基于高通量測序技術分析恩施州淡豆豉的細菌多樣性
2022-06-02 08:42:50馬上超莫開菊汪興平
食品科學 2022年10期
馬上超,程 超,田 成,陳 業,莫開菊,汪興平,
(1.湖北民族大學生物科學與技術學院,湖北 恩施 445000;2.生物資源保護與利用湖北省重點實驗室(湖北民族大學),湖北 恩施 445000)
豆豉是以大豆為主要原料,經過浸泡蒸煮,自然發酵或者純菌種接種發酵而成的傳統豆制品[1]。根據發酵菌種不同,可分為曲霉型、毛霉型、根霉型和細菌型4 種[2];根據生產工藝不同,可分為水豆豉、凍豆豉、霉豆豉、干豆豉。大豆經微生物發酵后,產生獨特的豆豉風味,同時產生黃酮、低聚肽、皂苷、豆豉纖溶酶等活性物質[3],具有一定的藥用價值,對消化不良、煩躁胸悶、焦躁不安有一定的緩解作用。日本納豆富含納豆激酶,通過長期深入的優勢菌種篩選和優化,培養獲得高產納豆激酶的菌種,納豆及其健康產品得到長足發展,而我國傳統豆豉營養豐富、種類多樣,但離產業化、健康化差距較大。因此,我國科技工作者亟需開展豆豉微生物菌群的多樣性研究,從眾多優勢菌群中篩選具有醫療保健價值的菌株,促進傳統豆豉的現代化發展。
高通量測序技術能更全面、精確地測定生物環境中各類微生物的相對豐度[4],已廣泛用于分析各種發酵產品的微生物群落結構[5]。李世瑞等[6]采用454高通量焦磷酸測序技術發現曲霉型豆豉發酵中厚壁菌門(Firmicutes)、變形菌門(Proteobacteria)、放線菌門(Actinobacteria)為優勢細菌門。王強等[7]采用Illumina MiSeq高通量測序技術對湖南省龍山縣傳統發酵豆豉細菌多樣性進行研究,結果表明,其優勢細菌門為厚壁菌門,相對豐度為7.43%,優勢細菌屬芽孢桿菌屬(Bacillus)和魏斯氏菌屬(Weissella),相對豐度分別為21.46%和15.83%。Chen Yuhang等[8]研究中國西南地區水豆豉微生物多樣性,發現其優勢菌門為厚壁菌門、擬桿菌門和變形菌門,主要優勢細菌屬為芽孢桿菌屬、擬桿菌屬(Bacteroides)和乳酸桿菌屬(Lactobacillus)。龐春霞等[9]采用Illumina MiSeq高通量測序技術分析納豆、豆豉、大醬、霉豆渣、天貝的細菌多樣性,其中相對豐度大于10%的主要優勢菌屬為芽孢桿菌屬、枝芽孢菌屬(Virgibacillus)、變形桿菌屬(Proteus)、依格納季氏菌屬(Ignatzschineria)、四聯球菌屬(Tetragenococcus)、假單胞菌屬(Pseudomonas)、不動桿菌屬(Acinetobacter)、解脲芽孢桿菌屬(Ureibacillus)、泛菌屬(Pantoea)和片球菌屬(Pediococcus)。通過以上研究發現,豆豉種類、產地不同,主要的優勢菌也有差異。
