用WBS探頭,沿垂直于肌纖維的方向將閉殼肌完全切斷,記錄剪切力值(g),每組測量8 次,去掉最大值和最小值,結果取其余數值的平均值。
1.3.7 低場核磁共振及磁共振成像分析
使用低場核磁共振分析儀測定樣品中的水分分布,橫向弛豫時間()采用CPMG脈沖序列進行測定。CPMG得到的指數衰減曲線用儀器自帶軟件進行反演。25 ℃條件下CPMG測試參數:90°脈沖時間18 μs;180°脈沖時間35 μs;時間點數據360 068;前置放大增益1;采樣頻率200 kHz;重復采樣間隔時間3 000 ms;回波時間0.3 ms;回波個數6 000;累加次數8。將整個閉殼肌直接放入直徑 60 mm核磁管中,每組至少取3 個樣品進行測定。
采用自旋回波成像序列獲得樣品加權圖像。所有圖像的視野為80 mm×80 mm,層厚3.0 mm,層間距2.5 mm。重復等待時間500 ms,回波時間18.124 ms,重復采樣8 次,質子密度圖由軟件(上海紐邁電子科技有限公司)統(tǒng)一映射、偽彩化。
1.3.8 冰晶形態(tài)的觀察與測定
使用Luan Lanlan等的方法進行蘇木精-伊紅(hematoxylin-eosin staining,HE)染色確定閉殼肌組織中冰晶形態(tài)的變化。將組織標本包埋在含蒸餾水、福爾馬林、乙酸和乙醇溶液的戴維森固定液中,然后用系列體積分數乙醇溶液對固定樣品脫水。將脫水后的肌肉組織用石蠟包埋,用切片機沿橫向和縱向切5 μm厚的切片。將組織切片進行HE染色,在光鏡下觀察肌肉組織微結構變化。
使用ImageJ圖像分析軟件,參考黃鴻兵等的方法并稍作修改,每組至少選取3 張圖片計算冰晶的平均橫截面積、平均當量直徑、圓度和拉伸度。其中,當量直徑為與冰晶面積相同圓的直徑;圓度值介于0~1,圓度值越大,冰晶越圓;拉伸度為主長軸與次長軸比值,拉伸度為1時,被認為呈圓形或正方形,拉伸度越大,冰晶越長。每張圖中冰晶樣本量均為300以上,以確保數據準確性。
1.3.9 肌纖維微觀結構測定
參考Tabilo-Munizaga等方法,取解凍好的閉殼肌,切成2~3 mm厚的小塊,經2.5%戊二醛溶液于4 ℃固定12 h,用0.2 mol/L pH 7.2磷酸緩沖液漂洗3 次,每次15 min;然后依次用體積分數70%、90%、95%乙醇溶液及無水乙醇進行梯度脫水,脫水時間均為30 min;最后用體積比為3∶1、1∶1、1∶3的乙醇-叔丁醇對樣品進行漂洗,每次10 min。冷凍干燥,鍍膜后置于掃描電子顯微鏡下觀察。
1.4 數據處理
2 結果與分析
2.1 抗凍蛋白對冷凍蝦夷扇貝閉殼肌持水性及質構特性的影響
