釀酒酵母表達側耳源單基因生物合成麥角硫因
2022-06-02 08:43:00林金德余穎豪郭麗瓊林俊芳
食品科學 2022年10期
關鍵詞:生物
潘 濤,林金德,余穎豪,郭麗瓊,2,林俊芳,2,
(1.華南農業大學食品學院,廣東 廣州 510642;2.廣東省微生態制劑工程技術研究中心,廣東 廣州 510642)
麥角硫因(ergothioneine,EGT),又稱2-巰基組氨酸-三甲基甜菜堿,最初由Chairles Tanret于1909年在麥角菌()中發現,是機體內重要的生物活性物質,廣泛存在于生物體內,具有清除自由基、解毒、保護DNA免受金屬離子的氧化損傷、維持細胞的正常生長和細胞免疫以及防止紫外線輻射損傷等多種生理功能。EGT是一種重要的天然抗氧化劑,有抗衰老、抗氧化、抗炎癥等功能,與神經退行性疾病、抑郁癥等精神疾病存在一定的關聯,與人類健康息息相關,但人體自身不能合成EGT,只能通過膳食攝入。EGT在制藥、生物醫學、化妝品和食品等領域展現出良好的應用前景。
生物合成法新近發展起來的利用多基因組合表達的高效生物制備方法,可以通過代謝調控等發酵過程控制手段有效地通過異源合成提高EGT產率。近年來,科學家們對EGT合成途徑的研究發現恥垢分枝桿菌EGT合成途徑有5 個合成酶基因(~),但是分枝桿菌等條件致病菌的EGT基因來源存在一定的安全隱患,同時基因數目較多等導致產業化生產的難度較大。后來有研究者發現粗糙脈孢菌()EGT生物合成途徑有2 個基因,即和,其中中的C—S裂解過程已被解析。Pluskal等從粟酒裂殖酵母()中挖掘出1 個與同源性較高的基因和4 個與C—S鍵斷裂相關()的基因,通過基因敲除驗證前者參與EGT的合成,而后者4 個基因中只有1 個參與了EGT的合成。本課題組在食用菌生物合成EGT方面已取得一定的研究成果并克隆得到金針菇和灰樹花的EGT合成途徑基因,在金針菇中有3 個基因(、和),在灰樹花中有兩個基因(和)。相比于恥垢分枝桿菌等原核細菌,真菌具有更加簡潔的EGT生物合成途徑,為EGT的代謝工程大規模化生產提供了便捷的手段。目前,EGT相關合成酶應用于EGT的生產相對較少,因此,進一步挖掘EGT合成酶基因將有利于尋找催化活性更好的EGT合成酶,從而提高EGT的產量,推動EGT產業的發展。
大多數側耳屬食用菌可以產EGT,但其相關合成酶尚未被挖掘和通過代謝工程應用于EGT的生產。故本實驗擬結合生物信息學分析等方法,挖掘、克隆側耳屬EGT相關合成酶基因,并進行功能鑒定,致力于尋找更加高效的EGT合成酶,促進我國EGT產業的發展。
1 材料與方法
1.1 材料與試劑
1.1.1 菌株與質粒
平菇()、白靈菇()、杏鮑菇()購自山東省壽光市食用菌研究所;大腸桿菌DH5a、釀酒酵母EC 1118、pYES 2載體、釀酒酵母表達載體pRS42K均由本實驗室保存;pCloneEZ-TOPO克隆載體購自上海唯地生物技術有限公司。