我國生產制作細菌型豆豉的地區很多,如云南、湖南、貴州、山東、湖北等地。湖北恩施山區氣候宜人,獨特的飲食文化孕育了種類繁多的豆豉產品,恩施傳統豆豉以作坊式生產為主,生產環境條件相對粗放且制作微環境差異大,但目前缺乏對恩施豆豉的微生物菌群特征的系統研究。因此本實驗收集恩施州不同產地5 種細菌型淡豆豉,利用Illumina PE250高通量測序技術對16S rRNA V3~V4區進行測序,分析恩施地區豆豉發酵微生物菌群結構,初步鑒定其優勢菌群,以期推進恩施豆豉的現代化生產,為提高恩施地區豆豉質量和深度開發保健性、安全性產品奠定理論基礎。
1 材料與方法
1.1 材料與試劑
樣品來源于湖北民族大學周邊2 km內及利川市關東村5 個不同商家制作的傳統自然發酵細菌型淡豆豉,制作時間均為12月初。選取5 種未經后加工的細菌型淡豆豉毛坯,使用已紫外滅菌的勺子取發酵壇內部豆豉,現場分裝于滅菌離心管,液氮冷凍運回實驗室,存放于-80 ℃冰箱備用,使用15 kg干冰進行樣品寄送。樣品編號分別為S1、S2、S3、S4、S5。
D3141 HiPure Stool DNA Kits 廣州美基生物科技有限公司;無水乙醇 廣州化學試劑廠;瓊脂糖 西班牙Biowest公司;GoldView I型核酸染色劑 武漢富鑫遠科技有限公司;聚合酶鏈式反應(polymerase chain reaction,PCR)用試劑 美國New England Biolabs公司;AMPure XP磁珠 美國貝克曼庫爾特公司。
1.2 儀器與設備
5427R離心機 德國艾本德股份公司;TM-D超純水儀器 上海樂楓生物科技有限公司;DWHL528S-80 ℃冰箱 中科美菱低溫科技股份有限公司;Mix-28+渦旋振蕩器 廣州圍谷潤儀器有限公司;NanoDrop 2000微量分光光度計、Qubit 3.0熒光計 美國賽默飛世爾科技公司;DYY-6C瓊脂糖凝膠電泳儀 北京六一儀器廠;Tanon-2500凝膠成像系統 上海天能科技有限公司;ETC811 PCR儀 蘇州東勝興業科學儀器有限公司。
1.3 方法
1.3.1 豆豉樣品中總DNA的提取和鑒定
取-80 ℃中保存的S1、S2、S3、S4、S5樣品,充分破碎攪拌,取200 mg豆豉樣品于離心管中,按照R Stool DNA試劑盒說明書提取總DNA。對提取的基因組DNA產物用2%瓊脂糖凝膠電泳進行檢測,目的條帶大小正確,濃度合適,將檢測合格樣品進行高通量測序,每組做4 次生物學重復。
1.3.2 豆豉16S rRNA高通量測序