2.1.1 持水性變化
持水性被定義為肌肉保持自身水分及吸附水分的能力。由表1可知,經過真空浸漬后各組得率均大于100%,表示閉殼肌浸漬后增加的質量均大于解凍損失,說明真空浸漬可以使部分浸漬液截留在閉殼肌內。4 個處理組得率依次為:抗凍蛋白組>焦磷酸鈉組>海藻糖組>空白組,抗凍蛋白組得率較空白組高4.61%。各組水分含量與得率變化趨勢一致,且抗凍蛋白組離心損失顯著低于其他浸漬組(<0.05)。
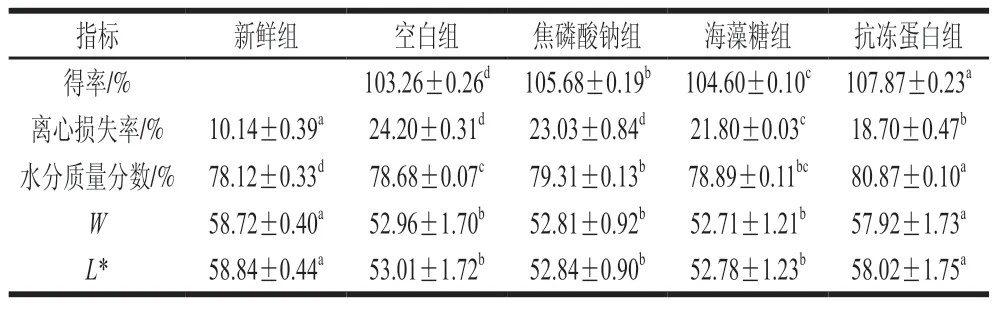
表1 蝦夷扇貝閉殼肌持水性及色澤的變化Table 1 Changes in water-holding capacity and color of scallop adductor muscle
肌肉細胞中高達85%的水分被肌原纖維所占據,因此影響汁液流失的主要原因為肌纖維的收縮程度和細胞外間隙的形成。由此推測,真空浸漬提高得率主要涉及絲狀晶格的膨脹和肌原纖維蛋白的增溶,從而導致組織內部氣體、自由水及可溶性物質等被部分排出,使浸漬液更容易滲透到組織內部。離心損失的降低主要歸因于組織較高的持水性。良好的持水性一方面由于肌原纖維的膨脹與增溶可以持留更多水分,另一方面,可能是由于抗凍劑抑制了冰晶生長,降低可凍結水含量,從而提高了閉殼肌的持水性。
2.1.2 色澤變化
Hughes等認為色澤、持水性與嫩度之間相互影響,肌肉的持水性及結構會影響肉的色澤和光散射特性。由表1可知,冷凍后抗凍蛋白組的*和相較于其他3 個浸漬組顯著升高,且與新鮮組無顯著差異(>0.05),這與速凍豬肉餅凍藏后的色澤變化一致。推測原因可能是由于冷凍過程中冰晶的生長導致肌絲晶格膨脹甚至斷裂,纖維束受損變形,從而使光的散射變弱,光路變長,反射光減少,導致*和顯著降低。因此,色澤改變可能是由于肌絲晶格排列變化影響光的散射,從而導致發(fā)生變化,也可能與組織表面水分增多,持水性降低有關。
2.1.3 質構特性變化
質構是衡量食品結構、評價冷凍水產品品質的重要指標。質構特性主要與肌纖維破壞程度以及蛋白質結構有關,冷凍過程中形成的大且形狀不規(guī)則的冰晶會對細胞膜和結締組織造成機械損傷,從而引起質構劣變。由圖2可知,抗凍蛋白組閉殼肌的硬度、黏聚性與新鮮組幾乎一致,顯著高于焦磷酸鈉、海藻糖組(<0.05),且比空白組分別提高了27.79%和55.47%。經冷凍后,所有浸漬組彈性均顯著下降,且抗凍蛋白組>海藻糖組>焦磷酸鈉組>空白組,抗凍蛋白組閉殼肌的彈性比空白組高10.28%。與鯡魚抗凍蛋白結合CS@FeO納米顆粒對真鯛解凍后質構特性的影響一致。