1.1.2 試劑與培養基
酶、PrimerSTAR Max Premix(2×)、限制性核酸內切酶R I 寶日醫生物技術(北京)有限公司;質粒小量提取試劑盒、瓊脂糖凝膠DNA回收試劑盒等南京諾唯贊生物科技股份有限公司;反轉錄試劑盒Script One-Step gDNA Removal and cDNA Synthesis SuperMix 北京全式金生物公司;引物和DNA測序由廣州天一輝遠基因科技有限公司完成。
LB(Luria Broth)培養基:胰蛋白胨10 g/L,酵母提取粉5 g/L,NaCl 10 g/L(固體培養基則加入2%瓊脂粉),121 ℃高壓滅菌30 min,抗性篩選時加入氨芐青霉素使其最終質量濃度為100 mg/L。酵母浸出粉胨葡萄糖(yeast extract peptone dextrose,YPD)培養基:酵母提取粉10 g/L,蛋白胨20 g/L,葡萄糖20 g/L(固體培養基則加入2%瓊脂粉),115 ℃高壓滅菌15 min,抗性篩選時加入G 418硫酸鹽使其最終質量濃度為100 mg/L。馬鈴薯葡萄糖(potato dextrose broth,PDB)培養基:取去皮土豆200 g的沸水提取物(煮沸30 min至土豆變軟),稱取20 g葡萄糖、1.5 g MgSO·7HO、3 g KHPO,加蒸餾水定容至1 000 mL,自然pH值,121 ℃滅菌30 min。
1.2 儀器與設備
LC-2030高效液相色譜(high performance liquid chromatography,HPLC)儀 日本株式會社島津制作所;電擊轉化儀 美國Bio-Rad公司;JY92-2D超聲波破碎儀 寧波新芝生物科技股份有限公司;YX-280D手提式壓力蒸汽滅菌鍋 合肥市華泰醫療設備有限公司。
1.3 方法
1.3.1 側耳屬EGT合成途徑基因序列的挖掘
利用已報道的粗糙脈孢菌EGT生物合成酶(NCU04343)、金針菇、灰樹花通過NCBI數據庫BLASTp和BLASTn,比對搜索側耳屬食用菌平菇、白靈菇和杏鮑菇的假定EGT合成酶基因,分別命名為、和。
1.3.2 側耳屬基因的克隆
將平菇、白靈菇和杏鮑菇接種于PDB培養基中進行振蕩培養,取新鮮的菌絲球用STE法進行總RNA的提取。采用全式金公司的Script One-Step gDNA Removal and cDNA Synthesis SuperMix反轉錄試劑盒將總RNA反轉錄成cDNA。設計引物,以反轉錄產物cDNA為模板,采用高保真酶PrimerSTAR Max Premix(2×)進行聚合酶鏈式反應(polymerase chain reaction,PCR)擴增,擴增產物回收純化后,按pCloneEZ-TOPO克隆載體試劑盒方法連接到TOPO載體,轉化至大腸桿菌DH5a感受態細胞中,挑取轉化子進行菌落PCR鑒定,將鑒定正確的轉化子送往廣州天一輝遠基因科技有限公司測序。克隆側耳屬基因所用引物如表1所示。