以豆豉總DNA為模板進行細菌16S rRNA V3~V4區域擴增。引物:341F(CCTACGGGNGGCWGCAG)和806R(GGACTACHVGGGTATCTAAT)。PCR擴增體系和程序參照Guo Mengjiao等[10]的方法并略作改動。第1輪PCR擴增體系:模板100 ng,2 mmol/L dNTPs 5 μL,10×Buffer KOD 5 μL,10 μmol/L正向引物1.5 μL,10 μmol/L反向引物1.5 μL,25 mmol/L MgSO43 μL,KOD 1 μL,無菌雙蒸水33 μL。第1輪PCR擴增程序:94 ℃預變性2 min,98 ℃變性10 s,55 ℃退火30 s,68 ℃延伸30 s,共循環30 次;68 ℃再延伸5 min。利用AMPure X磁珠純化第1輪PCR產物,純化后用Qubit 3.0熒光計定量。第2輪PCR擴增體系:模板100 ng,2 mmol/L dNTPs 5 μL,10×Buffer KOD 5 μL,25 mmol/L MgSO43 μL,10 μmol/L索引引物1 μL,10 μmol/L通用PCR引物1 μL,KOD 1 μL,無菌雙蒸水34 μL。第2輪PCR擴增程序:94 ℃預變性2 min,98 ℃變性10 s,65 ℃退火30 s,68 ℃延伸30 s,共循環12 次,68 ℃再延伸5 min。使用AMPure XP磁珠純化第2輪擴增產物,用StepOnePlus Real-Time PCR System進行定量,根據Novaseq 6000 PE250進行混合測序。
1.4 數據分析
使用Illumina PE250對高通量測序數據進行操作分類單元(operational taxonomic units,OTU)聚類分析、物種注釋、α多樣性分析、β多樣性分析[11-15];使用R語言工具進行主成分分析(principal components analysis,PCA)、非加權組平均法(unweighted pair-group method with arithmetic means,UPGMA)聚類樹、物種間相關系數計算和檢驗;使用R語言工具進行稀釋曲線、Shannon曲線、Simpson曲線、群落熱圖繪制;使用Omicsmart平臺(http://www.omicsmart.com)進行生物信息學分析。
2 結果與分析
2.1 細菌多樣性分析
由圖1可知,隨著測序量增大,測序檢出的OTU數量先增加后逐漸趨于平緩,表明繼續延長測序時間沒有更多物種被檢出,說明本次測序數據量合理,能夠較好反映出20 例恩施州豆豉樣品中的菌群信息。
從20 例恩施細菌型淡豆豉中共得到2 528 835 條序列,篩選出2 380 484 條高質量序列,平均每個樣品119 024 條高質量序列(范圍:107 609~131 494 條,標準偏差:6 705 條),平均序列有效性達92.5%。對這些序列進行分析,共得到3 901 個OTU(范圍:90~264 個,標準偏差:52 個),經對比匹配,這些OTU可以劃分為17 門、24 綱、43 目、78 科、152 屬以及93 種。
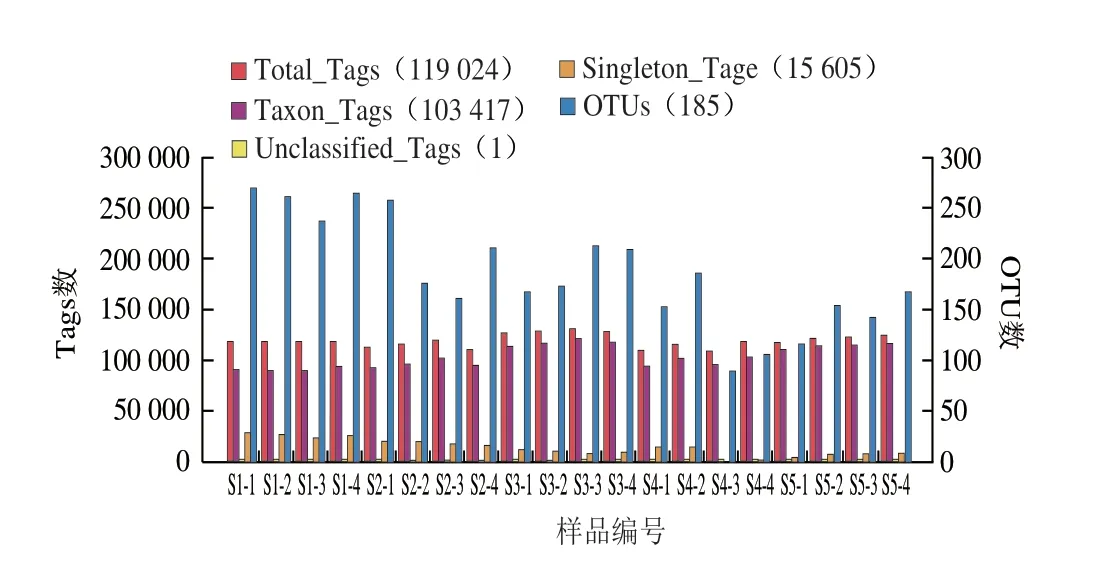