由圖2可知,新鮮組樣品剪切力顯著高于冷凍后4 組,抗凍蛋白組剪切力高于其他浸漬組(<0.05),且較空白組高16.11%。這與抗凍蛋白作用于鏡鯉后剪切力的變化情況一致。主要原因為凍結過程中冰晶的形成會破壞細胞膜、細胞器,使肌纖維結構部分降解,導致閉殼肌質地變軟,剪切力下降。且肌纖維斷裂程度越明顯,結締組織破壞越嚴重,剪切力就越小。以上結果充分證實了抗凍蛋白組質構特性與新鮮組更為接近,抗凍蛋白對質構特性的改善作用優(yōu)于焦磷酸鈉、海藻糖兩種傳統(tǒng)抗凍劑。
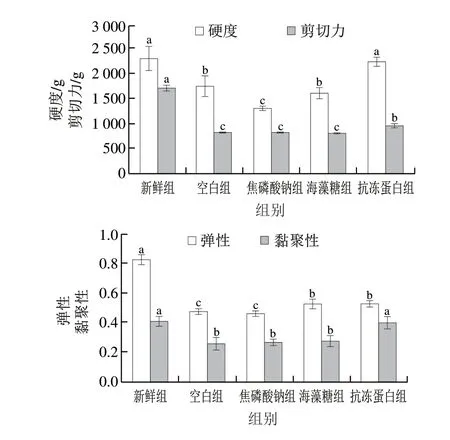
圖2 蝦夷扇貝閉殼肌質構特性的變化Fig.2 Changes in texture properties of scallop adductor muscle
2.2 抗凍蛋白對冷凍蝦夷扇貝閉殼肌水分分布及遷移的影響
如圖3所示,弛豫曲線反映閉殼肌中3 種類型水分,其中(0~10 ms)代表緊密結合大分子極性基團的結合水,(10~100 ms)和(100~1 000 ms)分別對應不易流動水(多為固定在肌原纖維網絡內的水分)和自由水(肌原纖維晶格中的水分)。由表2及圖3可知,冷凍后整體右移,說明細胞外冰晶的形成在一定程度上破壞了細胞膜,導致水分與肌肉組織結合能力變弱。冷凍后各浸漬組無顯著差異(>0.05),抗凍蛋白組顯著小于其他浸漬組(<0.05),原因可能為抗凍蛋白對冰晶形態(tài)的修飾作用減少了由肌纖維變性聚集引起的水分和蛋白質之間的化學交換,減緩了肌原纖維內水分遷移到細胞外。因此,抗凍蛋白組水分與肌原纖維結合更緊密,得率較高,持水性更好。
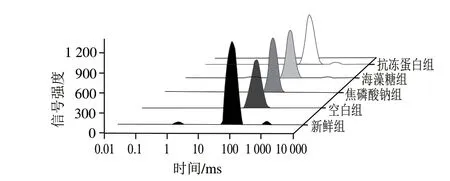
圖3 蝦夷扇貝閉殼肌中不同狀態(tài)水分T2曲線Fig.3 T2 relaxation time curves of different water states in scallop adductor muscle
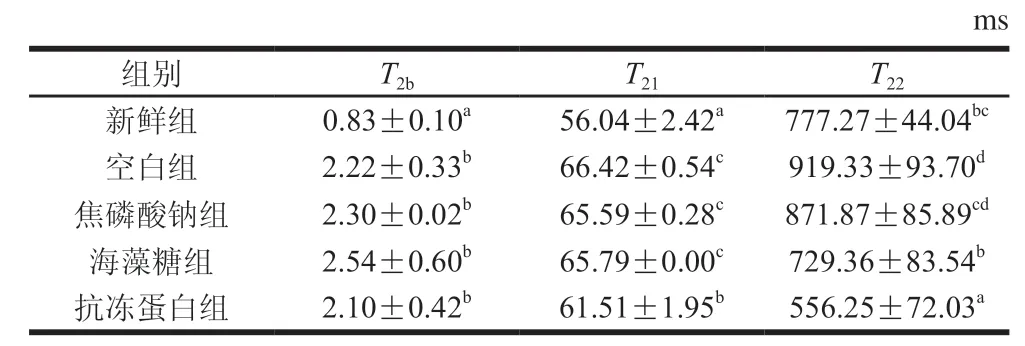
表2 蝦夷扇貝閉殼肌中不同狀態(tài)水分T2分布情況Table 2 Distribution of T2 relaxation time of different water states in scallop adductor muscle