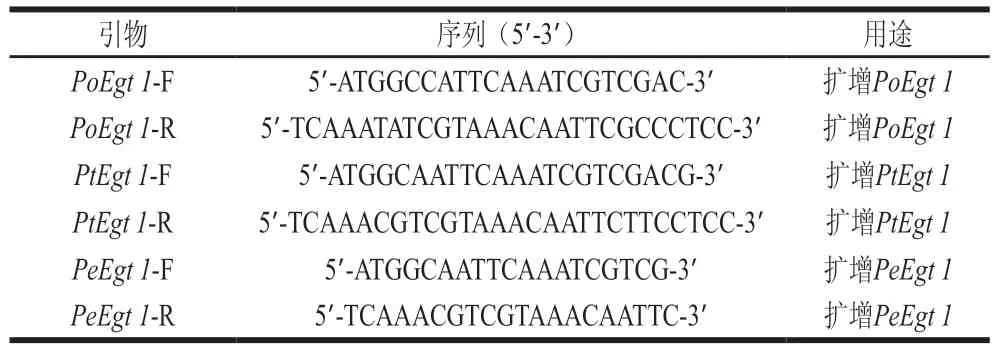
表1 Egt 1克隆引物Table 1 Primer sequences used for Egt 1 cloning
1.3.3 側耳屬的生物信息學分析
將側耳屬Egt 1蛋白氨基酸序列上傳至ExPASProtParam tool(http://web.expasy.org/protparam/),對其進行理化性質分析;再將氨基酸序列上傳至SWISS MODEL(https://swissmodel.expasy.org/)中,通過同源模板比對建模,進行蛋白質三級結構預測。
1.3.4 表達載體pRS42K---構建
以pRS42K和pYES 2質粒為模板,用高保真酶PrimerSTAR Max Premix(2×)擴增出帶pRS42K同源臂的啟動子和終止子,以測序正確的基因pCloneEZ-TOPO克隆載體質粒為模板,擴增基因并在上下游加上啟動子和終止子的同源臂,通過重疊延伸組建成一個表達框--,采用Clone ExpressII One Step Cloning Kit同源重組試劑盒將加同源臂的--表達框重組到線性化的pRS42K載體上并轉化至大腸桿菌DH5α感受態細胞中,挑取轉化子進行菌落PCR鑒定,將鑒定正確的轉化子送往廣州天一輝遠基因科技有限公司測序。
1.3.5 釀酒酵母EC 1118感受態的制備、轉化和鑒定
大量提取測序正確的pRS42K---大腸桿菌轉化子質粒,并轉化至釀酒酵母EC1118中。釀酒酵母感受態的制備和電轉化參考Zhang Yueping等的方法進行,轉化結束后將菌液涂布于含100 μg/mL G 418的YPD平板,30 ℃培養2 d,挑取轉化子進行菌落PCR鑒定。
1.3.6 釀酒酵母的發酵培養
種子液的制備:將成功構建的釀酒酵母工程菌株接種于50 mL含100 μg/mL G418的YPD液體培養基中,30 ℃、200 r/min過夜預培養。按照1%的接種量將構建成功的釀酒酵母工程菌液種子液接種到50 mL含100 μg/mL G418的YPD液體培養基中,30 ℃、220 r/min培養16 h。
1.3.7 釀酒酵母中平菇PoEgt 1重組酶體外酶活性測定
發酵結束后用磷酸鹽緩沖液(phosphate buffered saline,PBS)(pH 7.4)分別洗滌收集細胞,將平菇的工程菌株混勻在10 mL PBS中,加入反應底物以控制最終濃度為:0.02 mol/L組氨酸、0.02 mol/L半胱氨酸、0.005 mol/L-腺苷蛋氨酸、0.01 mol/L Fe、0.005 mol/L磷酸吡哆醛。混勻,超聲破碎菌體,破碎條件:功率33%,約250 W;總超聲時間6 min,工作3 s,間隔3 s,報警溫度40 ℃。超聲結束后,將反應體系放置在36 ℃反應2 h,期間每隔20 min混勻一次。反應產物利用0.22 μm水系濾膜過濾后用于HPLC檢測。