圖2 20 例樣品中Tags及OTU數Fig.2 Statistics of the number of tags and OTUs in 20 samples
2.1.1α多樣性分析
由表1可知,樣品的OTU覆蓋率均為100%,說明測序結果可以反映5 種豆豉樣品中細菌多樣性。Chao指數和ACE指數體現物種的豐富度信息,數值越大,多樣性越高;Shannon指數和Simpson指數綜合體現物種的豐富度和均勻度,數值越大,多樣性越高。S1、S2、S3物種多樣性和豐富度較高;S4、S5物種多樣性和豐富度較低,S4的Shannon指數和Simpson指數與其OTU數、Chao指數和ACE指數的趨勢不一致。
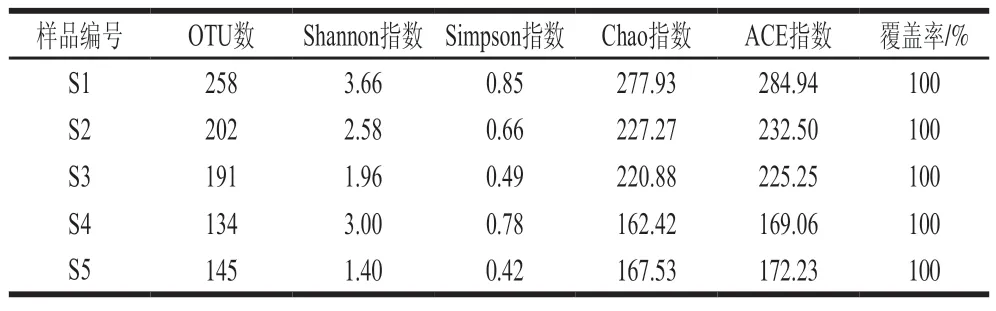
表1 樣品組內多樣性分析Table 1 Diversity analysis within sample groups
2.1.2β多樣性分析
2.1.2.1 PCA
由圖3可知,對各樣本中細菌相對豐度進行PCA時,PC1和PC2的貢獻率分別為67.47%和21.34%,累計貢獻率達88.81%,說明能很好區分5 種豆豉細菌群落。S1分布于第3象限,S2、S4分布于第2象限,S3、S5分布于第1象限,分布于同一象限兩組之間距離最近,且聚集度較高,說明其細菌群落結構較為相似。而第3象限的S1與第2象限的S2、S4在PC1上距離較近,因此,S1與S2、S4在微生物群落組成和結構上也存在一定相似。
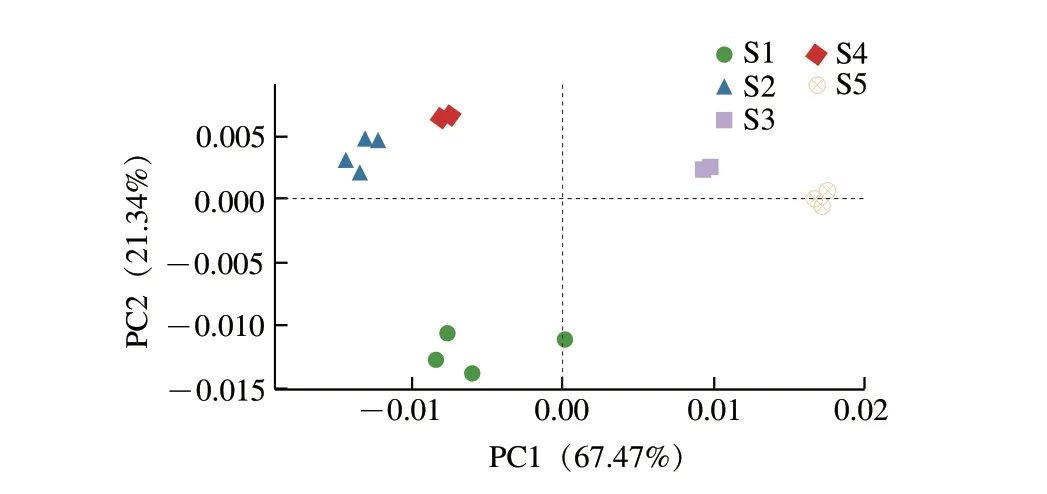
圖3 屬水平樣品加權PCAFig.3 Weighted PCA at genus level
2.1.2.2 UPGMA聚類樹分析
基于加權對樣品屬水平進行聚類分析,如圖4所示,5 種豆豉樣品可分為3 大類,S1獨立為一簇(I),S2、S4聚類為一簇(II),S3、S5聚類為一個簇(III);且第I簇與第II簇菌群結構較接近,其結果與加權PCA結果一致。
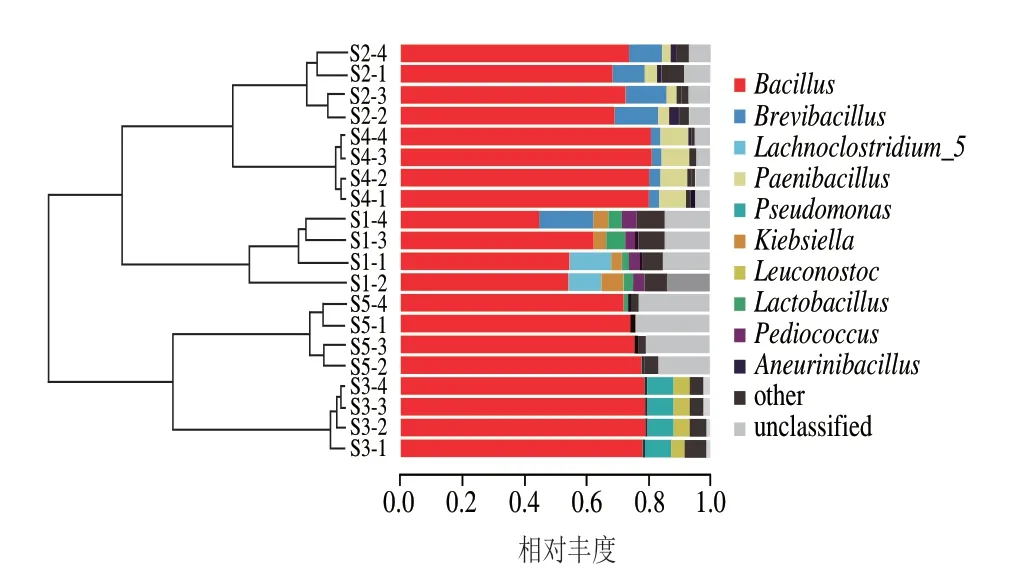
圖4 樣本間β多樣指數UPGMA聚類圖Fig.4 UPGMA clustering diagram of β-diversity index between samples
2.2 細菌群落結構分析
2.2.1 恩施豆豉門水平細菌群落結構分析
以平均相對豐度大于0.1%作為優勢菌門的評判標準,由圖5可知,恩施細菌淡豆豉中優勢菌門分別為厚壁菌門(88.18%(平均相對豐度,下同))、變形菌門(7.59%)、藍細菌門(Cyanobacteria,4.05%)、放線菌門(0.10%),且5 種樣品中優勢細菌門豐度有所差異,S1中厚壁菌門(82.62%)和變形菌門(16.49%)為主要優勢細菌門;S2和S4中均以厚壁菌門為絕對核心優勢菌門(98.31%和99.42%);S3中厚壁菌門(84.68%)和變形菌門(14.02%)為主要優勢細菌門;S5中厚壁菌門(75.88%)和藍細菌門(18.42%)為主要優勢細菌門。