通過質子密度加權成像的方法對蝦夷扇貝閉殼肌的水分分布情況進行可視化處理,其中紅色表示高質子信號密度,藍色表示低質子信號密度,黃色區(qū)域的信號強度則介于紅色和藍色之間。氫質子越密集的區(qū)域信號越強,圖譜越紅,說明該區(qū)域水分含量越多,持水性越好。由圖4可知,冷凍后抗凍蛋白組的質子信號密度與新鮮組幾乎一致,其他浸漬組質子信號密度較低,紅色區(qū)域逐漸被黃色區(qū)域代替。該變化趨勢與表1中各組得率基本相同,進一步證實抗凍蛋白組持水性最好。且冷凍后各組樣品從中心到周邊區(qū)域的紅色程度逐漸降低,邊緣都為黃色和淺藍色,這與牛半膜肌經凍融循環(huán)后觀察到的變化情況相似。可能是由于肌纖維遭到破壞后,水分從肌纖維內部逐漸排出到肌纖維間隙,再通過肌纖維間隙形成的通道從閉殼肌邊緣流出,從而導致邊緣區(qū)域的持水性最差。

圖4 蝦夷扇貝閉殼肌質子密度加權圖Fig.4 Proton density-weighted images of scallop adductor muscle
2.3 抗凍蛋白對冷凍蝦夷扇貝閉殼肌冰晶形態(tài)的影響
冷凍過程中形成的冰晶會影響細胞完整性,而冰晶的大小和形態(tài)決定了冷凍食品品質的劣變程度。通常較大且形狀不規(guī)則的冰晶會不可逆轉地破壞細胞結構,導致持水性、質構特性的劣變和營養(yǎng)物質的損失,而較細小的球形冰晶可以減少對肌纖維的機械損傷。由圖5可知,空白組形成了較大且形狀不規(guī)則的冰柱,焦磷酸鈉和海藻糖組可以在一定程度上改善冰晶形態(tài),但仍會出現明顯的組織間隙和形狀不規(guī)則的冰晶。而抗凍蛋白組的冰晶細小且呈球狀,較均勻地分散在細胞內外。這一現象直觀詮釋了抗凍蛋白組持水性及質構特性較其他組更好的原因。抗凍蛋白側鏈的親水基團通過氫鍵與冰晶棱柱面(冰晶生長表面)相匹配并結合,暴露出高表面自由能的疏水表面,從而降低了冰點,阻礙冰核的形成、冰晶生長以及重結晶。
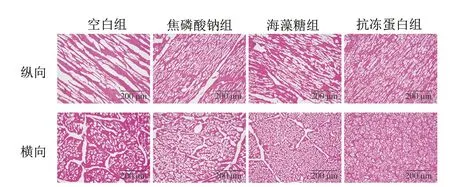
圖5 蝦夷扇貝閉殼肌冰晶形態(tài)的HE染色圖Fig.5 Micrographs of ice crystals in scallop adductor muscle
橫截面積與當量直徑直接反映冰晶表面積的大小,如圖6 A 所示,抗凍蛋白組的平均橫截面積((282.31±19.87)μm)和平均當量直徑((18.95±0.67)μm)最小,其平均橫截面積較空白組減小了65.06%。焦磷酸鈉和海藻糖組對冰晶形態(tài)也具有一定的修飾作用,但效果顯著低于抗凍蛋白組(<0.05)。冰晶圓度和拉伸度反映冰晶形狀的規(guī)則程度,相較于平均橫截面積和當量直徑的變化較小。如圖6B所示,3 種抗凍劑浸漬組的冰晶圓度無顯著差異(>0.05),但抗凍蛋白組的拉伸度低于其他浸漬組。進一步證實了抗凍蛋白對冰晶形態(tài)具有良好的修飾作用,從而減少了冰晶對肌纖維的擠壓和破壞,增強細胞完整性,提高冷凍后蝦夷扇貝閉殼肌的持水性及質構特性。