HPLC測定條件:色譜柱采用月旭(Welch)公司的UltimateHILIC Amphion II(4.6 mm×150 mm,5 μm),檢測波長257 nm,流速1 mL/min,流動相(乙腈∶水=80∶20),單次樣品分析采集時間30 min,對照組設置為釀酒酵母EC 1118正常菌株。利用1.5、3、6、12 mg/L的EGT標準品溶液進行EGT標準曲線的制作。
1.4 數據處理
發酵實驗做3 次平行,測定EGT含量取平均值,利用Excel 2019軟件進行數據處理;核酸及氨基酸序列同源性比對采用DNA MAN 8軟件。
2 結果與分析
2.1 側耳屬Egt 1基因的挖掘分析和克隆
利用粗糙脈孢菌、金針菇和灰樹花核酸序列及氨基酸序列,運用BLASTp和BLASTn進行比對得到平菇中的相似性較高的蛋白假定蛋白PLEOSDRAFT_1058188(KDQ26018.1)、杏鮑菇中相似性較高的蛋白假定蛋白BDN71DRAFT_1511997(KAF9489594.1)和白靈菇基因組中相似度較高的核酸序列(序列ID為MKZX01000041.1,區域為78 985~82 124)。以cDNA為的編碼序列擴增模板進行PCR擴增,擴增產物切膠回收后經1%瓊脂糖凝膠電泳檢測結果如圖1所示,平菇、杏鮑菇和白靈菇條帶大小約2 600 bp,片段大小符合。
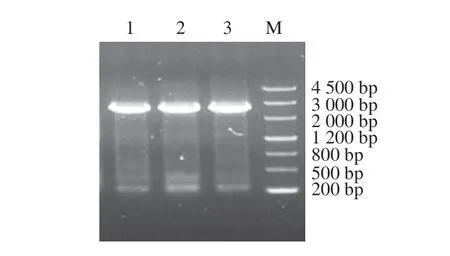
圖1 Egt 1擴增凝膠電泳圖Fig.1 Identification of Egt 1 by PCR
2.2 側耳屬Egt 1的生物信息學分析
2.2.1 側耳屬Egt 1蛋白的理化性質分析
通過ExPAS-ProtParam tool(http://web.expasy.org/protparam/)分別對所編碼蛋白質進行理化分析得到結果如下:PoEgt 1、PtEgt 1、PeEgt 1蛋白大小分別為96.9、96.7、96.8 kDa,依次由853、855、854 個氨基酸組成,等電點pI分別為5.26、5.20、5.20,總親水性的平均值依次為-0.509、-0.485、-0.455。親疏水性中正值越大表示越疏水,負值越大表示越親水,根據總親水性的平均值分析結果可以看出側耳屬蛋白為親水性蛋白。
2.2.2 側耳屬同源性比較
利用DNA MAN 8軟件比較克隆后測序得到的核酸序列和蛋白氨基酸序列(氨基酸序列比對結果如圖2所示),根據比對結果可知,從cDNA中克隆得到的杏鮑菇、平菇和白靈菇核酸序列同源性為97.03%,蛋白氨基酸序列同源性為97.93%。三者的核酸序列和氨基酸序列相似度極高,后續驗證活性實驗選取平菇進行。