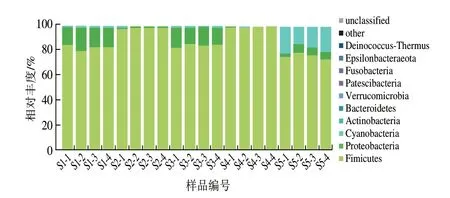
圖5 樣品中細菌門水平分布Fig.5 Distribution of bacterial phyla among samples
據報道厚壁菌門廣泛存在于發酵食品中[16];Yang Lin等[17]研究發現中國傳統豆豉的優勢菌群為厚壁菌門、放線菌門和變形菌門;貴州水豆豉中的厚壁菌門為絕對優勢菌,其次是變形菌門[18];江西豆豉中主要為藍細菌門、厚壁菌門[19];湖南豆豉中,厚壁菌門為其主要優勢菌群[20]。因此,在門水平上,各地細菌性豆豉的優勢菌群有高度一致性,厚壁菌門是最普遍、最豐富的菌門。
2.2.2 恩施豆豉屬水平細菌群落結構分析
選取所有樣本中相對豐度均值前10 位的菌屬作為優勢細菌屬,由圖6可知,恩施豆豉的優勢菌屬分別為芽孢桿菌屬(70.81%)、短芽孢桿菌屬(Brevibacillus,3.20%)、梭狀芽孢桿菌5(Lachnoclostridium_5,3.14%)、類芽孢桿菌屬(Paenibacillus,2.50%)、假單胞菌屬(Pseudomonas,1.87%)、克雷伯菌屬(Klebsiella,0.97%)、明串珠菌屬(Leuconostoc,0.97%)、乳桿菌屬(Lactobacillus,0.84%)、片球菌屬(Pediococcus,0.75%)、神經氨酸桿菌屬(Aneurinibacillus,0.68%)。其中芽孢桿菌屬為核心優勢細菌屬。不同樣品中優勢細菌屬的豐度不同,芽孢桿菌屬在S1、S2、S3、S4、S5中的相對豐度分別為48.77%、70.85%、78.89%、80.53%、75.04%。5 種樣品中次要菌群各不相同,S1中為梭狀芽孢桿菌5(15.24%),S2中為短芽孢桿菌屬(12.18%),S3中假單胞菌屬較豐富,相對豐度為8.82%;S4含8.80%的短芽孢桿菌屬(Brevibacillus);S5含0.36%假單胞菌屬。各樣品中不可鑒定到屬水平的序列含量相似,平均相對豐度為10.73%,說明恩施細菌型淡豆豉中仍有未發掘的細菌屬。
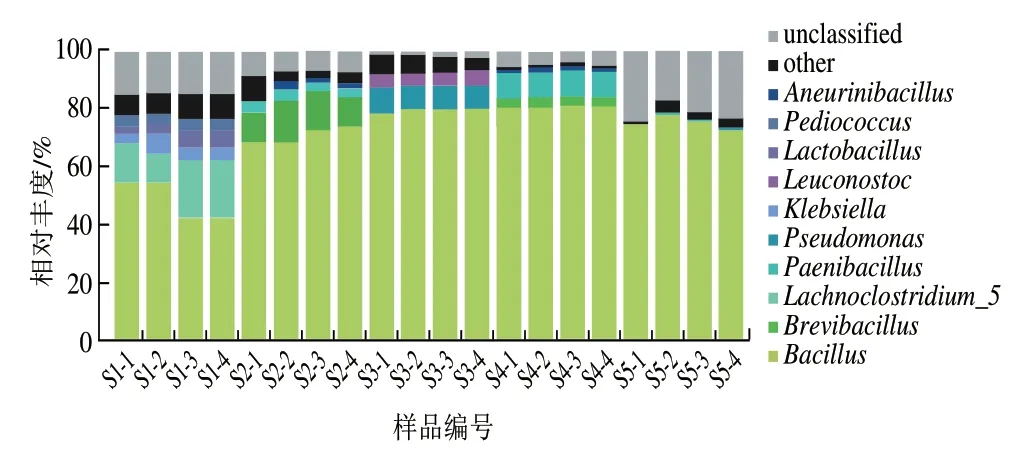
圖6 樣品細菌屬水平分布Fig.6 Distribution of bacterial genera among samples