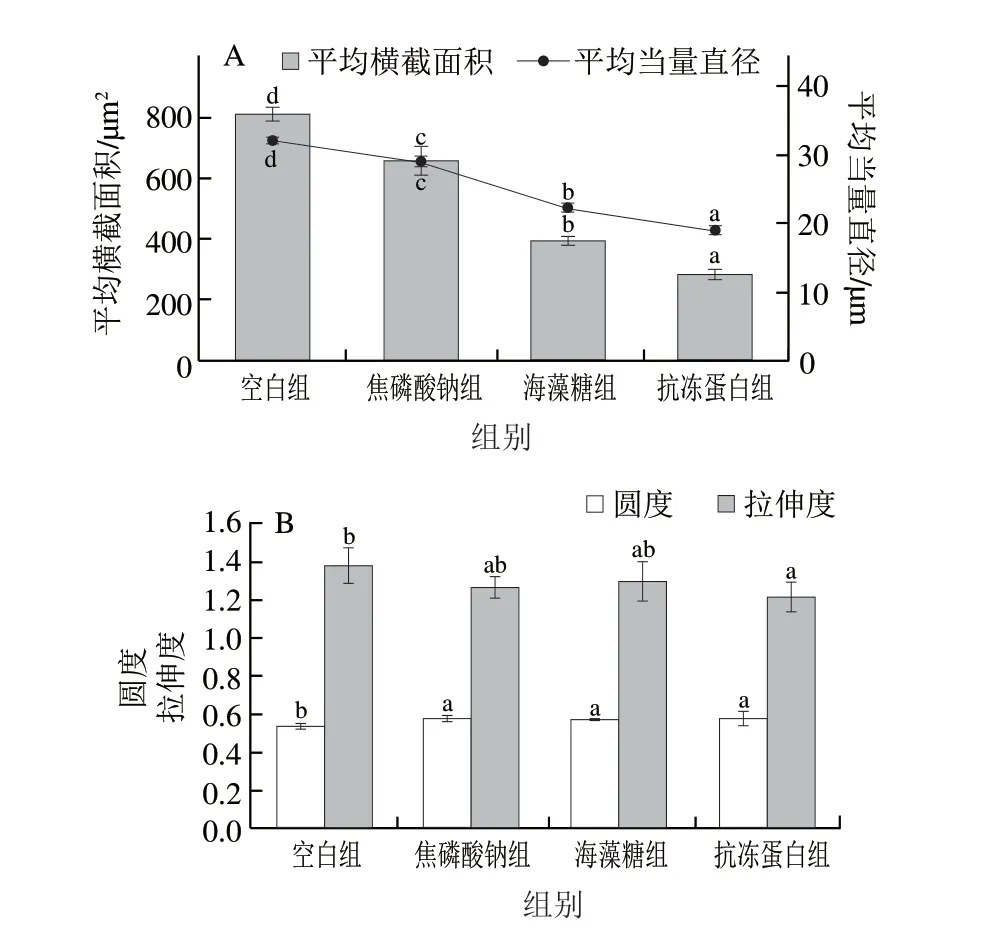
圖6 蝦夷扇貝閉殼肌的冰晶形態(tài)分析Fig.6 Analysis of ice crystal morphology in scallop adductor muscle
2.4 抗凍蛋白對冷凍蝦夷扇貝閉殼肌組織結構的影響
肌肉持水性和質構特性與其組織結構密切相關。如圖7所示,新鮮組纖維束排列有序,相互平行,具有清晰的網絡結構。而空白組肌纖維受損變形,存在較大孔洞和嚴重的斷裂情況。冰晶生長導致肌纖維斷裂,從而使組織內部產生了不可逆的形變。相較于空白組,焦磷酸鈉、海藻糖組肌纖維束相對整齊,纖維束間孔洞較小且形狀較規(guī)則,說明焦磷酸鈉和海藻糖一定程度上修飾了冰晶形態(tài),減少了冰晶對肌纖維的機械損傷程度。而抗凍蛋白組肌纖維排列仍清晰可見,纖維束緊密堆積,破壞程度最小,推測原因也可能為抗凍蛋白結合到冰晶表面后積聚在冰-水界面處,使冰晶曲面和邊緣表面積增大,蒸汽壓力升高,體系的平衡狀態(tài)發(fā)生改變,從而導致冰點降低,即Kelvin效應。因此抗凍蛋白對冰晶形態(tài)的修飾作用最佳,抗凍蛋白組硬度、黏聚性與新鮮組幾乎一致,持水性較其他組顯著提高。
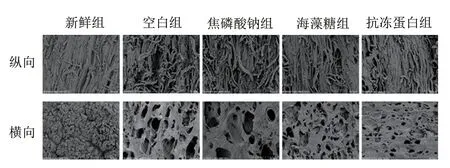
圖7 蝦夷扇貝閉殼肌組織微觀結構變化(×800)Fig.7 Changes in microstructural structure of scallop adductor muscle (× 800)
3 結論
抗凍蛋白有效改善了冷凍蝦夷扇貝閉殼肌的持水性及質構特性,且效果顯著優(yōu)于焦磷酸鈉和海藻糖。主要表現為抗凍蛋白組硬度、黏聚性及質子密度與新鮮組幾乎一致,且顯著高于焦磷酸鈉和海藻糖組。而相比于其他浸漬組,抗凍蛋白組得率、彈性及剪切力顯著增加,顯著縮短(<0.05),冰晶細小且呈球狀,肌纖維結構較清晰完整,且冰晶平均橫截面積比空白組減少了65.06%。本研究證實了抗凍蛋白具有吸附于冰晶表面從而抑制冰晶生長、修飾冰晶形態(tài)的作用,通過真空浸漬處理后對冷凍蝦夷扇貝閉殼肌持水性及質構特性的調控作用優(yōu)于焦磷酸鈉、海藻糖,是一種有效的冷凍水產品抗凍劑,為進一步探究抗凍蛋白在冷凍過程中對蛋白變性的影響及抗凍機制提供參考。