圖2 PoEgt 1、PeEgt 1、PtEgt 1氨基酸比對結果Fig.2 Alignment of amino acid sequences of PoEgt 1,PeEgt 1 and PtEgt 1
2.2.3 側耳屬Egt 1的結構及功能預測
利用Swiss-Model對Egt 1蛋白進行預測分析,分析結果顯示,側耳屬Egt 1蛋白氨基酸序列能明顯檢測到與分枝桿菌中和的相似性。進一步利用Swiss-Model對側耳屬Egt 1進行三級結構預測,預測結果如圖3所示,3 個蛋白的預測三維結構相似。再利用NCBI-Conserved Domains Search database對側耳屬Egt 1蛋白氨基酸序列進行保守結構域預測分析發現,側耳屬Egt 1可能含有2 個結構域,分別為甲基轉移酶超家族結構域和硫酸酯酶超家族結構域。其中,甲基轉移酶結構域被描述為組氨酸特異性甲基轉移酶,擁有-腺苷甲硫氨酸依賴性,由分枝桿菌基因簇的一部分表達,即基因,可將3 個甲基轉移至組氨酸的-氨基部分;硫酸酯酶超家族結構域對應Egt 1的后半部分,描述為硫酸酯酶修飾因子酶,該結構域存在于翻譯后硫酸酯酶修飾SUMF1蛋白所需的真核蛋白中,其中基因編碼甲酰甘氨酸生成酶,是一種硫酸酯酶,可能將半胱氨酸氧化成半胱氨酸亞硫磺,再催化成甲酰甘氨酸。
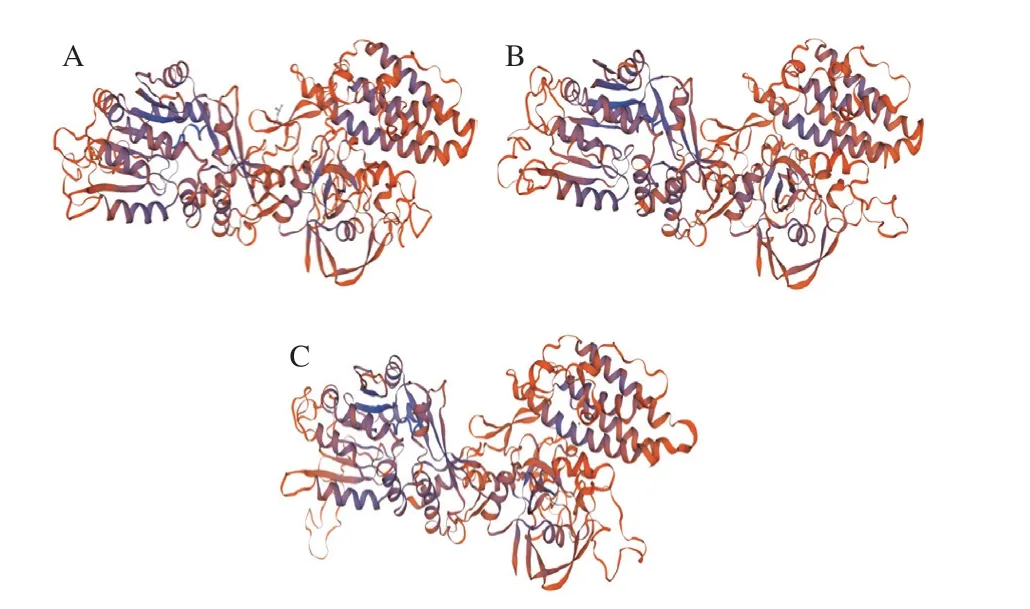
圖3 PoEgt 1(A)、PeEgt 1(B)和PtEgt 1(C)三維結構圖Fig.3 Structural diagram of PoEgt 1 (A),PeEgt 1 (B) and PtEgt 1 (C)
2.3 釀酒酵母PoEgt 1表達載體的構建
進行表達載體pRS42K---構建及轉化,通過OEPCR得到的表達框--與線性化的pRS42K進行重組后轉化至大腸桿菌DH5α,菌落PCR鑒定后送測序,測序結果正確,表達載體pRS42K--構建成功,提取質粒轉化進釀酒酵母EC 1118中。
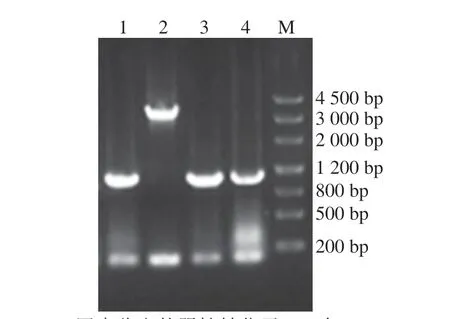
圖4 釀酒酵母轉化菌株pRS42K-TEF1p-PoEgt 1-CYC1t PCR鑒定Fig.4 PCR identification of S.cerevisiae transformants for strain pRS42K-TEF1p-PoEgt 1-CYC1t
2.4 釀酒酵母工程菌株EC 1118-42K-TEF1p-PoEgt 1-CYC1t發酵產物的檢測
EGT標準曲線方程為=76 922-4 570.4,式中,為EGT質量濃度/(mg/L),為峰面積;相關系數=0.999 8,說明EGT在1.5~12 mg/L之間線性關系良好。