貴州水豆豉中芽孢桿菌屬是絕對優勢菌屬,其次是變形桿菌屬[18];江西豆豉主要為藍細菌門未分類屬(unclassified_Cyanobacteria)、葡萄球菌屬(Staphylococcus)和乳桿菌屬[19];湖南豆豉發酵過程中,芽孢桿菌屬相對豐度高,乳酸菌種類較多、相對豐度較高[20]。由此可見,各地細菌型豆豉的發酵主要為厚壁菌門中芽孢桿菌屬,可能由于地區間環境微生物和發酵制作工藝差異等,微生物群落結構中個別菌群存在差異,從而導致各地豆豉間風味和營養不同。在豆豉發酵過程中,芽孢桿菌屬、乳桿菌屬具有益生菌潛力[21]。神經氨酸桿菌和短桿菌屬有助于豆豉香氣形成[22],短桿菌發酵可以產生谷氨酸[23],而大多數假單胞菌可以產生一種或多種蛋白酶、脂肪酶和卵磷脂酶[24],從而增強豆豉的風味。
2.2.3 恩施豆豉種水平細菌群落結構分析
由圖7可知,豆豉樣品中不可鑒定到種水平的序列含量較高。樣品S2、S3、S4中不可鑒定到種水平的序列水平相似,平均相對豐度為14.87%。S1中鑒定到種水平的序列相對豐度為36.64%,S5僅鑒定出2.64%的種水平序列。說明在S1、S5中有大量種水平細菌未被鑒定出。
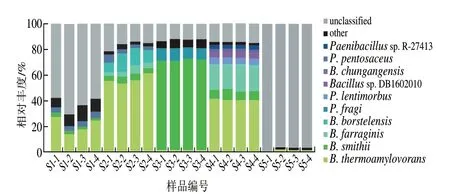
圖7 樣品細菌種水平分布Fig.7 Distribution of bacterial species among samples
5 種恩施豆豉種水平的優勢細菌均為芽孢桿菌屬,相對豐度前10 位的優勢菌種分別為嗜熱淀粉芽孢桿菌(Bacillusthermoamylovorans,23.77%)、史密斯芽孢桿菌(Bacillus smithii,17.16%)、蘇云金芽孢桿菌(Bacillus farraginis,4.73%)、波茨坦短芽孢桿菌(Brevibacillus borstelensis,2.58%)、脆假單胞菌(Pseudomonas fragi,1.78%)、遲鈍芽孢桿菌(Paenibacillus lentimorbus,1.68%)、類芽孢桿菌亞種DB1602010(Bacillussp.DB1602010,0.81%)、中港芽孢桿菌(Bacilluschungangensis,0.80%)、發酵乳桿菌(Pediococcus pentosaceus,0.74%)、類芽孢桿菌亞種R-27413(Paenibacillussp.R-27413,0.72%)。其中嗜熱淀粉芽孢桿菌、史密斯芽孢桿菌為核心優勢菌種,在5 種樣品中均存在,但相對豐度有差異。S1中嗜熱淀粉芽孢桿菌相對豐度為20.48%,S2中為56.22%,S3中為1.32%,S4中為40.02%,S5中為0.82%。5 種恩施豆豉中史密斯芽孢桿菌相對豐度為0.24%~70.36%,在S3中最豐富。此外,5 種樣品中次要菌種差異較大,S1為戊糖片球菌(Pediococcus pentosaceus)(3.59%),S2、S4為蘇云金芽孢桿菌(4.06%和19.23%),S3為脆假單胞菌(8.80%),S5為中港芽孢桿菌(0.13%)。
Chen Tingtao等[25]采用聚合酶鏈反應-變性梯度凝膠電泳發現枯草芽孢桿菌、解淀粉芽孢桿菌、假單胞菌、釀酒酵母和粉狀畢赤酵母在大多數豆豉樣品的菌群結構中具有顯著地位。黃曉潤等[26]研究發現,貴州自然發酵型水豆豉制曲和后發酵期間的優勢菌種為枯草芽孢桿菌、嗜熱淀粉芽孢桿菌和地衣芽孢桿菌(Bacillus licheniformis)。研究表明,豆豉發酵過程中,芽孢桿菌代謝產生的各種酶類使脂肪、蛋白質、碳水化合物和黃酮苷類分解轉化為有機酸、氨基酸和糖苷配基等,是豆豉特有風味、營養形成的關鍵菌[27]。
3 結論與討論
在5 種恩施州細菌型淡豆豉菌群結構中共鑒定出17 門、24 綱、43 目、78 科、152 屬以及93 種。其中厚壁菌門占絕對優勢,其次是變形菌門、藍細菌門、放線菌門。優勢菌種為厚壁菌門的芽孢桿菌屬中的嗜熱淀粉芽孢桿菌和史密斯芽孢桿菌。由于恩施豆豉為自然發酵生產,因此其微生物種屬非常豐富,本研究中還檢測到短芽孢桿菌屬、梭狀芽孢桿菌5、類芽孢桿菌屬、假單胞菌屬、克雷伯菌屬、明串珠菌屬、乳桿菌屬、片球菌屬、神經氨酸桿菌屬。后期需深入研究恩施豆豉菌群中是否存在高產功能成分的菌種,以提高產品的健康性。
芽孢桿菌屬作為核心優勢細菌群,在細菌型淡豆豉的發酵過程中發揮至關重要的作用。日本納豆中主要為芽孢桿菌屬枯草芽孢桿菌亞種——納豆芽孢桿菌(Bacillus natto)[28],其高產具有溶栓作用的納豆激酶。國內多地利用高通量測序技術對豆豉的優勢菌群進行研究,經對比分析,恩施州細菌型豆豉的優勢菌群與甘肅[29]和貴州[18]相似,以芽孢桿菌屬為主。芽孢桿菌屬作為核心優勢菌群,可產生大量的酶,如蛋白酶[30]和淀粉酶[31],因此,芽孢桿菌的存在可以加速豆豉的發酵過程,并提供其他細菌所需營養物質[32]。此外,各地豆豉呈現顯著的細菌多樣性,相應也表現出風味多樣性。
本研究中S1、S2、S3、S5取樣地點接近,樣品的微生物菌群組成并未表現出地域相似性,β多樣性分析顯示S2與S4、S3與S5較相似,S1獨立成一簇。因此,菌群的差異性主要來自于生產時的原始菌群和生產環境的微生物控制。
Chen Tingtao等[33-34]在豆豉制曲過程中檢測到葡萄球菌、腸桿菌和氣球菌等病原菌。貴州自然發酵水豆豉中檢測到黏質沙雷菌(Serratia marcescens)、糞產堿菌(Alcaligenes faecalis)、蠟樣芽孢桿菌和奇異變形桿菌(Proteus mirabilis)4 種致病菌[26];在云南豆豉中發現有屬于人類致病菌和植物病原菌的序列[35]。本研究在S1中檢測到梭狀芽孢桿菌5(15.24%)、克雷伯菌屬(4.68%)這類能引起炎癥的病原菌,說明其在加工過程中可能被條件致病菌污染,其他樣品中也存在微量梭狀芽孢桿菌屬、克雷伯菌屬和沙雷氏菌。因此,在傳統豆豉制作過程中應注意控制發酵條件以減少致病菌污染。