含表達質粒pRS42K---的酵母發酵培養后,破碎菌體酶促反應產物出現與標準品EGT出峰時間相同的吸收峰(圖5),表明酶促反應體系中有EGT生成,產量為(2.5±0.08)mg/L。而釀酒酵母對照菌株EC 1118(培養時不加G 418抗生素)相同處理后HPLC檢測沒有出峰,即沒有EGT生成,表明具有單基因生物合成EGT的功能。
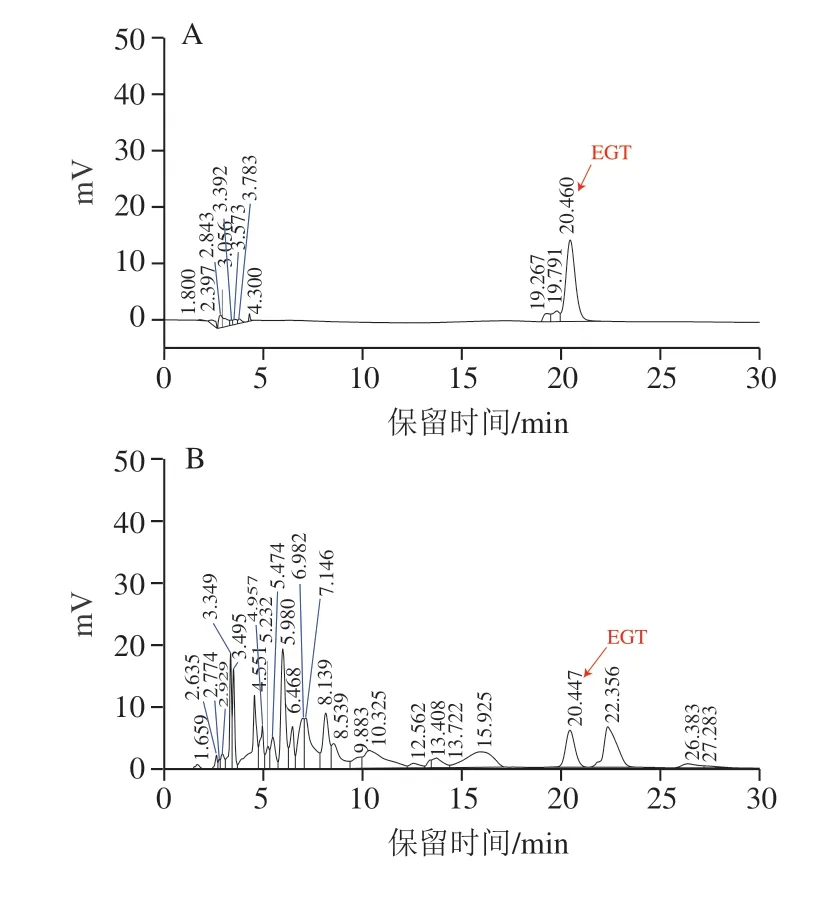
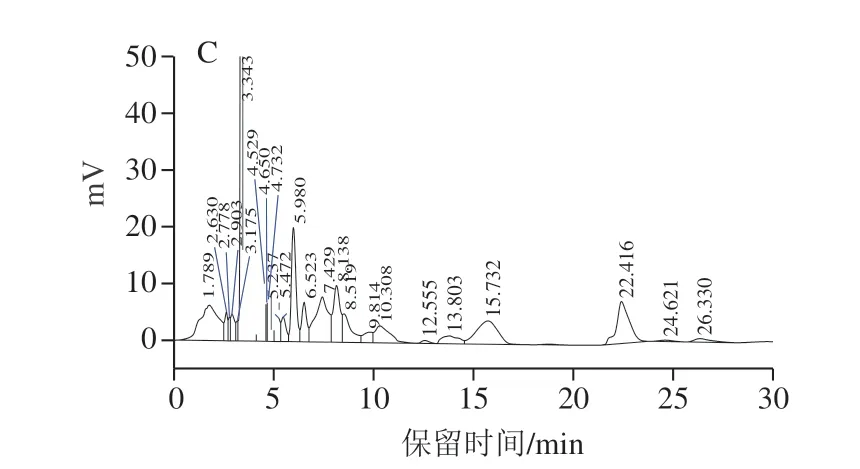
圖5 EGT HPLC檢測結果Fig.5 HPLC profiles of EGT from different sources
3 討論
自1909年EGT被發現以來已有100多年,但是直到21世紀初才通過體外表征得出恥垢分枝桿菌EGT由五基因簇負責合成,其合成途徑繁瑣、基因數目過多導致異源表達時載體構建麻煩,同時過大的載體也會對菌體造成負擔。后來發現真核生物粗糙脈孢菌、粟酒裂殖酵母等也能合成EGT,體內生物合成途徑有2 個酶(Egt 1和Egt 2),這大大簡化了EGT的生物合成途徑,推動了異源合成EGT的進展,并證明了挖掘EGT合成酶基因新來源的重要性。
食藥用菌中含有豐富的EGT,但目前對于食藥用菌中EGT相關合成基因的研究鮮有報道。在灰樹花中,甲基化形成的組氨酸甜菜在介導下可以與半胱氨酸反應形成C—S鍵,隨后經過裂解C—S鍵脫去半胱氨酸生成EGT。在金針菇中,可催化組氨酸和半胱氨酸形成組氨酸甜菜堿半胱氨酸亞砜再在和的共同作用下將組氨酸甜菜堿半胱氨酸亞砜轉化為EGT,而第一步在分枝桿菌中需要-的共同參與,合成途徑繁瑣。在粗糙脈孢菌中,只能催化組氨酸甜菜堿生成組氨酸甜菜堿半胱氨酸亞砜,而組氨酸如何轉化為組氨酸甜菜堿還尚不清楚。本研究采用生物信息學分析軟件和數據庫進行側耳屬蘑菇EGT合成途徑基因的挖掘、蛋白結構和功能的預測,克隆得到側耳屬EGT合成途徑基因、和,在釀酒酵母中進行異源表達并檢測到產物EGT,產量為(2.5±0.08)mg/L,證明了該基因具有單基因合成EGT的活性,豐富了EGT合成酶基因來源。進一步通過建模、功能預測等發現側耳屬Egt 1是由2 個功能結構域組成的酶,分別對應分枝桿菌中和的功能,主要介導組氨酸甲基化的形成和半胱氨酸C—S鍵的形成。許多大腸桿菌和釀酒酵母中本身就含有半胱氨酸脫硫酶,所以只需向含有半胱氨酸脫硫酶的工程菌中轉入便可以獲得EGT,這與本課題組前期關于灰樹花的研究一致,還有待進一步挖掘、研究側耳屬EGT合成途徑中的其他基因以及通過分子手段(如基因編輯技術等)對EGT代謝通路進行改造,更換啟動子、運用整合型表達載體將EGT合成酶基因整合進酵母基因組中,提高其在異源表達宿主中的穩定性,再進一步通過發酵優化等手段提高EGT產量。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
